Identification and functional analysis of two oligopeptide transporters supporting the growth of Lacticaseibacillus paracasei strain Shirota in milk
Shiho Tanaka, Masaki Serata, Tomohiko Terai, Daichi Fujii, Takekazu Okumura

TL;DR
This study identifies two oligopeptide transporters in Lacticaseibacillus paracasei Shirota that are essential for its growth in milk by transporting nitrogen sources.
Contribution
The first experimental evidence of oligopeptide transporters in Lactobacilli capable of transporting peptides up to eight residues long.
Findings
OppLc and DppLc are the primary oligopeptide transporters in LcS, essential for growth in milk.
OppLc transports peptides of 4–8 residues, while DppLc transports peptides of 3–7 residues.
OppA1 is the only oligopeptide-binding protein in OppLc, while DppA2 and DppA3 may function as subunits in DppLc.
Abstract
Lacticaseibacillus paracasei strain Shirota (LcS) employs two distinct oligopeptide transporters, OppLc and DppLc, to facilitate nitrogen acquisition during growth in milk. These transporters exhibit differential substrate specificity, enabling the uptake of oligopeptides of varying lengths derived from milk proteins. Oligopeptide transporters are important proteins in several lactic acid bacteria (LAB) that facilitate the transport of oligopeptides, the primary nitrogen source for growth in milk. Although the proteolytic mechanisms are well understood in some LAB species, limited research has been conducted on the peptide transport systems of Lacticaseibacillus paracasei (formerly Lactobacillus casei) strain Shirota (LcS), particularly its oligopeptide transporters. This study investigated the nitrogen uptake mechanism of LcS, a probiotic lactic acid bacterium, by generating gene…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
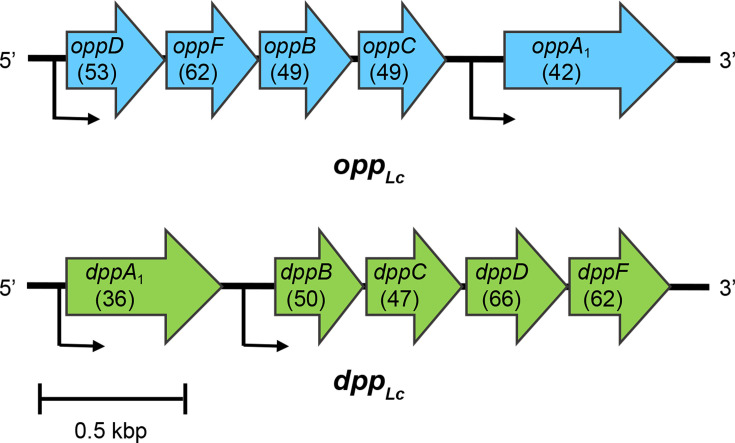 Fig. 1
Fig. 1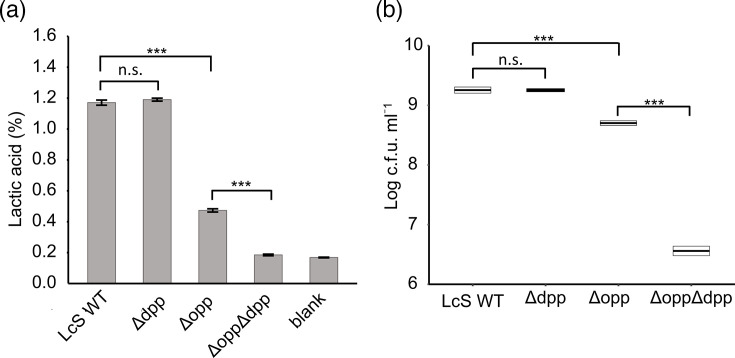 Fig. 2
Fig. 2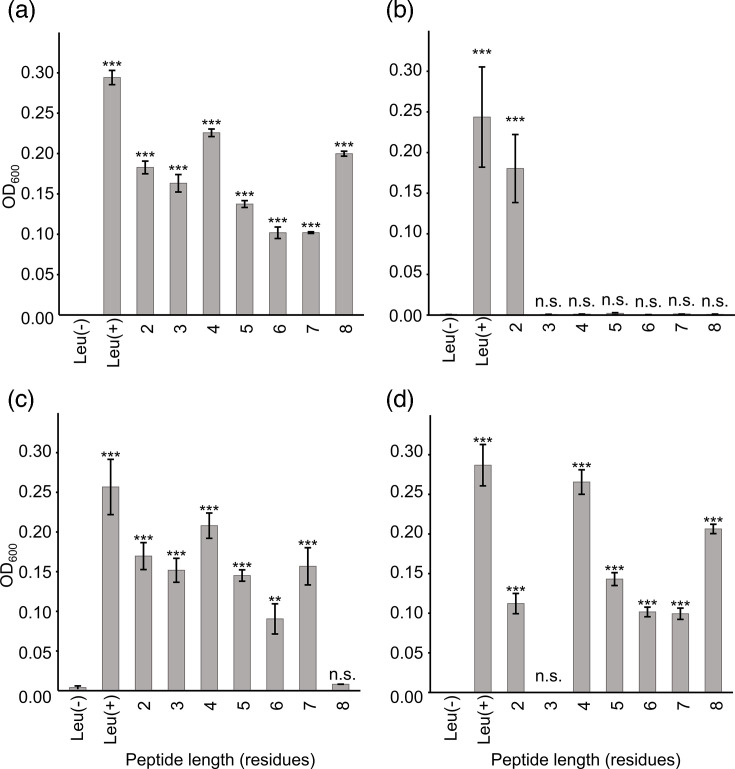 Fig. 3
Fig. 3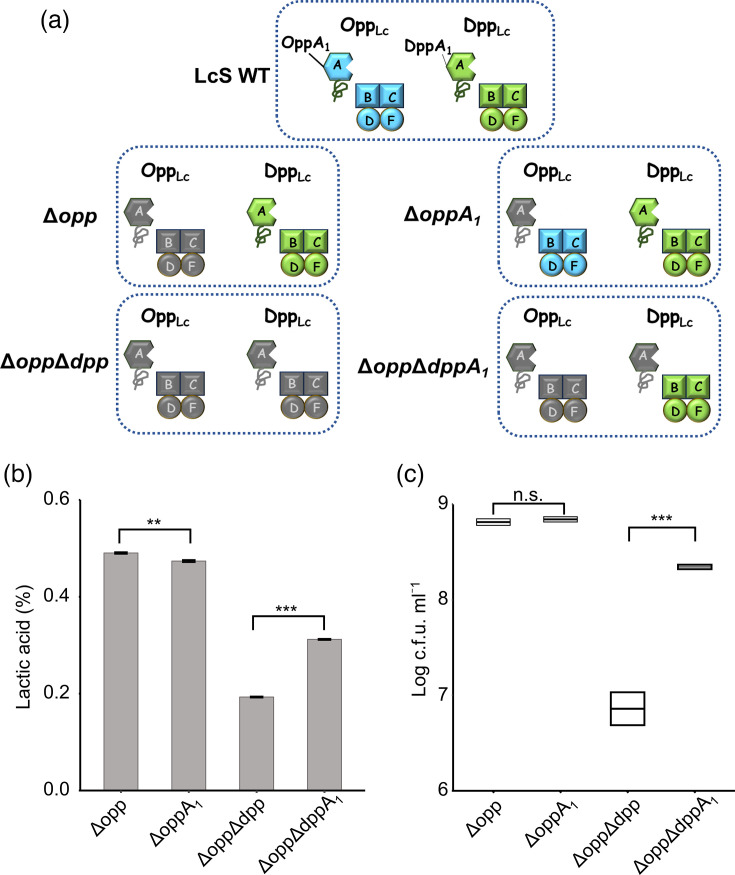 Fig. 4
Fig. 4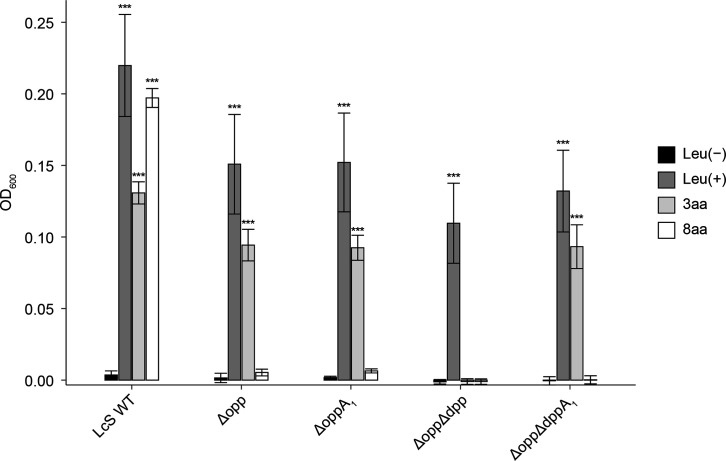 Fig. 5
Fig. 5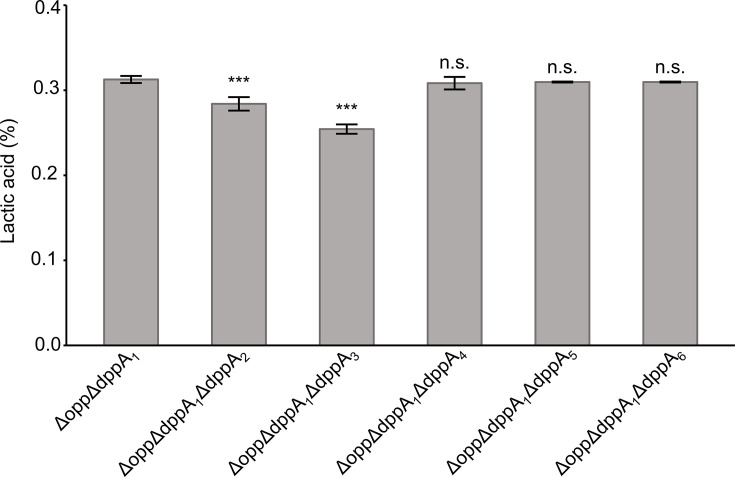 Fig. 6
Fig. 6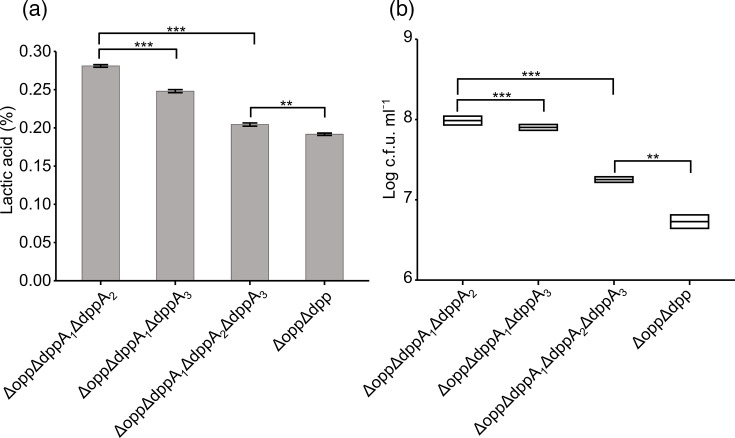 Fig. 7
Fig. 7| Strain or plasmid | Characteristic or relevant genotype | Source or reference |
|---|---|---|
|
| ||
| JM109 | recA1, endA1, gyrA96, thi-1, hsdR17(rK− mK+), e14− (mcrA−), supE44, relA1, Δ (lac-proAB)/F' (traD36, proAB+, lacIq, lacZΔM15) | TaKaRa Bio |
|
| ||
| YIT 9029 (strain Shirota: LcS) | WT | Our collection |
| Δ | OppLc− derivative of LcS | This study |
| Δ | DppLc− derivative of LcS | This study |
| Δ | OppLc− DppLc− derivative of LcS | This study |
| Δ | OppA1− derivative of LcS | This study |
| Δ | OppLc− DppA1− derivative of LcS | This study |
| Δ | OppLc− DppA1− DppA2− derivative of LcS | This study |
| Δ | OppLc− DppA1− DppA3− derivative of LcS | This study |
| Δ | OppLc− DppA1− DppA4− derivative of LcS | This study |
