Identification and Validation of a New Functional Gene TSC22D3 for hBMSCs Osteogenesis
Shuhong Zhang, Xiao Li, Zhanping Yang, Jialong Li, Gang Liu, Yongkun Sun, Huigen Feng, Xianwei Wang

TL;DR
This study identifies TSC22D3 as a key gene promoting bone formation by regulating osteogenic differentiation in stem cells.
Contribution
The novel contribution is the identification of TSC22D3 as a core regulator of osteogenic differentiation and its role in cytoskeletal rearrangement.
Findings
TSC22D3 is significantly upregulated during osteogenic differentiation and promotes osteogenic marker expression.
Downregulation of TSC22D3 reduces ALP activity, calcium deposition, and osteogenic marker genes like Runx2 and OCN.
TSC22D3 influences cytoskeletal rearrangement and is linked to WNT signaling and cell cycle pathways.
Abstract
Osteogenic differentiation is a crucial process in which bone marrow mesenchymal stem cells (BMSCs) differentiate into osteoblasts, involving the regulation of multiple genes and signaling pathways. The TSC22D3 gene plays an important role in various biological processes (BPs), but its specific function in osteogenic differentiation remains unclear. This study aims to explore the regulatory role of the TSC22D3 gene in osteogenic differentiation and its molecular mechanisms. By analyzing microarray datasets (GSE12266, GSE18043, and GSE80614), the limma package was used to screen for differentially expressed genes (DEGs). Combined with Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses, key genes and signaling pathways related to osteogenic differentiation were identified. Further, through protein–protein interaction (PPI) network analysis and the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
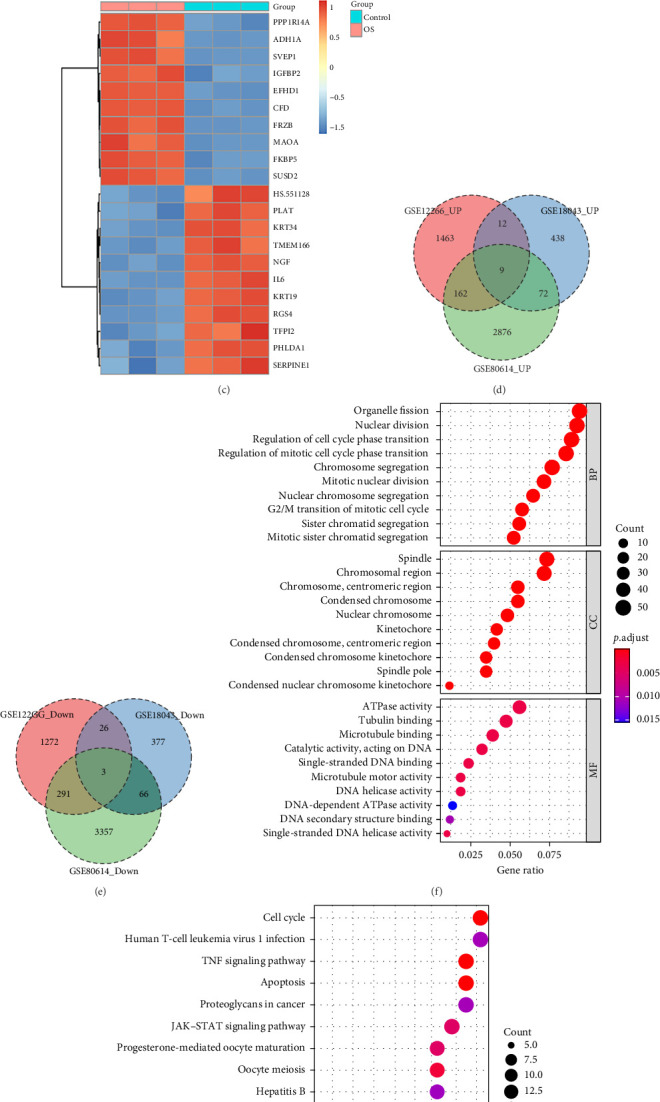 Figure 1
Figure 1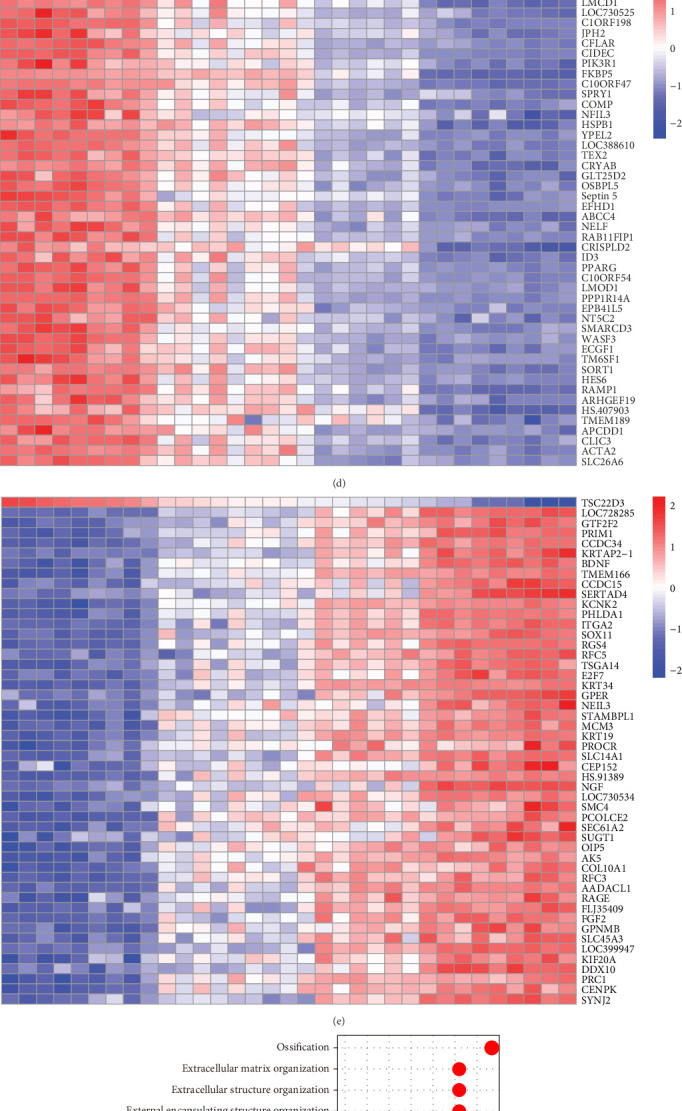 Figure 2
Figure 2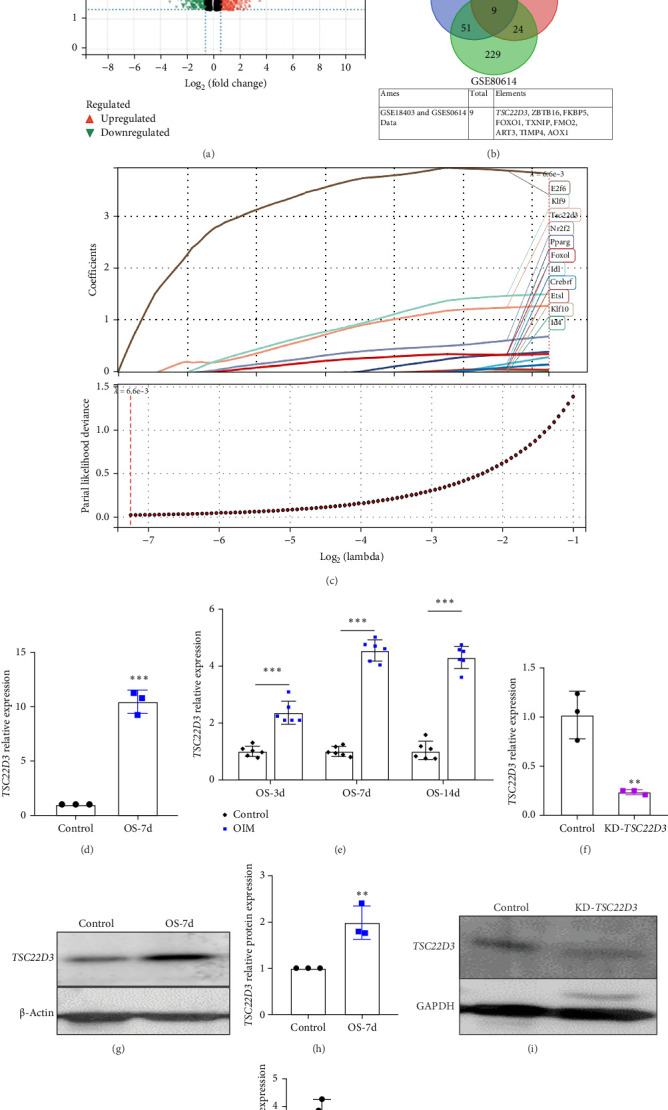 Figure 3
Figure 3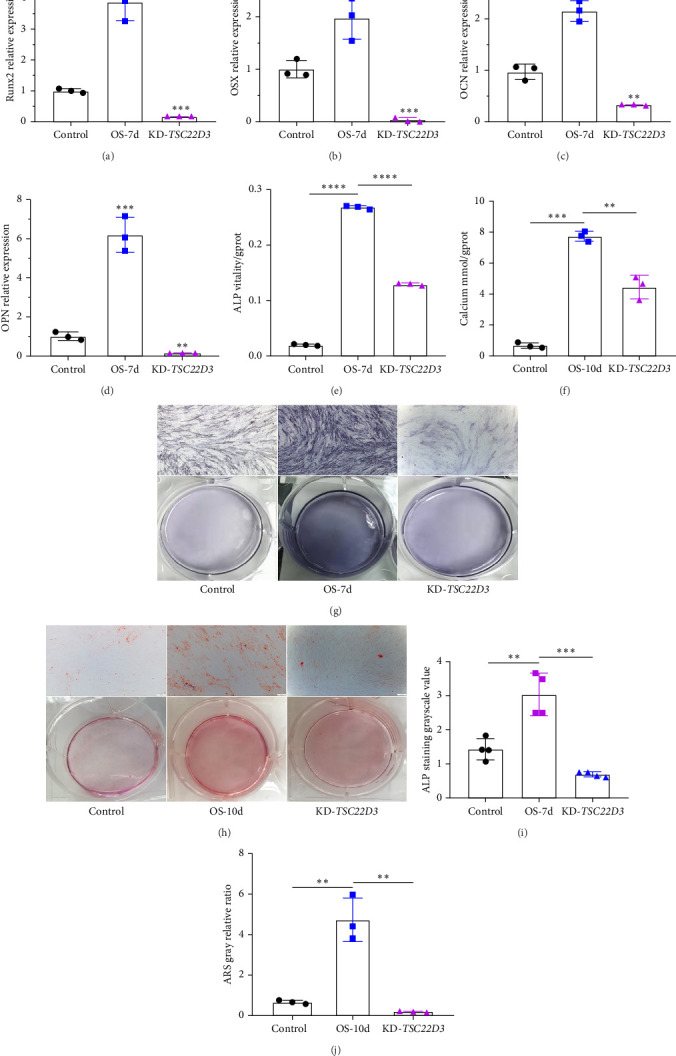 Figure 4
Figure 4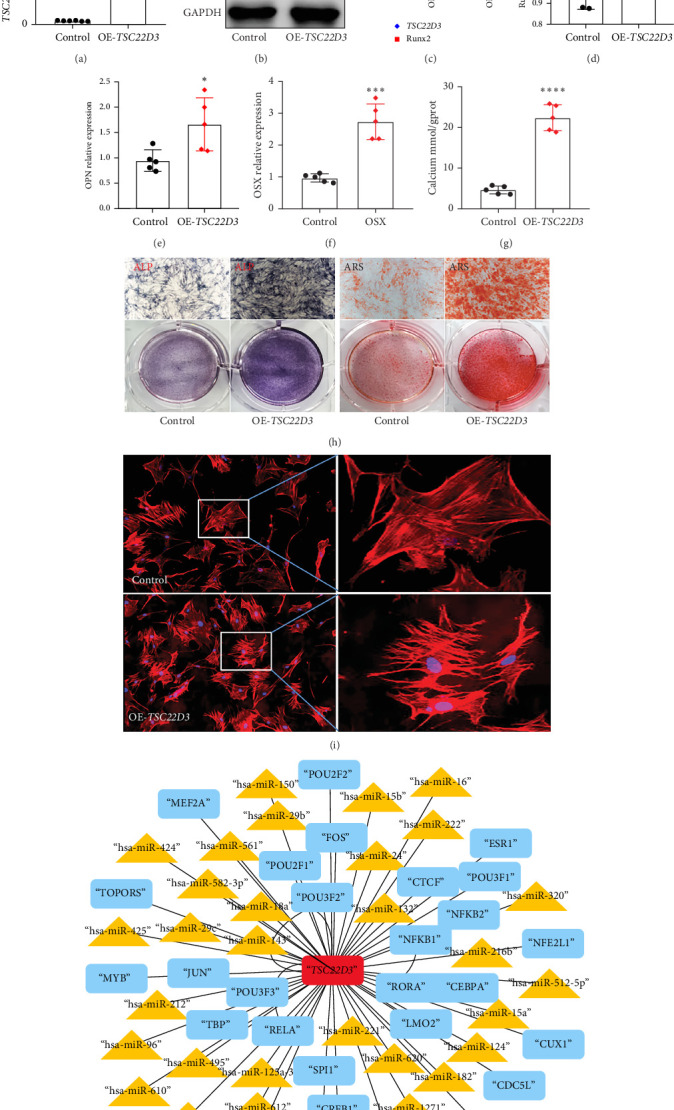 Figure 5
Figure 5- —Scientific and Technological Innovative Teams in Universities of Henan Province
- —Xinxiang Medical University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHeterotopic Ossification and Related Conditions · Connective tissue disorders research · Bone Metabolism and Diseases
1. Introduction
Skeletal pathologies, such as osteoporosis and osteoarthritis, involve complex processes that affect bone formation, remodeling, and function, ultimately leading to severe functional impairment [1]. Bone marrow mesenchymal stem cells (BMSCs) can self-renew and differentiate into osteoblasts, chondrocytes, and adipocytes, making them valuable for clinical applications [2]. BMSCs play a crucial role in bone repair through osteogenic differentiation, which is regulated by complex gene networks and signaling pathways [3]. In vivo studies have shown that stem cell fate and differentiation are influenced by factors such as the matrix microenvironment, scaffold composition, and cytokines [4]. Understanding the molecular mechanisms controlling osteogenic differentiation is essential for optimizing bone scaffold transplantation and improving bone injury repair [5]. While high-throughput sequencing has helped deepen our understanding of osteogenic differentiation, there remains a limitation in integrating data to identify key regulatory genes, and this hinders our comprehensive understanding of the relevant molecular mechanisms.
