Keeping cool: A brassinosteroid receptor BRL3 helps Arabidopsis cope with heat stress
Gunjan Sharma, Akanksha Bhatnagar

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
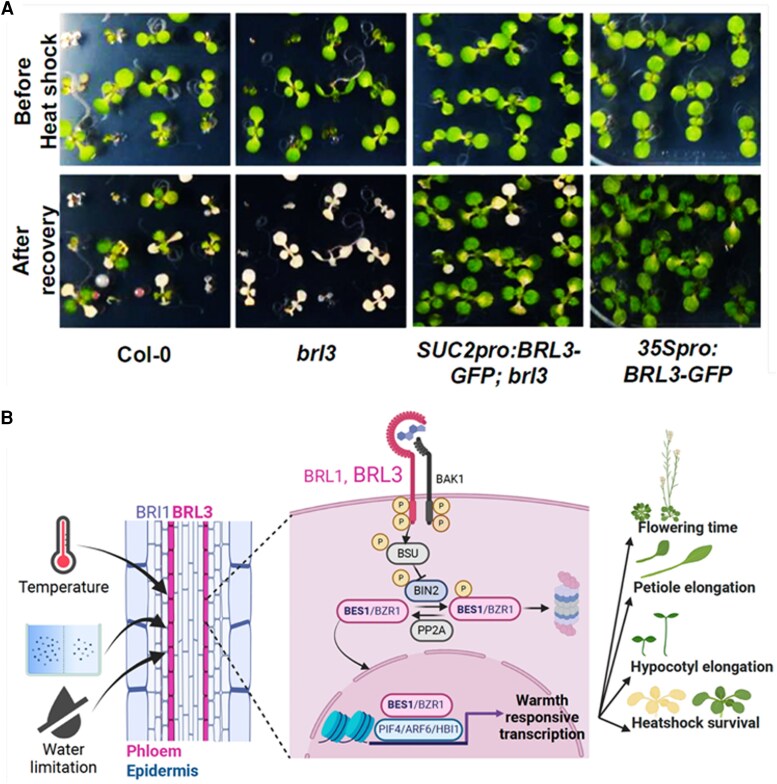 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Gene Expression Analysis · Plant Stress Responses and Tolerance
Climate change has increased the occurrence of frequent and intense heat waves imposing heat stress on plants and reducing crop productivity (Zhu et al. 2019; Cohen and Leach 2020). Plants naturally adapt to heat stress through thermomorphogenesis; elongating hypocotyls, roots, and petioles; and inducing early flowering and leaf hyponasty (Casal and Balasubramanian 2019). Molecular mechanisms governing thermomorphogenesis are well reported and involve the sensing of heat stress followed by signal transduction and adaptive response. Interestingly, plants also retain a memory of moderate heat exposure through epigenetic modifications, as reviewed by Staacke et al. (2025). However, the role of cell-specific sensing and signaling of heat stress is not yet fully understood.
A plasma membrane–localized leucine-rich-repeat receptor like kinase BRASSINOSTEROID-INSENSITIVE1 (BRI1) binds brassinosteroid (BR) and interacts with a co-receptor BRI1 ASSOCIATED RECEPTOR KINASE 1 (BAK1), activating downstream BR-mediated signaling. The roles of BRI1 as a growth promoter are well characterized in Arabidopsis. Interestingly, 2 other homologs of BRI namely BRI1-LIKE 1 (BRL1) and BRI1-LIKE 3 (BRL3) are predominantly expressed in vascular cells compared with BRI1, which exhibits ubiquitous expression (Cano-Delgado et al. 2004). Overexpression of BRI1 leads to drought tolerance but at the cost of a growth penalty. Surprisingly, overexpression of BRL3 confers drought tolerance without hampering plant growth (Fabregas et al. 2018). BRL3 is certainly a better candidate for engineering drought-tolerant plants because it uncouples reduced growth from drought tolerance. But how BRL3 behaves differently from its ubiquitous homolog BRI1 is still unknown.