| Δ | OppLc− DppA1− DppA5− derivative of LcS | This study |
| Δ | OppLc− DppA1− DppA6− derivative of LcS | This study |
| Δ | OppLc− DppA1− DppA2− DppA3− derivative of LcS | This study |
| Δ | Δ | This study |
| Plasmids | ||
| pYSSE3 | [ | |
| pYAP300 | [ | |
| pYSSE3-Δopp | pYSSE3 carrying upstream region with the N-terminus of | This study |
| pYSSE3-Δdpp | pYSSE3 carrying upstream region with N-terminus of | This study |
| pYAP300-opp | pYAP300 carrying | This study |
| pYSSE3-ΔoppA1 | pYSSE3 carrying upstream region with N-terminus of | This study |
| pYSSE3-ΔdppA1 | pYSSE3 carrying upstream region with N-terminus of | This study |
| pYSSE3-ΔdppA2 | pYSSE3 carrying upstream region with N-terminus of | This study |
| pYSSE3-ΔdppA3 | pYSSE3 carrying upstream region with N-terminus of | This study |
| pYSSE3-ΔdppA4 | pYSSE3 carrying upstream region with N-terminus of | This study |
| pYSSE3-ΔdppA5 | pYSSE3 carrying upstream region with N-terminus of | This study |
| pYSSE3-ΔdppA6 | pYSSE3 carrying upstream region with N-terminus of | This study |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Bacterial Genetics and Biotechnology · Amino Acid Enzymes and Metabolism
Data Summary
The authors confirm that all supporting data and protocols have been provided within the article or through supplementary data files. The series of genes comprising opp_Lc_ and dpp_Lc_ was registered as a gene cluster in the DDBJ under accession numbers LC855157 and LC855158, respectively. The nucleotide sequences of dppA_2_ (accession no. LC855159), dppA_3_ (accession no. LC855160), dppA_4_ (accession no. LC855161), dppA_5_ (accession no. LC855162) and dppA_6_ (accession no. LC855163) were also registered.
Introduction
Lactic acid bacteria (LAB) obtain various aa from their environment that cannot be biosynthesized [12]. In milk, proteins are the primary nitrogen source for LAB owing to their limited free aa. To meet their nutritional requirements, LAB have developed proteolytic systems that hydrolyse milk proteins and absorb the resulting aa and peptides [3]. These processes have been well studied in dairy-associated strains, such as Lactococcus lactis [45], Lactobacillus helveticus [67], and Streptococcus thermophilus [8]. Despite species and strain differences, LAB typically process milk proteins through three main steps: (1) degradation of milk proteins into peptides by extracellular proteases, (2) transport of peptides – primarily oligopeptides – into bacterial cells and (3) breakdown of peptides into aa by intracellular peptidases [3].