The TSC22D3 gene encodes a glucocorticoid-induced leucine zipper (GILZ), which plays an important role in the anti-inflammatory and immunosuppressive responses [6]. This protein can inhibit proinflammatory molecules and is involved in various biological processes (BPs) such as immune regulation and cell proliferation [7]. Recent research has expanded our understanding of TSC22D3 and explored its role in different diseases and conditions. For example, it has been identified as an immune-related prognostic biomarker in acute myeloid leukemia (AML) [8], and its expression is associated with patient prognosis and tumor microenvironment [9]. In addition, the study also demonstrated the involvement of TSC22D3 in stress response, particularly in the context of cancer. The study suggests that TSC22D3 may influence cell fate determination by regulating transcription factors or epigenetic modifications [10]. These findings highlight the different roles of TSC22D3 in health and disease, but its specific mechanism in BMSCs osteogenic differentiation still needs further investigation. Additionally, current osteogenic differentiation-related gene screening often relies on single datasets, which may introduce technical bias, and there is a need for multidataset joint analysis to improve the reliability of the results.
This study identifies key osteogenic differentiation genes through integrated analysis of microarray datasets (GSE12266, GSE18043, and GSE80614) and our transcriptome sequencing data. Using differentially expressed genes (DEGs) screening, enrichment analysis, and a protein–protein interaction (PPI) networks, we systematically mapped BPs and identified hub genes. Functional validation of TSC22D3 was performed through lentiviral interference, RT-qPCR, Western blotting, and osteogenic assays in BMSCs. These research findings deepen the understanding of osteogenic mechanisms and provide potential therapeutic targets for bone diseases.
2. Materials and Methods
2.1. Data Processing and Analysis
2.1.1. DEG Screening and Gene Enrichment Analysis
DEGs were identified from microarray datasets (GSE12266, GSE18043, and GSE80614) using the limma package (Supporting Information File 1). To address false discovery rates in multiple testing, adjusted p-values were applied. DEGs distinguishing osteogenic differentiation samples from normal controls were selected using stringent thresholds (adjusted p < 0.05 and log2 fold-change ≥0.5). Genes showing consistent upregulation or downregulation in at least two datasets were prioritized (Supporting Information File 2). Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed on these DEGs using the clusterProfiler package in R.
2.1.2. Identification and Analysis of Hub Genes in Osteogenic Differentiation
Hub genes and regulatory networks were identified through integrated bioinformatics approaches. First, the Finding Regulatory Elements by Differential Expression and Network-Based Statistical Analysis (FRIEND) method was applied using the GOSemSim package in R to select the top 50 candidate genes based on GO enrichment results (Supporting Information File 3). Concurrently, a PPI network was constructed using DEGs via the STRING database. The cytoHubba plugin in Cytoscape was used to identify the top 50 PPI network genes. Intersection of these two gene lists yielded final hub genes, with high-scoring candidates selected for experimental validation (Supporting Information File 4). Subsequent analyses of the hub genes included correlation analysis, GO, KEGG, and Gene Set Enrichment Analysis (GSEA) (Supporting Information File 5).
The results of the aforementioned bioinformatics analysis were intersected with the transcriptomic sequencing data from our previous experiments on osteogenic differentiation induction in rat BMSCs, to further identify the core molecules closely associated with osteogenic differentiation.
2.2. Main Materials and Reagents
Human BMSCs (hBMSCs) were obtained from Saiye Biotechnology Co., Ltd. (product number: HUXMA-01001). Lentivirus-mediated interference plasmid for downregulation of TSC22D3 was custom-synthesized by Anshengda Biotechnology Co., Ltd. (order number: 80-839735734). Osteogenic induction medium (OIM), consisting of the following components: Dulbecco's Modified Eagle Medium (DMEM), 10% fetal bovine serum (FBS), 10 mM β-glycerophosphate, 50 μg/mL L-ascorbic acid, 0.1 μmol/L dexamethasone, and 1% double antibiotic. Alkaline phosphatase (ALP) assay kit (Nanjing Jiancheng Bioengineering Institute, product number: A059-2); calcium assay kit (Nanjing Jiancheng Bioengineering Institute, product number: C004-2-1); TSC22D3 antibody (Proteintech, product number: 12352-1-AP).
2.3. Main Experimental Methods
2.3.1. Cell Culture and Osteogenic Induction
hBMSCs were maintained in minimum essential medium alpha (α-MEM) supplemented with 10% FBS at 37°C in a 5% CO_2_ humidified incubator. Cells at passages 3–5 were used for experiments. Osteogenic differentiation was induced by culturing hBMSCs in OIM medium for the required experimental duration.
