Effects of sour soup on silage fermentation performance and bacterial community of Broussonetia papyrifera
Qiming Cheng, Meiling Hou, Maoya Li, Yao Lei, Jiachuhan Wang, Yulian Chen, Yinghao Liu, Dianpeng Liu, Xiaoqing Zhang, Haoran Yu

TL;DR
This study explores how sour soup, a fermented food, can improve the fermentation quality of Broussonetia papyrifera silage by affecting bacterial communities and fermentation characteristics.
Contribution
The novel contribution is the evaluation of sour soup as a silage additive, showing its effectiveness in improving fermentation quality compared to Lactobacillus acidophilus.
Findings
Sour soup treatment improved fermentation quality by increasing crude protein and acetic acid while reducing pH and fiber.
Additives increased Lactobacillus abundance and decreased bacterial diversity.
Lower pH from additives was linked to higher protein and betaine content via structural equation modeling.
Abstract
Sour soup is a traditional fermented food, enriched with abundant organic acids, minerals and other nutrients, which contribute to human and animal health. However, due to more consumer rejection of products caused by opportunistic contamination and sulfur-containing compounds during the spontaneous fermentation of sour soups, therefore, the development and utilization of sour soup additives can be tried, such as silage improvement applications. The purpose of the current study was to evaluated the effects of sour soup as an anaerobic fermentation additive on the fermentation characteristics, microbial diversity, community composition, and alkaloids of Broussonetia papyrifera. To compare the effects of sour soup additive on Broussonetia papyrifera silage, we selected two additives, Lactobacillus acidophilus (LAB) and sour soup (S), and no additive treatment (CK). The results indicated…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
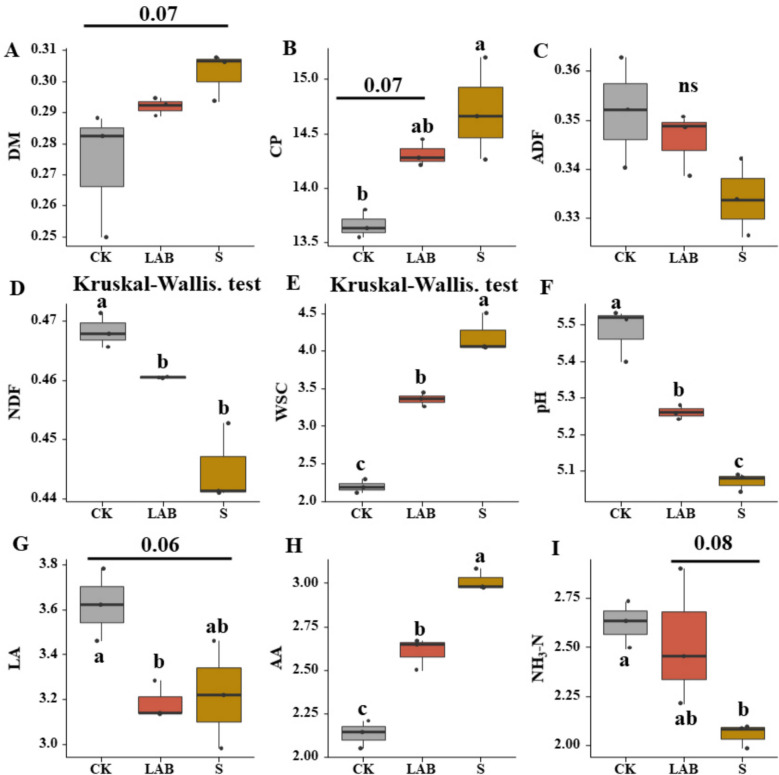 Figure 1