The uptake of oligopeptides by LAB depends on oligopeptide transporters. In L. lactis, the most studied ABC oligopeptide transporter is the oligopeptide transport system (Opp system) [910], which includes the components OppA, OppB, OppC, OppD and OppF, each with specific functions in peptide transport. OppA is a membrane-associated substrate-binding protein, OppB and OppC form transmembrane domains and OppD and OppF are intracellular ATP-binding proteins. Although opp genes are conserved in lactobacilli [11], peptide transport characteristics vary among species and strains [12]. OppA initiates peptide transport via the Opp transport system and is a key for determining substrate specificity [13]. Studies on Opp-transported peptides have shown that OppA binds to various peptides, with 4–35 residue peptides as the primary targets of OppA in L. lactis [14]. Di- and tripeptides that cannot be transported by the Opp system are transported by the di-tripeptide transport system (Dpp system) [15], which includes five DppABCDF subunits and belongs to the same ABC transporter superfamily as the Opp system. Additionally, the L. lactis di-tripeptide transport protein (DtpT) system [16], which belongs to the peptide transport family, contains a single subunit and uses a proton concentration gradient for transport.
Variations in the number of oligopeptide-binding proteins (OBPs) in Opp and Dpp systems differ among species and strains [12]. L. lactis possesses only one OppA; however, some LAB have multiple copies of OBPs in their Opp systems. For example, the Ami transport system, which is analogous to the Opp system in S. thermophilus, uses three peptide-binding proteins as a single oligopeptide transporter [17]. In the L. lactis Dpp system, a second OBP, DppP, functions alongside DppA [1518]. Among lactobacilli, only Lactobacillus bulgaricus has been shown to express two OppAs as operons [19]. Hence, the presence of multiple OBP is considered a microbial survival strategy to enhance transport efficiency in the peptide transport system, as it increases both the number of OBP molecules and the types of peptides that can be transported.
The molecular mechanism of peptide transport in lactococci, especially L. lactis, which is widely used in dairy production, has been thoroughly studied since the early 1980s, aided by advancements in genetic engineering [20]. However, much less is known about these mechanisms in L. bulgaricus and L. helveticus, despite their importance in yoghurt and cheese production. Furthermore, few studies have clarified the characteristics of their oligopeptide transporters. This scarcity could be due to the underexploration of efficient genetic engineering tools, especially for L. bulgaricus, which have only recently been established [21]. Nevertheless, genetic engineering approaches, including knockout (KO) strain generation, have been established for the Lacticaseibacillus paracasei strain Shirota (LcS), the strain used in this study [22]. Although L. paracasei is often isolated from non-dairy environments, such as the intestinal tract and oral cavity [2324], LcS has a long history of use in dairy products, suggesting it may be uniquely adapted for growth in milk.