2.3.2. Transcriptome Sequencing Analysis
The research team previously extracted BMSCs from rats and conducted primary culture. Cells from the P4 generation were selected and induced to undergo osteogenic differentiation in OIM for 7 days. RNA was then extracted and sent to a company (Shanghai Paisenuo Biotechnology Co., Ltd., contract number: PN20241127035) for transcriptome sequencing. The samples were categorized into two groups: a control group and an osteogenic differentiation group (OS) (Supporting Information File 6). Subsequently, the sequencing results underwent comprehensive bioinformatics analysis.
2.3.3. Lentiviral Interference
Lentiviral packaging was performed in 293T cells using the target shRNA, psPAX2, and Pmg2G plasmids at a ratio of 6:4:2. After 48–60 h of transfection, the supernatant from the 293T cells was collected. Each well containing hBMSCs was supplemented with 1 mL of the 293T supernatant, followed by the addition of 1 mL of OIM medium to each well. Finally, approximately 8 μg/mL polybrene solution was added to each well. The wells were gently mixed, and 4–6 h after transfection, the medium was replaced with fresh OIM.
2.3.4. RT-qPCR and Western Blotting
In this study, the expression of hub genes and osteogenic marker genes was identified by RT-qPCR and Western blotting. The primer sequences used in this experiment were listed in Table 1. The experimental results were plotted and statistically analyzed using Primer 8.0 software.
2.3.5. Osteogenic Differentiation Related Assays
ALP staining was assessed using the BCIP/NBT ALP chromogenic reagent kit, and ALP activity was measured using an ALP assay kit. Calcium deposition in hBMSCs was evaluated through Alizarin Red (ARS) staining and a calcium assay kit. The experimental results were analyzed for grayscale using Image Proplus 6.0 software.
2.3.6. Cytoskeleton Staining
Use a phalloidin kit to stain the cytoskeleton. First, place the target adherent cells (cultured to 50%–80% confluency) on a coverslip. After aspirating the culture medium, fix the cells with precooled 4% paraformaldehyde at room temperature for 10–15 min. Next, permeabilize the cells with PBS containing 0.1%−0.5% Triton X-100 at room temperature for 5–10 min, followed by blocking with PBS containing 1%–5% bovine serum albumin (BSA) at room temperature for 30 min. Subsequently, dilute the fluorescently labeled phalloidin in proportion, cover the cells with it, and incubate in the dark at room temperature for 20–30 min. Then, add DAPI staining solution and incubate in the dark at room temperature for 5–10 min. After each incubation step, wash the cells with PBS three times (5 min each time). Finally, mount the sample with antifluorescence quenching mounting medium and perform scanning imaging.
2.3.7. Statistics
Statistically significant differences between means were determined using the GraphPad Prism 8.0 software. Unless stated otherwise, an unpaired Welch's t test was performed for the comparison of two groups. Results were considered significant at p < 0.05.
3. Results
3.1. Identification and Analysis of DEGs During Osteogenic Differentiation Process
Heatmaps (Figure 1A–C) were employed to highlight the most significant genes among all DEGs within the OS in comparison to the control group. Conduct an intersection analysis of DEG within the three datasets. Intersection analysis identified nine upregulated genes (Figure 1D) and three downregulated genes (Figure 1E). Subsequently, prioritize genes that exhibit concurrent upregulation or downregulation in at least two of the datasets for further in-depth analysis.
We performed GO analysis on DEGs to identify key functional aspects of genes in osteogenic differentiation (Figure 1F). Key findings included BPs like organelle fission, nuclear division, and cell-cycle phase transition regulation. Cellular components (CCs) such as the spindle and chromosomal regions were also highlighted. Molecular functions (MFs) linked to osteogenic differentiation included ATPase activity and tubulin/microtubule binding. We performed KEGG pathway analysis to identify signaling pathways involved in the osteogenic differentiation of hBMSCs, revealing 294 significant pathways (p < 0.05). Key pathways included those related to the cell cycle, human T-cell leukemia virus 1 infection, TNF signaling, apoptosis, and proteoglycans in cancer (Figure 1G). These results enhance our understanding of the molecular mechanisms underlying osteogenic differentiation in hBMSCs.
3.2. Screening and Bioinformatics Analysis of Hub Gene TSC22D3
To identify hub genes, we selected the top 50 candidate genes using the FRIENDS method (Figure 2A), including TROAP, FANCA, and TSC22D3, and the top 50 key genes from PPI analysis (Figure 2B), such as IL6, BRCA1, CCNB1, and VEGFA. Intersection analysis of these genes revealed four hub genes: TSC22D3, BIRC5, GMNN, and BRCA1 (Figure 2C). TSC22D3, with the highest FRIENDS score, was identified as the primary hub gene. Correlation analysis showed genes positively and negatively associated with TSC22D3 (Figure 2D, E), indicating functional links.
GO analysis of TSC22D3 highlighted its role in osteogenic differentiation (Figure 2F), while pathway analysis linked it to human papillomavirus infection, cytoskeleton regulation, and WNT signaling (Figure 2G). GSEA using reactome pathways (Figure 2H) further revealed subtle yet significant gene expression changes, aiding in understanding upstream and downstream regulatory mechanisms.