In a recently published article in Plant Physiology, Rico-Medina et al. (2025) revealed that vascular localized BRL3 regulates heat stress responses. The brl3 mutant exhibited defective thermomorphogenesis and reduced survival during heat stress exposure. Phloem-specific complementation of brl3 mutant with wild-type (WT) BRL3 rescued the heat stress–adaptive developmental plasticity. BRL3 activates the BRI1-EMS-SUPRESSOR (BES1) transcription factor, regulating growth and development necessary for heat stress acclimation (Fig.).
The authors functionally characterize the brl3 mutant. The brl3 mutant was phenotypically similar to WT Arabidopsis at an ambient temperature of 22 °C. However, brl3 mutant exhibited a short hypocotyl and impaired petiole elongation when exposed to elevated temperature (28 °C) due to a reduction in cell length. This contrasts with bri1, which exhibits a short hypocotyl both at 22 °C and 28 °C. Interestingly, the transcript levels of BRI1 and other BRLs were unchanged in brl3 mutant background during heat stress, indicating a distinct BRL3-mediated signaling pathway to regulate thermomorphogenic responses. The authors showed that a downstream gain-of-function bes1 mutant rescued the growth defects of brl3 mutant at high temperature, suggesting that BRL3 uses the same downstream pathway as BRI1. A reduced de-phosphorylation of BES1 was also observed in brl3. BES1 localizes to nucleus in response to BR and regulates transcription of multiple genes involved in elongation growth. A BRASSINOSTEROID-INSENSITIVE 2 (BIN2) kinase negatively regulates the BR signaling pathway by phosphorylating and destabilizing BES1 (Yin et al. 2002).
Furthermore, RNA transcriptome sequencing of brl3 and WT shoots at 22 °C vs. at high temperature (28 °C) revealed that BRL3 targets the core thermomorphogenesis genes such as HEAT-SHOCK TRANSCRIPTION FACTOR A7A (HSFA7A), TEMPERATURE-INDUCED LIPOCALIN (TIL), and MULTIPROTEIN BINDING FACTOR 1C (MBF1C) at an elevated temperature. The brl3 mutant was unable to induce abiotic stress-responsive genes, while over-induction of negative regulators of ethylene, carboxylic acids, and carbon-catabolic processes was observed. Carboxylic acid intermediates of the tricarboxylic acid cycle are diverted toward generating protective compounds and signaling molecules during abiotic stresses instead of catabolism-mediated energy production (Antonio et al. 2016).
The authors further investigated the spatiotemporal contribution of BRL3 in regulating elevated temperature stress responses using tissue-specific promoters in brl3 mutant background for complementation analysis. BRL3-GFP expression under stele- (WOLpro:BRL3–GFP) and phloem companion cell-specific (SUC2pro:BRL3–GFP) promoters rescued brl3 mutant developmental defects, while phloem-pole pericycle (CALS8pro:BRL3–GFP) and protophloem sieve elements (NAC86pro:BRL3–GFP) specific expression conferred partial rescue at 28 °C. The pericycle is a layer of cells surrounding the procambium cells, xylem, and phloem, and can be distinguished into 2 cell types based on their position relative to xylem and phloem: the xylem pole pericycle and phloem pole pericycle (Zhang et al. 2022). Interestingly, epidermis (atrichoblasts: GL2pro:BRL3–GFP and trichoblasts: EXP7pro:BRL3–GFP), endodermis (SCRpro:BRL3–GFP), root stem cell niche (WOX5pro:BRL3–GFP), and root apical meristem (RPS5Apro:BRL3–GFP) specific promoters failed to rescue, suggesting an inner phloem (companion cells and sieve elements) specific function. Transcriptome profiling of the brl3 mutant complemented with phloem companion cell-specific BRL3-GFP at 22 °C vs. elevated temperature at 28 °C revealed enrichment of genes involved in photorespiration at high temperature, leading to better energy usage. Thus, BRL3 regulates growth responses to high temperature by controlling energy usage in response to hormonal stimuli.
Since BRL3 was found to modulate the abiotic stress–responsive genes in transcriptome analysis, the authors also observed the effects of heat and osmotic stresses on brl3 mutants. The brl3 mutant exhibited a reduced survival rate compared with the WT. However, these growth defects were restored by complementing the mutant with BRL3-GFP under the control of the phloem companion cell–specific promoter. Cumulatively, these results demonstrate the prominent role of BRL3 not only in thermomorphogenesis but also in regulating osmotic stress responses.