Given the limited functional analyses of peptide transport systems in lactobacilli, in this study, we aimed to investigate the nitrogen utilization mechanisms in LcS. Specifically, we sought to identify the key transport systems that facilitate growth in milk. Understanding these mechanisms will not only provide insights into the metabolic adaptations of LcS for dairy environments but also contribute to the broader understanding of LAB peptide transport systems. The findings may provide useful insights for optimizing dairy fermentation processes.
Methods
Bacterial strains
The strains and plasmids used in this study are listed in Table 1. Escherichia coli and L. paracasei strains were cultivated in Luria-Bertani medium or Difco Lactobacilli de Man, Rogosa, Sharpe (MRS) broth (BD Biosciences), respectively, and subsequently preserved at −80 °C. Frozen stock was used as the inoculum for subsequent experiments.
Sequence analysis and construction of gene engineering strains
Gene and aa sequence analyses, as well as promoter identification, were conducted using Genetyx version 14.1.0 (NIHON SERVER Co., Ltd.). Gene KO and genetic complementation strains were generated using a method described previously with slight modifications [2526]. Briefly, KO strains were generated using PCR by synthesizing two gene fragments, each containing the upstream and downstream regions of the target gene. These fragments were subsequently ligated into the pYSSE3 vector using restriction enzymes or an In-Fusion HD Cloning Kit (TaKaRa Bio Inc.). Plasmids carrying the gene fragments were first introduced into the E. coli strain JM109 via electroporation. After amplification within the bacterial cells, the plasmids were extracted and subsequently introduced into the LcS WT or KO strains via electroporation. Single-crossover strains harbouring the gene of interest were then selected. Erythromycin-sensitive strains that emerged by homologous recombination in the process of repeated passaging of the selected single cross-over strains on medium without erythromycin were selected as KO strains (Fig. S1, available in the online Supplementary Material). To generate the opp_Lc_ complemented strain (Δopp^c^), the gene region containing the predicted Shine–Dalgarno and CDSs of opp_Lc_ was cloned into the pYAP300 vector [22], designated as pYAP300-opp. This plasmid (pYAP300-opp) was then introduced into the opp_Lc_ KO strain (Δopp) via transformation. Erythromycin-resistant strains were selected using PCR for the presence of the opp_Lc_ insert. The resulting strain was designated as Δopp^c^. All primers used to generate the transgenic strains are listed in Table S1.
Growth assays
To prepare the milk medium, skimmed milk powder (Megmilk Snow Brand) was reconstituted in water at a final concentration of 10% (w/v) and sterilized by autoclaving at 121 °C for 15 min. To evaluate the growth of each strain in the MRS medium, each frozen stock was inoculated into the MRS medium at 0.1% (v/v) and incubated for 24 h at 37 °C. The bacterial culture was transferred to fresh MRS medium under the same conditions and incubated for 24 h. For growth evaluation in milk, the inoculum was added at 0.1% (v/v) to a milk medium and incubated for 48 h. Each solution was diluted with 0.85% (w/v) saline to the appropriate concentration and spread on MRS agar plates using a spiral plate (EDDY JET2; IUL Instruments). The plates were incubated at 37 °C for 72 h, and the colony counts as c.f.u. were quantified using ProtoCOL3 (Synbiosis). Cumulative organic acid production by LAB during growth, an alternative indicator of bacterial proliferation, was assessed by measuring titration acidity. Each sample was diluted to the appropriate concentration in Milli-Q water, and the volume of 0.1 N NaOH needed to reach pH 8.5 was calculated per 9 g sample using an automatic acidity titrator (HIRANUMA Co., Ltd., Mito City, Japan). The percentage of lactic acid in a 9 g sample was calculated using the following formula:
Transport test of leucine-containing peptides
Chemically defined medium lacking leucine [CDM (Leu−)] was prepared based on the composition reported by Alcántara et al. for culturing L. paracasei BL23 strain [27]. l-Lysine and l-threonine were added at 0.25 g l^−1^ to this medium, whereas leucine was excluded from the composition. Leucine or leucine-containing peptides were added to CDM at a final concentration of 100 µM, and 100 µl of the mixture was dispensed into each well of a 96-well plate. Leucine-containing peptides were selected based on sequences described by Tynkkynen et al. [9] and included Leu-Gly, Leu-Gly-Gly and Tyr-Gly-Gly-Phe-Leu (Leu-enkephalin), which were purchased from Peptide Institute, Inc. (Osaka, Japan). Additional peptides, including Gly-Leu-Gly-Lys, Tyr-Gly-Gly-Phe-Leu-Lys (Leu-enkephalin-Lys), Ser-Ile-Gly-Ser-Leu-Ala-Lys and Val-His-Leu-Thr-Pro-Val-Gly-Lys, were synthesized by Eurofins Genomics Inc. (Tokyo, Japan) with a purity of >95 %. Bacteria cultured for 24 h in MRS medium were diluted tenfold in 0.85% (w/v) saline, and 1% (v/v) of the diluted mixture was added to CDM (Leu−) containing each peptide. The cultures were incubated at 37 °C for 48 h. Turbidity at OD_600_ was measured using a SpectraMax M2 plate reader (Molecular Devices LLC, CA, USA).
Statistical analysis
All statistical analyses were performed using R version 4.4.1 [28]. For selected pairwise comparisons of interest – that is, specific pairs of data points chosen for statistical comparison based on the experimental design and research questions – Welch’s t-tests were performed, and P-values were adjusted using the Bonferroni correction to account for multiple testing. To compare each group with a designated control, one-way ANOVA was conducted using the aov function, followed by Dunnett’s post hoc test using the glht function in the ‘multcomp’ package [29], with the mcp method set to ‘Dunnett’. All visualizations were generated using the ‘ggplot2’ package [30] or Microsoft PowerPoint.