3.3. Experimental Verification of TSC22D3 Expression and Interference
In previous studies, our research group has completed transcriptome sequencing of a rat BMSC osteogenic induction model. The volcano plot (Figure 3A) and bar chart (Figure 3D) demonstrate that TSC22D3 showed significantly higher expression in osteogenically induced cells compared to the control group in these transcriptomic sequencing results. The Venn diagram (Figure 3B) reveals the intersection of upregulated DEGs between our transcriptome sequencing data and two datasets (GSE18403 and GSE80614), identifying nine coupregulated genes, including TSC22D3. LASSO analysis of the sequencing results (Figure 3C) further indicated that TSC22D3 ranked prominently among the top regression coefficients (top 3, coefficient = 1.28). These experimental findings collectively suggest that the TSC22D3 gene may play a significant role in osteogenic differentiation processes.
To validate the findings from our bioinformatics analysis, we examined both mRNA and protein expression levels of TSC22D3 in hBMSCs. The RT-qPCR results demonstrated a significant upregulation of TSC22D3 expression in cell samples undergoing osteogenic differentiation at 3, 7, and 14 days compared to the control group (day 0) (Figure 3E). Western blotting analysis (Figure 3G, H) further confirmed that TSC22D3 protein expression increased following osteogenic induction.
In this study, we utilized lentiviral transduction to knockdown (KD) TSC22D3 expression in hBMSCs. To validate the KD efficiency, we performed comprehensive assessments using both RT-qPCR and Western blotting analyses, as shown in Figure 3F, I, J.
3.4. The TSC22D3 Gene Plays a Crucial Role in the Osteogenic Differentiation of hBMSCs
After downregulating the expression of TSC22D3 gene, we evaluated the expression levels of key osteogenic marker genes runt-related transcription factor 2 (Runx2), osterix (OSX), osteocalcin (OCN), and osteopontin (OPN) using RT-qPCR technology, and detected ALP activity and calcium deposition. Contrast to the control group, the KD of TSC22D3 (KD-TSC22D3) resulted in a substantial reduction in the expression of osteogenic marker genes (Figure 4A–D), a concomitant decrease in ALP activity (Figure 4E), and a discernible diminishment in calcium deposition (Figure 4F). Collectively, these experimental findings lend credence to the hypothesis that the TSC22D3 gene exerts a regulatory influence over the osteogenic differentiation process in hBMSCs.
ALP and ARS staining assays were carried out on cellular specimens derived from the control group, the OS-7d group, and the KD-TSC22D3 group, as depicted in Figure 4G–J. Notably, within the KD-TSC22D3 cell population, a conspicuous reduction in both ALP content and calcium deposition was observed. This compelling observation implies a pivotal role of the TSC22D3 gene in modulating the osteogenic differentiation process of hBMSCs, thereby underscoring the need for further experimental validation to elucidate the precise underlying mechanisms.
3.5. Overexpressing TSC22D3 Promotes hBMSCs Osteogenic Differentiation and Cytoskeleton Rearrangement
To further verify the correlation between TSC22D3 and the osteogenic differentiation of BMSCs, we overexpressed TSC22D3 in hBMSCs via lentiviral packaging and transfection, and verified the indicators of osteogenic differentiation. First, RT-qPCR and Western blot were used to detect the efficiency of TSC22D3 overexpression (OE). Compared with the control group, the mRNA and protein levels of the target gene TSC22D3 in the OE group (OE-TSC22D3) were significantly upregulated (Figure 5A–C). Meanwhile, it was detected that after OE-TSC22D3, the expression of Runx2 protein was upregulated (Figure 5B,C). We evaluated the expression levels of key osteogenic marker genes Runx2, OPN, and OSX using RT-qPCR technology, and detected calcium deposition. Compared with the control group, OE-TSC22D3 resulted in an increase in the expression of osteogenic marker genes (Figure 5D–F), and an increase in calcium deposition (Figure 5G). In ALP and ARS staining assays, enhanced ALP and ARS staining was observed in the OE-TSC22D3 group compared with the control group (Figure 5H). All these experimental results suggest that TSC22D3 can promote the osteogenic differentiation of hBMSCs.
Since previous bioinformatics analysis showed that TSC22D3 is closely related to the cytoskeletal system (Figure 2G, H), we used phalloidin to stain the cytoskeleton of hBMSCs with OE-TSC22D3. The results, as shown in Figure 5I, revealed structural rearrangement of the cytoskeleton, with a visible increase in the proportion of clustered and longitudinally arranged cytoskeletal structures. This result further indicates that TSC22D3 may be involved in the osteogenic differentiation process by altering the cytoskeletal structure. In addition, through bioinformatics analysis, we screened potential transcription factors and miRNAs that may regulate TSC22D3 (Figure 5J), which further suggests its possible molecular mechanism.