Figure 1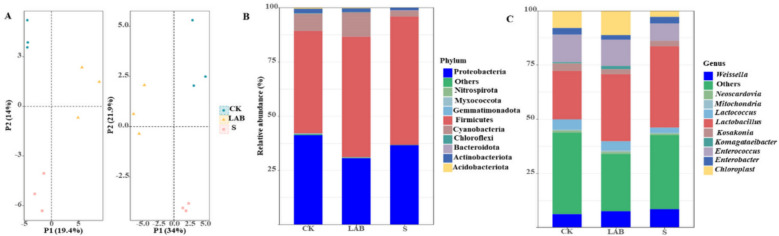 Figure 2
Figure 2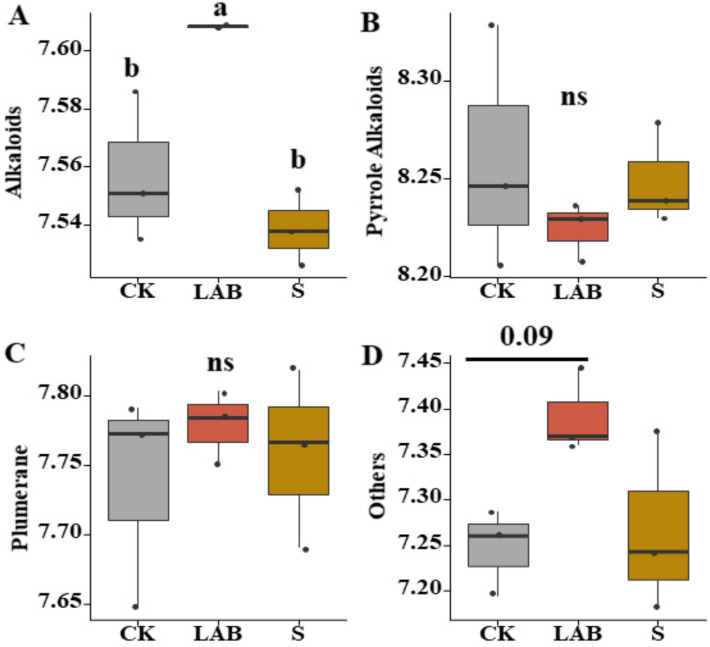 Figure 3
Figure 3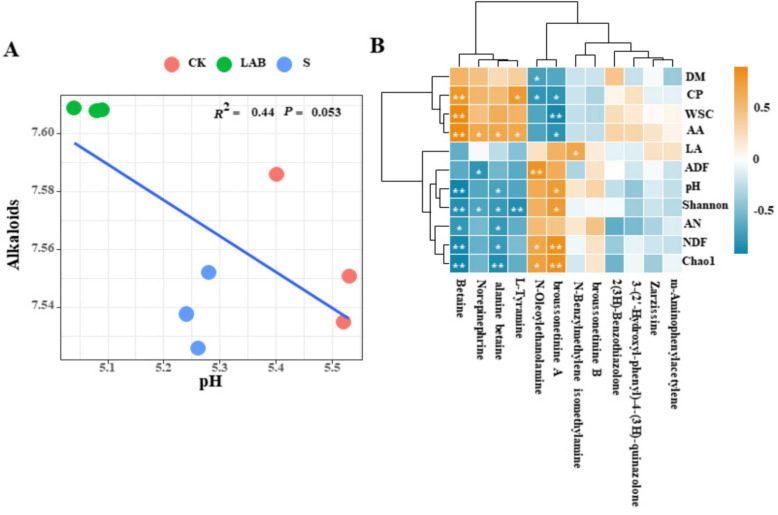 Figure 4
Figure 4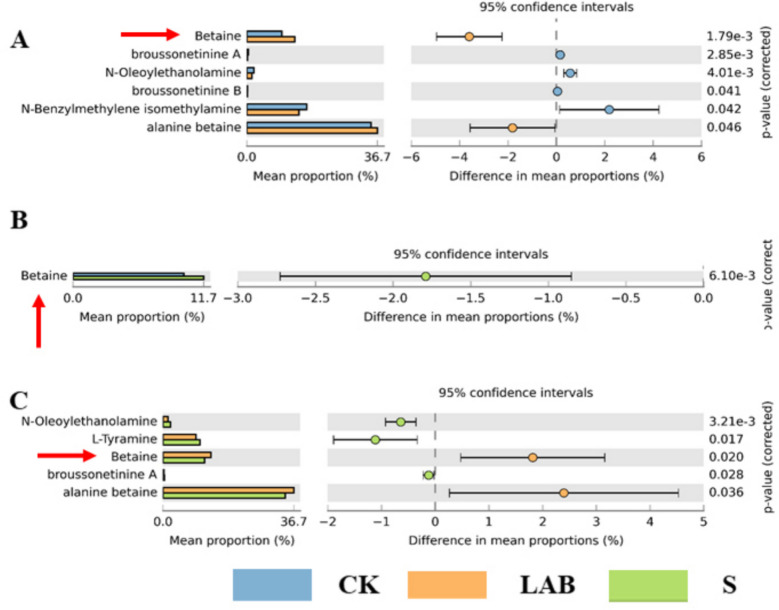 Figure 5
Figure 5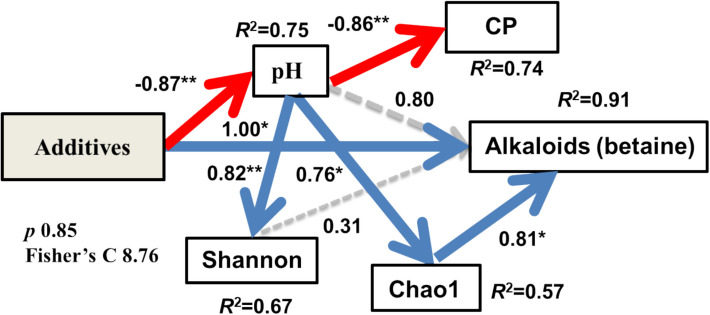 Figure 6
Figure 6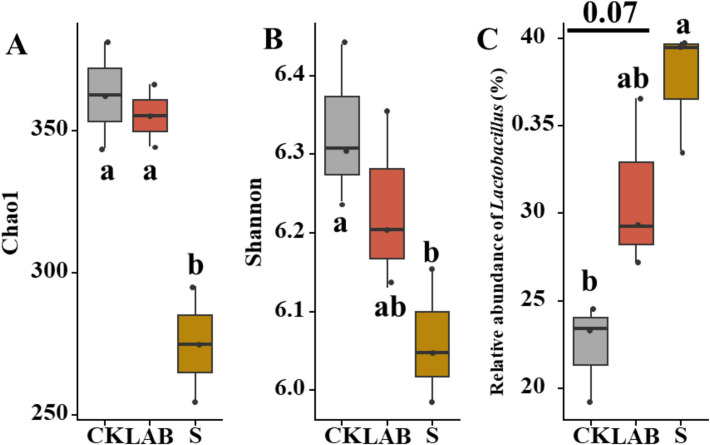 Figure 7
Figure 7- —the National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRuminant Nutrition and Digestive Physiology · Bioactive natural compounds · African Botany and Ecology Studies
Introduction
Broussonetia papyrifera L. (B. papyrifera) possesses a high protein content comparable to that of alfalfa, rendering it a promising alternative for mitigating feed shortages [1, 2]. Furthermore, paper mulberry contains a wealth of biologically active compounds, including flavonoids and terpenes with antioxidant and anti-inflammatory properties, as well as alkaloids that enhance animal intestinal digestion, thereby conferring beneficial effects on ruminant animals [3–5]. B. papyrifera grows in areas with high rainfall and humidity, making it difficult to store in the form of hay [6]. Silage exhibits good palatability, minimal nutrient loss, and is regarded as an effective method for storge fresh feed [7]. However, the high moisture content, low water-soluble carbohydrate (WSC) concentration, and high levels of acid detergent fiber (ADF) and neutral detergent fiber (NDF) result in poor fermentation quality and make silage difficult to produce, as evidenced by extensive proteolysis, unpleasant odor, and elevated butyric acid levels [8, 9]. Silage additives are frequently utilized in feed production to improve fermentation quality effectively [10]. Lactic acid bacteria inoculants possess the ability to rapidly accumulate lactic acid and reduce pH levels during the initial stages of silage production, thereby improving fermentation quality [11]. The application of these inoculants, such as L. acidophilus, promotes the dominance of Lactobacillus during ensiling, thereby accelerating and enhancing fermentation [12].