Results
Search for oligopeptide transporter candidate genes encoded in the LcS genome
To identify the oligopeptide transporters important for LcS growth in milk, we investigated the presence of Opp or Dpp systems in LcS by performing a homology search using L. lactis sequences as references [910]. Two operons, opp and dpp, encoding oligopeptide transporters, were identified in LcS (Fig. 1). The opp operon gene of LcS (oppLc) was located on a complementary strand in the order oppDFBCA1, with each subunit showing high identity to the corresponding aa sequences in the L. lactis Opp transport system: 53%, 62%, 49%, 49% and 42%, respectively (Fig. 1). Similarly, the dpp operon gene (dppLc) was also encoded in the order dppA1BCDF, with each subunit showing high homology to the L. lactis Dpp system at 36%, 50%, 47%, 66% and 62%, respectively, demonstrating high similarity in all subunits, as observed in oppLc (Fig. 1). In both LcS transport systems, oppD and oppF in oppLc and dppF and dppD in dppLc contain highly conserved ATP-binding domains across different species and show higher homology than other subunits. Additionally, promoters were predicted at two locations in each operon: one upstream of the substrate-binding protein genes (oppA1 and dppA1) and another upstream of the remaining four genes in each operon (Fig. 1).
Genetic organization of the oppLc and dppLc operons in LcS. The predicted promoters are indicated using arrows. Numbers in parentheses indicate the percentage of identical aa residues in the amino sequence via sequence comparison with opp (accession no. L18760) or dpp (accession no. AF247635) of L. lactis.
Growth of KO strains lacking oppLc and dppLc in milk
Investigating the functional significance of Opp_Lc_ and Dpp_Lc_ in LcS grown in milk is crucial. To this end, we generated three KO strains, Δopp, Δdpp and ΔoppΔdpp, in which opp_Lc_, dpp_Lc_ or both operons were genetically disrupted and compared their growth performance in milk with that of WT strains. The viable cell counts and titration acidity of the three test strains and the WT strain cultured in MRS medium were as follows: all strains exhibited viable cell counts ranging from 1.6×10⁹ to 2.1×10⁹ c.f.u. ml^−1^ and titration acidity values between 1.44 and 1.49%, which were comparable to those of the WT strain (2.0×10⁹ c.f.u. ml^−1^ and 1.46%, respectively) (Fig. S2). These findings indicate that all strains grew sufficiently under the preculture conditions. These parameters were subsequently measured in the samples after inoculating the cultures into milk medium. The viable cell count of the Δdpp strain (1.8×10^9^ c.f.u. ml^−1^) was comparable to that of the LcS WT strain (2.9×10^9^ c.f.u. ml^−1^). In contrast, Δopp and ΔoppΔdpp strains showed ~5.8-fold and 793-fold lower counts than LcS WT, at 5.0×10^8^ and 3.7×10^6^ c.f.u. ml^−1^, respectively (Fig. 2a). The titration acidity also showed a similar trend, with 1.06 and 1.19% in LcS WT and Δdpp strains, respectively, whereas it was reduced in Δopp and ΔoppΔdpp strains (0.41 and 0.19 %, respectively) (Fig. 2b). The average acidity of the milk medium without the inoculum was 0.17% (Fig. 2a). The initial bacterial count was ~1×10^6^ c.f.u. ml^−1^. The ΔoppΔdpp strain showed almost no growth and produced no acid during the 48-h incubation period. To confirm that the reduced proliferation of Δopp in milk medium was due to the loss of opp_Lc_ function, a genetic complementation strain, Δopp^c^, was generated and grown under the same conditions. Δopp^c^ demonstrated a significant recovery in viability, with its viable cell count reaching nearly the same level as that of LcS WT (Fig. S3A). The titration acidity of Δopp^c^ was marginally lower than that of LcS WT but was notably higher than that of Δopp (0.58%) (Fig. S3B).
*Growth characteristics of oppLc and dppLc KO strains. (a, b) Viable cell counts (a) and titratable acidity (b) of LcS WT, KO strains and a complemented strain after 48 h of growth in milk medium. Error bars indicate sd (n=3) for each graph. Only comparisons of interest are indicated with brackets. **P<0.001, n.s. assessed using Welch’s t-test with Bonferroni correction. ∆opp, KO strain lacking oppLc; Δdpp, KO strain lacking dppLc; ΔoppΔdpp, KO strain lacking oppLc and dppLc.
Characteristics of peptide lengths transported by KO strains oppLc and dppLc
As LcS is leucine auxotrophic, it cannot grow under leucine-deficient conditions [2]. Therefore, we grew the WT as well as the mutants in media supplemented with peptides of different lengths to assess whether functional differences between the two proposed oligopeptide transporters, Opp_Lc_ and Dpp_Lc_, contribute to growth in milk. We investigated the peptide lengths transported by these oligopeptide transporters. LcS WT grew under all conditions when peptides consisting of 2–8 residues were added (Fig. 3a). In contrast, the deletion of both opp_Lc_ and dpp_Lc_, referred to as ΔoppΔdpp, resulted in no growth when peptides containing 3–8 residues were added (Fig. 3b), suggesting additional pathways are involved in transporting 2-residue peptides. A similar analysis with Δopp showed growth in the presence of 2–7 residue peptides but not with 8-residue peptides (Fig. 3c). For Dpp_Lc_, Δdpp grew with the addition of 2 or 4–8 residue peptides but not with 3-residue peptides (Fig. 3d). These findings suggest that Opp_Lc_ and Dpp_Lc_ are involved in the uptake of 4–8- and 3–7-residue peptides, respectively, under the tested conditions. Specifically, Dpp_Lc_ and Opp_Lc_ were responsible for the uptake of the 3- and 8-residue peptides, respectively, whereas the 4–7-residue peptides were taken up by both oligopeptide transporters.