In conclusion, Rico-Medina et al. (2025) revealed a BRL3-mediated BR response signaling pathway specifically active in the phloem companion cells regulating cellular plasticity for temperature adaptation (Fig.). While BRI1 functions constitutively under all climate conditions, the cell type–specific expression of BRL3 not only promotes normal plant growth but also coordinates responses to high temperature and osmotic stress. However, it is intriguing to further investigate how BRI1 and BRL3 distinctly use similar downstream signaling components. BRI1 is SUMOylated (Naranjo-Arcos et al. 2023), it would be interesting to study the post-translational modifications such as SUMOylation in regulating BRL3 protein abundance. BRL3 is a promising candidate for breeding and gene editing to produce “climate smart” crops that can survive adverse weather conditions without growth penalty.
Recent related articles in Plant Physiology:
Cackett et al. (2025) reported how the cell-specific manipulation of BR signaling can regulate the chloroplast compartment in rice bundle sheath cells. Zolkiewicz and Gruszka (2025) reviewed strategies for manipulating brassinosteroid homeostasis in cereals for adapting to environmental stress.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1António C, Päpke C, Rocha M, Diab H, Limami AM, Obata T, Fernie AR, van Dongen JT. Regulation of primary metabolism in response to low oxygen availability as revealed by carbon and nitrogen isotope redistribution. Plant Physiol. 2016:170(1):43–56. 10.1104/pp.15.0026626553649 PMC 4704563 · doi ↗ · pubmed ↗
- 2Cackett L, Luginbuehl LH, Hendron RW, Plackett ARG, Stanley S, Hua L, Wang N, Kelly S, Hibberd JM. Increased chloroplast area in the rice bundle sheath through cell-specific perturbation of brassinosteroid signaling. Plant Physiol. 2025:197(4):kiaf 108. 10.1093/plphys/kiaf 10840173381 PMC 11997305 · doi ↗ · pubmed ↗
- 3Cano-Delgado A, Yin Y, Yu C, Vafeados D, Mora-Garcia S, Cheng JC, Nam KH, Li J, Chory J. BRL 1 and BRL 3 are novel brassinosteroid receptors that function in vascular differentiation in Arabidopsis. Development. 2004:131(21):5341–5351. 10.1242/dev.0140315486337 · doi ↗ · pubmed ↗
- 4Casal JJ, Balasubramanian S. Thermomorphogenesis. Annu Rev Plant Biol. 2019:70(1):321–346. 10.1146/annurev-arplant-050718-09591930786235 · doi ↗ · pubmed ↗
- 5Cohen SP, Leach JE. High temperature-induced plant disease susceptibility: more than the sum of its parts. Curr Opin Plant Biol. 2020:56:235–241. 10.1016/j.pbi.2020.02.00832321671 · doi ↗ · pubmed ↗
- 6Fàbregas N, Lozano-Elena F, Blasco-Escámez D, Tohge T, Martínez-Andújar C, Albacete A, Osorio S, Bustamante M, Riechmann JL, Nomura T, et al Overexpression of the vascular brassinosteroid receptor BRL 3 confers drought resistance without penalizing plant growth. Nat Commun. 2018:9(1):4680. 10.1038/s 41467-018-06861-330409967 PMC 6224425 · doi ↗ · pubmed ↗
- 7Naranjo-Arcos M, Srivastava M, Deligne F, Bhagat PK, Mansi M, Sadanandom A, Vert G. SUMO/de SUM Oylation of the BRI 1 brassinosteroid receptor modulates plant growth responses to temperature. Proc Natl Acad Sci U S A. 2023:120(4):e 2217255120. 10.1073/pnas.221725512036652487 PMC 9942830 · doi ↗ · pubmed ↗
- 8Rico-Medina A, Lozano-Elena F, Marqués-Bueno M, Fontanet-Manzaneque JB, Gupta A, Ana I, Caño-Delgado AI. Brassinosteroid receptor BRL 3 triggers acclimation to elevated temperature from phloem companion cells in Arabidopsis. Plant Physiol. 2025:kiaf 548. 10.1093/plphys/kiaf 54841208039 · doi ↗ · pubmed ↗