4. Discussion
This study systematically explored the regulatory role of TSC22D3 gene in the osteogenic differentiation of hBMSCs by integrating bioinformatics analysis and experimental verification. The research results revealed the potential mechanism of TSC22D3 as a key hub gene in osteogenic differentiation, and its importance was verified through functional experiments, providing a new perspective for a deeper understanding of the molecular regulatory network of osteogenic differentiation.
Chronic diseases or injuries that damage bone tissue can disrupt daily life and complicate treatment. Repair often requires in vitro scaffolds or composite structures with seed cells [11]. The use of hBMSCs in tissue-engineered bone repair has gained attention [12]. While stem cells hold promise for tissue repair, their regenerative potential is influenced by complex molecular networks, and the underlying mechanism remains unclear.
Research on stem cell differentiation and regeneration now focuses on understanding the cells and signaling molecules in the local matrix microenvironment [13]. Signaling regulation is key to osteogenic differentiation, but the exact mechanisms behind osteogenesis, tissue regeneration, and repair remain unclear, limiting clinical applications. BMSCs are typically induced to become osteoblasts under certain conditions. Studies have highlighted the role of key signaling pathways, such as TGF-β/BMP [14], WNT [15], and FGF [16], in regulating BMSCs differentiation and lineage commitment [17]. The biological modules and core genes identified through experimental validation may ultimately serve as biomarkers for the detection and treatment of bone injury [18].
This study identified the pivotal gene TSC22D3, which is significantly upregulated during osteogenic differentiation, through a joint analysis of multiple datasets. Experimental validation showed that the expression of TSC22D3 was significantly increased at day 3, 7, and 14 of osteogenic induction. KD-TSC22D3 significantly inhibited the expression of osteogenic marker genes (such as Runx2, OSX, OCN, and OPN), and decreased aALP activity and calcium deposition levels. OE-TSC22D3; however, promotes osteogenic differentiation. This finding provides the first evidence of the positive regulatory role of TSC22D3 in osteogenic differentiation of hBMSCs, filling the gap in the study of this gene in bone metabolism research.
Our GO and KEGG analyses revealed several interconnected BPs related to BMSCs osteogenic differentiation, including organelle fission, nuclear division, and cell-cycle phase regulation. Notably, signal regulation governing cell cycle phase transitions has implications in various biological phenomena such as obesity, hyperinsulinemia, hormonal regulation, the local microenvironment, and cellular aging [19–21]. Our biological analysis identified fluid shear stress as a factor influencing osteogenic differentiation. Studies indicate that fluid shear stress regulates vascular tension and blood pressure via endothelial adrenomedullin signaling [22, 23]. This discovery indirectly suggests that there may be some network like regulatory relationships between abnormal regulation of osteogenic differentiation and the occurrence and development of a series of diseases in the body, which can be further explored.
GO and KEGG analyses of the hub gene TSC22D3 showed that this gene is closely related to osteogenic differentiation regulation, bone mineralization, cytoskeleton regulation, WNT signaling pathway, and TGF-β signaling pathway. In addition, LASSO regression analysis and GSEA results suggest that TSC22D3 may exert its effects through synergistic or independent pathways, complementing the regulatory network of classical osteogenic transcription factors such as Runx2 and OSX recently discovered [24]. The experimental results also confirmed that TSC22D3 can induce cytoskeletal rearrangement. The above experimental results further support the important role of TSC22D3 in the regulation of osteogenic differentiation. However, the specific regulatory mechanism of TSC22D3 within these pathway remains to be further elucidated.
TSC22D3 is a protein encoding member of the TSC-22/DIP/Bun family, as its role in certain diseases has also received attention in recent years. Especially in glucocorticoid signaling, TSC22D3 is a key regulatory molecule [25]. Corticosteroids play an important role in treating certain bone diseases, such as rheumatoid arthritis, but long-term or high-dose use of glucocorticoids can inhibit bone formation, increase the risk of osteoporosis, and fractures. TSC22D3 is induced to express under the action of glucocorticoids and may indirectly affect bone metabolism by regulating inflammatory responses, cell proliferation, and other processes, especially in regulating bone diseases caused by glucocorticoids such as osteoporosis [26]. Studies have shown that upregulation of TSC22D3 can alleviate glucocorticoid related bone damage by improving the function of osteoblasts and reducing bone resorption [27]. Researchers may explore its potential as a drug target. In patients with long-term use of glucocorticoids, the expression of TSC22D3 may become an indicator for evaluating the risk of osteoporosis, helping doctors assess bone health status and take corresponding intervention measures.
In addition, the downregulation and loss of TSC22D3 in vitro and in vivo contributes to the aging process of macrophages and peripheral blood leukocytes [28]. The decrease in TSC22D3 mRNA and protein levels is associated with an increase in inflammation severity in systemic lupus erythematosus (SLE), ulcerative colitis, psoriasis, and other autoimmune inflammatory diseases [29]. It is worth noting that TSC22D3 exhibits enhanced expression during T cell transformation and has clinical relevance in tumor staging [30]. In the bioinformatics analysis of cervical cancer-related genes, TSC22D3 has significant features in the cell proliferation signaling system [31]. TSC22D3 is highly expressed in the brain tissue of morphine tolerant mice, activating the iron death pathway, enhancing cell apoptosis and promoting brain cell inflammatory response [32]. The expression of TSC22D3 in liver biopsy samples of patients with liver fibrosis is negatively correlated with the expression of CCL2 [33].