*Peptide transport ability of oppLc and dppLc KO strains. WT LcS (LcS WT) and gene KO strains were grown for 48 h in CDM lacking leucine and supplemented with a leucine-containing peptide. (a) LcS WT; (b) ΔoppΔdpp, KO strain lacking oppLc and dppLc; (c) Δopp, KO strain lacking oppLc; (d) Δdpp, KO strain lacking dppLc. Error bars indicate sd (n=2) for each graph. Data represent the means of two parallel experiments. Asterisks indicate significant differences from the Leu(-) group assessed using Dunnett’s test. ***P<0.001, *P<0.01 and n.s., P≥0.1. ΔoppΔdpp, KO strain lacking oppLc and dppLc; Δopp, KO strain lacking oppLc; Δdpp, KO strain lacking dppLc.
Growth of KO strains lacking oppA1 and dppA1
To identify the OBPs that constitute Opp_Lc_ and Dpp_Lc_ and determine if OBPs other than OppA_1_ and DppA_1_ function as Opp_Lc_ and Dpp_Lc_ subunits, we generated oppA_1_ and dppA_1_ gene disruption strains ΔoppA_1_ and ΔoppΔdppA_1_ and examined their peptide transport capacity (Fig. 4a). In the absence of functional OBPs other than OppA_1_ or DppA_1_, ΔoppA_1_ and ΔoppΔdppA_1_ were expected to completely lose their peptide transport ability via OppA_1_ or DppA_1_, resulting in growth potential equivalent to that of oligopeptide transporter deletion strains, Δopp and ΔoppΔdpp, respectively. Although a significant difference in titration acidity was observed between the two strains (estimated difference: 0.0167, 95% confidence interval: 0.012–0.021), no significant difference was found in viable cell counts, suggesting that the acidity difference is not biologically meaningful. In contrast, the titration acidity and viable cell count of ΔoppΔdppA_1_ were significantly higher than those of ΔoppΔdpp by ~0.2% and 29 times, respectively (Fig. 4b, c).
*Growth characteristics of KO strains of OBP. (a) Schematic representation of OppLc and DppLc expression in each strain. Defective proteins in each gene KO strain are indicated in grey. (b, c) Titration acidity (b) and viable cell counts (c) of KO strains after 48 h of growth in milk. Error bars indicate the sd (n=3) for each graph. Only comparisons of interest are indicated with brackets. ***P<0.001, *P<0.01, n.s., P≥0.1 assessed using Welch’s t-test with Bonferroni correction. ∆opp, KO strain lacking oppLc; ΔoppA1, KO strain lacking oppA1; ΔoppΔdpp, KO strain lacking oppLc and dppLc; ΔoppΔdppA1, KO strain lacking oppLc and dppA1.
Characteristics of peptide lengths transported by KO strains lacking oppA1 or dppA1
To determine whether OBPs other than OppA_1_ and DppA_1_ play a role in Opp_Lc_ and Dpp_Lc_ function, we compared the characteristics of the transported peptides in the OBP-deletion strains, ΔoppA_1_ and ΔoppΔdppA_1_, with the oligopeptide transporter KO strains, Opp_Lc_ and Dpp_Lc_, respectively. In this analysis, we focused on the ability to incorporate 3- or 8-residue peptides containing leucine, which are considered responsible for the transport functions of Opp_Lc_ and Dpp_Lc_, respectively. Uptake of these peptides into each strain using [CDM (Leu−)] revealed that ΔoppA_1_, similar to Δopp, could not grow when eight residues were present (Fig. 5). In other words, both opp_Lc_-deficient (Δopp) and oppA_1_-deficient (ΔoppA_1_) strains lost their ability to take up 8-residue peptides. Conversely, the ΔoppΔdppA_1_ strain grew when three residues were present, whereas the ΔoppΔdpp strain was unable to do so (Fig. 5). These findings suggest that the ΔoppΔdppA_1_ strain differs from the ΔoppΔdpp strain in its ability to incorporate these 3-residue peptides.
*Peptide transport ability of oppA1 and dppA1 KO strains. WT LcS (LcS WT) and KO strains were cultured for 48 h in CDM (Leu−) and supplemented with leucine-containing peptide. Error bars indicate sd (n=3). Asterisks indicate significant differences from the Leu(-) group assessed using Dunnett’s test; **P<0.001. ∆opp, gene KO strain lacking oppLc genes; ΔoppA1, KO strain lacking oppA1 gene; ΔoppΔdpp, KO strain lacking oppLc and dppLc genes; ΔoppΔdppA1, KO strain lacking oppLc and dppA1 genes.
Growth of KO strains in milk compared to OBP candidate gene disruption strains (other than DppA1)
As Dpp_Lc_ may have a functional subunit other than DppA_1_, we searched the LcS genome for genes predicted to encode OBPs and examined their homology with DppA_1_. All genes encoding potential OBP subunits for Dpp_Lc_, referred to as DppA_2_ to DppA_6_, were independently encoded and did not share an operon structure with other oligopeptide transporter genes (Fig. S4). We generated KO strains for five candidate DppA proteins and investigated the titration acidity of each strain after growth in milk. The function of Dpp_Lc_ can only be displayed when Opp_Lc_ is nonfunctional. These five dppA gene disruption strains were generated using the Δopp strain as the host organism. The results showed that the titration acidity of ΔoppΔdppA_1_ΔdppA_4_, ΔoppΔdppA_1_ΔdppA_5_ and ΔoppΔdppA_1_ΔdppA_6_, targeting DppA_4_, DppD_5_ and DppA_6_, respectively, did not differ significantly compared to that of ΔoppΔdppA_1_. However, the titration acidity of the two gene disruption strains ΔoppΔdppA_1_ΔdppA_2_ and ΔoppΔdppA_1_ΔdppA_3_, which target DppA_2_ and DppA_3_, respectively, was significantly lower than that of the ΔoppΔdppA_1_ strain targeting DppA_1_. The difference in titration acidity between these two strains with ΔoppΔdppA_1_ was 0.03% and 0.06%, respectively (Fig. 6). Furthermore, we conducted the same analysis with the ΔoppΔdppA_1_ΔdppA_2_ΔdppA_3_ strain, where dppA_2_, dppA_3_ and dppA_1_ were deleted simultaneously, which showed worse growth in milk than the ΔoppΔdppA_1_ΔdppA_2_ and ΔoppΔdppA_1_ΔdppA_3_ strains. Both titration acidity and viable cell counts in strains ΔoppΔdppA_1_ΔdppA_2_ΔdppA_3_ and ΔoppΔdppA_1_ΔdppA_3_ were significantly lower than those in ΔoppΔdppA_1_ but similar to those in the ΔoppΔdpp strain with deficient Dpp_Lc_ function (Fig. 7).