Sour soup, a regional cuisine, is rich in various organic acids, vitamins, minerals, and other nutrients [13]. It is commonly utilized as a traditional food condiment. However, opportunistic contamination and the presence of pathogenic bacteria, as well as the production of sulfur-containing compounds during fermentation [14, 15], have made people more cautious in the choice of sour soups as a type of food addition, while ruminants are more resistant as their gut characteristic relative to human, and prefer the smell [16]. The predominant microorganisms in sour soup are Lactobacillus, which hold significant potential as silage feed additives [13–15]. Research has confirmed that sour soup is beneficial to human health, studies have confirmed that sour soup contains a large number of beneficial microorganisms and has a low pH value, both of which demonstrate its potential as a silage additive. [17]. In humid climates, it can also help prevent diarrhea and other illnesses. Moreover, due to improper human use, a large amount of sour soup waste is generated, which not only pollutes the environment but also leads to unnecessary costs. For example, studies have explored whether sour soup can regulate the gut microbiota and enhance immunity [18]. Therefore, using sour soup as a novel, alternative feed additive can not only help maintain animal health and prevent animal diseases but also reduce costs and make effective use of food waste. Therefore, the development of sour soup additive silage applications can effectively utilize resources. There is currently limited research on the addition of sour soup to silage feed. At present, there is no report on the application effect of using sour soup condiment as an anaerobic fermentation additive.