In conclusion, the TSC22D3 gene plays a crucial role in a variety of physiological and pathological processes, and it has attracted increasing attention as a potential therapeutic target. Although the TSC22D3 gene is associated with multiple diseases, its connection to osteogenesis has been rarely studied. Through analysis and experimental verification, this study confirmed the association between TSC22D3 and osteogenic differentiation, laying a foundation for in-depth exploration of the molecular mechanism and functional role of this gene in osteogenic differentiation.
This section discusses the strengths and weaknesses of the methodology used in the study. The research applied a thorough bioinformatics approach (e.g., limma package for gene screening and STRING database for PPI network construction), alongside experimental validation (e.g., RT-qPCR, Western blot, and phenotype detection), to ensure result reliability. However, there are some limitations: the specific downstream targets of TSC22D3 and their relationship with signaling pathways remain unclear, requiring further exploration using techniques like ChIP-seq or dual luciferase reporter assays.
Future research on TSC22D3 can focus on: (1) exploring its regulatory network and interactions with osteogenic pathways like WNT, TGF-β, and BMP; (2) creating a conditional knockout mouse model to study its role in bone formation and regeneration; (3) investigating its epigenetic regulation, such as methylation or noncoding RNA; (4) developing molecular interventions targeting TSC22D3 for potential applications in osteoporosis or bone defect treatment.
5. Conclusions
The identified gene TSC22D3 is a key regulator in osteogenic differentiation, which provides a novel biological insight in osteogenic differentiation of BMSCs. This study reveals that TSC22D3 is essential for osteogenic differentiation. Its upregulation promotes osteogenic marker expression, ALP activity, and calcium deposition, while its downregulation inhibits these processes. TSC22D3 affects cytoskeletal rearrangement during osteogenesis. The research findings of this article provide a potential therapeutic target for bone injury repair and in vitro bone tissue engineering.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhang Q. Dong J. Zhang P. Zhou D. Liu F. Dynamics of Transcription Factors in Three Early Phases of Osteogenic, Adipogenic, and Chondrogenic Differentiation Determining the Fate of Bone Marrow Mesenchymal Stem Cells in Rats Frontiers in Cell and Developmental Biology 2021910.3389/fcell.2021.768316768316 PMC 857656834765608 · doi ↗ · pubmed ↗
- 2Kenswil K. J. G. Pisterzi P. Sánchez-Duffhues G. Endothelium-Derived Stromal Cells Contribute to Hematopoietic Bone Marrow Niche Formation Cell Stem Cell 2021284653670.e 1110.1016/j.stem.2021.01.00633561425 · doi ↗ · pubmed ↗
- 3Cao S.-S. Li S.-Y. Geng Y.-M. Prefabricated 3D-Printed Tissue-Engineered Bone for Mandibular Reconstruction: A Preclinical Translational Study in Primate ACS Biomaterials Science & Engineering 20217125727573810.1021/acsbiomaterials.1c 0050934808042 PMC 8672350 · doi ↗ · pubmed ↗
- 4Li Y. Y. Choy T. H. Ho F. C. Chan P. B. Scaffold Composition Affects Cytoskeleton Organization, Cell–Matrix Interaction and the Cellular Fate of Human Mesenchymal Stem Cells Upon Chondrogenic Differentiation Biomaterials 20155220822010.1016/j.biomaterials.2015.02.0372-s 2.0-8493263534925818427 · doi ↗ · pubmed ↗
- 5Wang J. Liu S. Li J. Zhao S. Yi Z. Roles for mi RN As in Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells Stem Cell Research & Therapy 201910110.1186/s 13287-019-1309-72-s 2.0-85068114948197 PMC 659937931253175 · doi ↗ · pubmed ↗
- 6Nataraja C. Flynn J. Dankers W. GILZ Regulates Type I Interferon Release and Sequesters STAT 1 Journal of Autoimmunity 202213110.1016/j.jaut.2022.10285810285835810690 · doi ↗ · pubmed ↗
- 7Newton R. Anti-Inflammatory Glucocorticoids: Changing Concepts European Journal of Pharmacology 201472472423123610.1016/j.ejphar.2013.05.0352-s 2.0-8489372512723747654 · doi ↗ · pubmed ↗
- 8Li Y. Huang H. Zhu Z. Chen S. Liang Y. Shu L. TSC 22D 3 as an Immune-Related Prognostic Biomarker for Acute Myeloid Leukemia i Science 202326810.1016/j.isci.2023.107451107451 PMC 1041593137575189 · doi ↗ · pubmed ↗