*Growth characteristics of candidate gene-disrupted strains of DppLc. Titration of acidity after 48 h of growth in milk medium. Error bars indicate sd (n=3). Asterisks indicate significant differences from the ΔoppΔdppA1 group assessed using Dunnett’s test; **P<0.001, n.s., P≥0.1. ∆opp, KO strain lacking oppLc genes; ΔoppΔdpp, KO strain lacking oppLc and dppLc genes; ΔoppΔdppA1, KO strain lacking oppLc and dppA1 genes; ΔoppΔdppA2, KO strain lacking oppLc and dppA2 genes; ΔoppΔdppA3, KO strain lacking oppLc and dppA3; ΔoppΔdppA4, KO strain lacking oppLc and dppA4; ΔoppΔdppA5, KO strain lacking oppLc and dppA5; ΔoppΔdppA6, KO strain lacking oppLc and dppA6.
*Growth characteristics of KO strains lacking DppA1, DppA2 or DppA3. Titration acidity (a) and viable cell count (b) after 48 h of growth in milk. Error bars indicate the sd (n=3) for each graph. Only comparisons of interest are indicated with brackets. ***P<0.001, *P<0.01. ΔoppΔdpp, gene KO strain lacking oppLc and dppLc; ΔoppΔdppA1ΔdppA2, KO strain lacking oppLc, dppA1 and dppA2; ΔoppΔdppA1ΔdppA3, KO strain lacking oppLc, dppA1 and dppA3; ΔoppΔdppA1ΔdppA2ΔdppA3, KO strain lacking oppLc, dppA1, dppA2 and dppA3.
Discussion
To investigate nitrogen utilization, particularly oligopeptide uptake, in LcS, we analysed the functions of oligopeptide transporters during growth in milk. The LcS genome contains two oligopeptide transporter-encoding operons, Opp_Lc_ and Dpp_Lc_, which exhibit high aa sequence homology with the Opp [9] and Dpp [18] transport systems in L. lactis, both of which are crucial for milk-based growth. In LcS, opp_Lc_ deficiency (Δopp) significantly impaired its proliferation in milk, whereas the simultaneous deletion of opp_Lc_ and dpp_Lc_ (ΔoppΔdpp) further reduced proliferation, indicating functional redundancy. The partial restoration of titratable acidity observed in Δopp^c^, the opp_Lc_-complemented strain, could be attributed to differences between the promoter used in the complementation vector pYAP300 and the native promoter of opp_Lc_ in the LcS WT strain, particularly in terms of expression timing and strength. Furthermore, ΔoppΔdpp also lost the ability to uptake 3–8 residue-long peptides. These findings suggest that Opp_Lc_ and Dpp_Lc_ are crucial oligopeptide transporters for LcS growth in milk, with absorptions of peptides with more than three residues serving as primary nitrogen sources. However, in MRS medium, which is rich in free aa, the ΔoppΔdpp strain showed growth comparable to that of the WT strain, indicating that it can proliferate adequately when aa are the primary nitrogen source. In addition, the inability of the ΔoppΔdpp strain to proliferate in milk is because of the scarcity of free aa in milk, making it reliant on the absorption of peptides with three or more residues as a sufficient nitrogen source. Our findings indicate that oligopeptides are the primary nitrogen source for LcS growth in milk, consistent with a report that L. lactis relies on oligopeptides for more than 90% of its nitrogen requirements during milk-based growth [10].
Our findings also revealed that Opp_Lc_ and Dpp_Lc_ differ in their transportable peptide lengths. Dpp_Lc_ transports peptides with 3–7 residues, whereas Opp_Lc_ transports peptides with 4–8 residues. This overlapping transport range suggests that Opp_Lc_ can compensate for most of the functions of Dpp_Lc_, which may explain why Δdpp showed no significant growth difference in milk compared with the WT strain. However, in this study, only a single peptide sequence was tested for each peptide length, limiting our understanding of how peptide sequence variability affects transport efficiency. Therefore, future studies are required to assess the effect of sequence variation on transporter function.
In L. lactis, the proton-coupled transporter, DtpT, is responsible for di- and tripeptide uptake [31]. In this study, we did not investigate dtpT. Nevertheless, the observation that simultaneous deletion of both Opp_Lc_ and Dpp_Lc_ (ΔoppΔdpp) completely abolished the ability to incorporate 3-residue peptides suggests that DtpT_Lc_ is not involved in the uptake of peptides with three or more residues. However, the possibility that DtpT_Lc_ contributes to dipeptide uptake cannot be excluded, which we intend to explore in future studies. Previous studies in L. bulgaricus B14 demonstrated Opp-mediated transport of 5-residue peptides only [19]. In this study, we showed that Opp_Lc_ in LcS can transport peptides up to eight residues, the longest tested peptide, making this the first direct evidence demonstrating the function of oligopeptide transporters in transporting peptides longer than six residues in lactobacilli. However, the ability of Opp_Lc_ to transport >8-residue-long peptides remains to be explored.