B. papyrifera is rich in alkaloids [19]. Alkaloids are a type of metabolic product in plants, and some alkaloids have anti-inflammatory, antiviral and antioxidative activity effects, making them an auspicious source for feed additives [20, 21]. Study found that augmenting the content of beneficial alkaloids in silage feed indirectly promotes the enhancement of ruminant production performance [22]. However, there is currently limited research on beneficial alkaloids in woody feed and the regulatory effects of silage additives on alkaloid content [23, 24]. Additionally, certain alkaloids belong to toxic components, such as colchicine, tropane, or pyrrolizidine alkaloids (PA) [25]. Consequently, it is crucial to objectively assess changes in both beneficial and harmful alkaloids content in silage feed. Nevertheless, there is little information about the beneficial and harmful alkaloids content in paper mulberry silage.
The objectives of the present study were to examine the ensilage characteristics, bacterial community, and alkaloids content of B. papyrifera following the addition of L. acidophilus and sour soup. The study aims to: a) assess whether the addition of sour soup (S) improves the fermentation quality of B. papyrifera silage base on microbiology; b) determine whether L. acidophilus (LAB) and sour soup additives regulate the alkaloid content of B. papyrifera silage feed, and assess their impact on beneficial or harmful substances; and c) identify the pathways through which additives drive changes in alkaloids and fermentation quality of B. papyrifera silage.
Materials and methods
Raw material and silage preparation
B. papyrifera was cultivated at the experimental site in Jinbi town, Bijie city, China. The plants were harvested on June 14, 2020. The harvested plants were then chopped into approximately 2 cm lengths, homogenized, and divided into three treatments: one group with the addition of L. acidophilus (1 × 10^6^ cfu/g fresh matter), another group with the addition of sour soup (6 ml per kilogram FW of sour soup), and a control group with no additives (CK). After thorough mixing, each mixture (approximately 300 g) was packed into vacuum-sealed polyethylene plastic bags (dimensions 225 mm × 350 mm) and ensiled at ambient temperature (20 ~ 25 °C). Three bags of each mixture were opened after 45 days to assess the dynamic profiles of fermentation characteristics and bacterial community composition. Chopped ingredients such as rice soup, chili pepper, and ginger are mixed with salt and sealed for a period of time to prepare sour soup. The resulting sour soup contains various organic acids, such as lactic acid and acetic acid, as well as minerals.
Ensiling performance and nutritive values analyses
Clean containers were used to collect fresh matter (FM) and B. papyrifera silage after uniform blending for ensiling performance and nutritive value analyses. The dry matter (DM) content of the FM and silage samples was measured after drying the samples for 72 h at 65 °C in an oven. The dried samples were ground and passed through a 1 mm screen for analysis of nutritive values. The crude protein (CP) content was analyzed according to the method described by the Association of Official Analytical Chemists (AOAC, 2005). The NDF and ADF contents were determined using an ANKOM A200i Fiber Analyzer (ANKOM Technology, Macedon, NY, USA) following the method described by Van Soest [26]. The WSC content was determined using the anthrone method as described by [27]. A 20-g portion of the wheat straw silage samples was mixed with 180 mL of sterile water and stored for 24 h at 4 °C for extraction. The extracts were then filtered through four layers of cheesecloth. The pH value of the filtrate was measured using a glass-electrode pH meter (Model LEICI pH S-3C, Shanghai). The concentrations of organic acids in the filtrate, primarily lactic acid (LA) and acetic acid (AA), were measured using high-performance liquid chromatography (Model e2695, USA) methods [28]. The NH_3_-N content was analyzed using the sodium hypochlorite and phenol method described by[29].
Microbial analysis
The genomic DNA of bacterial community was extracted from the FM and wheat straw silage samples by the CTAB method. The Nano Drop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) was used to determine the concentrations and qualities of the extracted genomic DNA. The V3–V4 regions of 16S rDNA gene was targeted with the universal primer pair 341 F and 806R. The Illumina NovaSeq 6000 platform (IlluminaInc., San Diego, CA, USA) was used to sequence.The Raw pair-end reads were analyzed by the Qiime2 platform (https://qiime2.org/). Amplicon sequence variants (ASVs) were obtained by eliminating low-quality data using DADA2 (Callahan et al., 2016). Subsequently, the ASVs were taxonomically annotated against the SILVA database (https://www.arb-silva.de/, Release 138) using mothur.