These findings align with those of previous studies on the Opp system in L. lactis MG1363, where Opp does not import 3-residue peptides but does facilitate the uptake of 4–8-residue peptides [9]. The functional similarity between Opp_Lc_ and L. lactis Opp, supported by their aa sequence homology, highlights their analogous roles. Similarly, the capacity of Dpp_Lc_ to transport 3-residue peptides, which Opp_Lc_ cannot, aligns with previous findings [9]. Conversely, the characteristics of the peptides assimilated via Dpp in L. lactis vary by strain, depending on the type of OBPs involved. For example, some strains, such as MG1363, express a Dpp optimized for di- and tripeptide uptake, whereas strains like IL1403, SK11, Wg2 and SKM6 possess a second peptide-binding protein, DppP, in addition to DppA, enabling the uptake of 2–9-residue peptides [5]. Our findings confirmed that Dpp_Lc_ exhibited an uptake capacity of up to seven residues, similar to the Dpp type, which is capable of transporting peptides longer than three residues. Nevertheless, further investigation is warranted to determine whether Dpp_Lc_-like oligopeptide transporters function in other L. paracasei strains and additional lactobacilli beyond LcS.
Analysis of the OppA_1_ KO strain characteristics revealed that Opp_Lc_ utilizes only one identified functional OBP, OppA_1_, under the tested conditions. This finding suggests that OppA_1_ is likely responsible for transporting 4–8 aa residues. However, the possibility of the existence of other Opp_Lc_-associated OBPs that function under alternative environmental or physiological conditions cannot be ruled out. Therefore, while our data support a central role for OppA_1_ in the tested context, further investigation is required to comprehensively elucidate its substrate range and regulatory mechanisms. In contrast, a comparative analysis of the DppA_1_ and Dpp_Lc_ gene-disruption strains revealed disparities in both proliferation in milk medium and the peptides available for uptake, indicating that Dpp_Lc_ possesses an OBP other than that of DppA_1_. We evaluated the proliferation in milk medium with five candidate genes that were potential OBPs of Dpp_Lc_ and confirmed the functional involvement of two of them, DppA_2_ and DppA_3_. Gene disruption, in which DppA_2_ and DppA_3_ were deleted simultaneously with DppA_1_, showed marginally better proliferation in milk than the gene disruption strain in which the function of the Dpp_Lc_ transport system was completely deleted. This suggests that DppA_2_ and DppA_3_ share certain peptides involved in transport and complement each other’s functions.
Moreover, we showed that dppA_2_ and dppA_3_, which are proposed to be OBPs for Dpp_Lc_, are genetically distant from the dpp_Lc_ operon. This pattern has also been observed in the Ami transport system, which is analogous to the Opp transport system of S. thermophilus [17], and in the Hpp transport system, which is equivalent to the Opp transport system of Streptococcus gordonii [32]. Together, these findings suggest that such a genomic configuration is a conserved feature among peptide transport systems in certain LAB species.
Collectively, our findings suggest that two oligopeptide transporters, Opp_Lc_, comprising five subunits OppA_1_BCDF, and Dpp_Lc_, comprising seven subunits DppA_1_A_2_A_3_BCDF, are crucial for nitrogen acquisition in LcS during milk growth. Furthermore, we showed that some oligopeptide transporters found in LcS resembled those in the phylogenetically distantly related L. lactis, highlighting the important role of oligopeptide transporters across diverse dairy-related strains. The expression of both transport systems is likely to broaden the range of peptide lengths that LcS can absorb. This phenomenon may represent a survival strategy for LcS, a probiotic lactic acid bacterium used in dairy production, to enhance peptide uptake efficiency, which is vital for nitrogen acquisition in milk. This study provides insights into the characteristics of peptides that Opp_Lc_ and Dpp_Lc_ can transport; however, the analysis was limited to the test peptides of eight residues or fewer. It remains unclear whether transport efficiency is influenced primarily by peptide length or specific aa motifs. Since the test peptides contained conserved residues such as Gly and Leu, future studies should investigate sequence-specific uptake preferences, including peptides of identical length but differing in primary sequence. We plan to conduct a more comprehensive analysis of the peptides taken up by oligopeptide transporters, particularly in the context of LcS growth in milk medium or other fermentation substrates.
Supplementary material
10.1099/mic.0.001624Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chopin A Organization and regulation of genes for amino acid biosynthesis in lactic acid bacteria FEMS Microbiol Rev 199312213710.1111/j.1574-6976.1993.tb 00011.x 8398216 · doi ↗ · pubmed ↗
- 2Morishita T Fukada T Shirota M Yura T Genetic basis of nutritional requirements in Lactobacillus casei J Bacteriol 19741201078108410.1128/jb.120.3.1078-1084.19744215789 PMC 245885 · doi ↗ · pubmed ↗
- 3Savijoki K Ingmer H Varmanen P Proteolytic systems of lactic acid bacteria Appl Microbiol Biotechnol 20067139440610.1007/s 00253-006-0427-116628446 · doi ↗ · pubmed ↗
- 4Kok J Leenhouts KJ Haandrikman AJ Ledeboer AM Venema G Nucleotide sequence of the cell wall proteinase gene of Streptococcus cremoris Wg 2Appl Environ Microbiol 19885423123810.1128/aem.54.1.231-238.19883278687 PMC 202426 · doi ↗ · pubmed ↗
- 5Doeven MK Kok J Poolman B Specificity and selectivity determinants of peptide transport in Lactococcus lactis and other microorganisms Mol Microbiol 20055764064910.1111/j.1365-2958.2005.04698.x 16045610 · doi ↗ · pubmed ↗
- 6Griffiths MW Tellez AM Lactobacillus helveticus: the proteolytic system Front Microbiol 201343010.3389/fmicb.2013.0003023467265 PMC 3587842 · doi ↗ · pubmed ↗
- 7Zhang H Xu M Hu S Zhao H Zhang B The enzyme gene expression of protein utilization and metabolism by Lactobacillus helveticus CICC 22171 Microorganisms 202210172410.3390/microorganisms 1009172436144326 PMC 9501481 · doi ↗ · pubmed ↗
- 8Rodríguez-Serrano GM Garcia-Garibay JM Cruz-Guerrero AE Gomez-Ruiz L del C Ayala-Nino A et al Proteolytic system of Streptococcus thermophilus J Microbiol Biotechnol 2018281581158810.4014/jmb.1807.0701730196594 · doi ↗ · pubmed ↗