Alkaloids metabolites
Metabolites extraction
Biological samples were placed in a lyophilizer for vacuum freeze-drying,and then grind to powder with a grinder. Weigh 100 mg of powder and dissolve it in 1.2 ml of 70% methanol extract, and then vortex once every 30 min, each time lasting for 30 s, a total of 6 times. Place the sample in the refrigerator at 4° C overnight, After centrifugation, the supernatant was aspirated and filtered with microporous filter membrane and stored in the injection bottle for UPLC-MS/MS analysis.
LC–MS/MS analysis
LIT and triple quadrupole (QQQ) scans were acquired on a triple quadrupole-linear ion trap mass spectrometer (QTRAP), QTRAP LC–MS/MS system, equipped with an ESI Turbo Ion-Spray interface, operating in positive and negative ion mode and controlled by Analyst 1.6.3 software (Sciex). The ESI source operation parameters were as follows: source temperature 500 °C; ion spray voltage (IS) 5500 V (positive)−4500 V (negative); ion source gas I (GSI), gas II (GSII), curtain gas (CUR)were set at 55,60, and 25.0 psi, respectively; the collision gas (CAD) was high. Instrument tuning and mass calibration were performed with 10 and 100 μmol/L polypropylene glycol solutions in QQQ and LIT modes, respectively. A specific set of MRM transitions was monitored for each period according to the metabolites eluted within this period.
Data analysis
After normalizing the original peak area information with the total peak area, the follow-up analysis was performed. Principal component analysis and Spearman correlation analysis were used to judge the repeatability of the samples within group and the quality control samples. The identified compounds are searched for classification and pathway information in KEGG, HMDB and lipid maps databases. According to the grouping information, calculate and compare the difference multiples, T test was used to calculate the difference significance p-value of each compound. The R language package ropls was used to perform OPLS-DA modeling, and 200 times permutation tests was performed to verify the reliability of the model. The VIP value of the model was calculated using multiple cross-validation. The method of combining the difference multiple, the P value and the VIP value of the OPLS-DA model was adopted to screen the differential metabolites. The screening criteria are FC > 1, P value < 0.05 and VIP > 1. The difference metabolites of KEGG pathway enrichment significance were calculated using hypergeometric distribution test.
Statistical analysis
The chemical compositions, bacterial alpha diversity (Shannon, Chao1) and alkaloid quantification (alkaloids, pyrrole alkaloids, plumerane and others, including Piperidine alkaloids, Pyridine alkaloids and Benzylphenylethylamine alkaloids) data of B. papyrifera silage were analyzed by a one-way ANOVA. Tukey’s test were utilized to assess significant differences in comparisons at the 5% level. To determine the relationship between alkaloids and CP, pH, Alpha diversity, Pearson’s correlation analysis was conducted. To compare functional profiles among groups, metabolites from different treatments were analyzed using a T-test (LAB and S, LAB and CK, S and CK). Betaine-alkaloids were enriched in silage of additions. Heat map analysis was used to confirm the relationship between the alkaloids and the fermentation quality and alpha diversity in silage. A structural equation model (SEM) was constructed to further determine the pathways by which additives drive changes in silage CP and betaine. We used piecewise SEM, and all data statistics and figures were constructed in the software of R (version 4.3.3).
Results
Chemical of raw materials
The chemical characteristics of fresh B. papyrifera are listed in Table 1. The DM contents of B. papyrifera was 34.21%. Additionally, the WSC, CP, ADF and NDF of raw material were 4.71%DM, 16.56%DM, 36.29%DM and 47.39%DM, respectively.Table 1. Chemical and microbial characteristics of substrates before ensilingItemsBroussonetia papyriferaDry matter (%)34.21 ± 1.42Water-soluble carbohydrates (% DM)4.71 ± 0.28Crude protein (% DM)16.56 ± 1.02Acid detergent fiber (% DM)36.29 ± 0.09Neutral detergent fiber (% DM)47.39 ± 0.03
Chemical composition of silage
The fermentation parameters of B. papyrifera silage are depicted in Fig. 1. Both LAB and S treatments increased CP and AA content of the silage. The AA and CP values were significantly higher in the S-treated silage compared to the CK silage (p < 0.05). Furthermore, additions of L. acidophilus and sour soup silage exhibited significantly lower pH, ADF and NDF values compared to CK silage (p < 0.05). The WSC content of L. acidophilus and sour soup treatments was significantly higher than that of CK (p < 0.05), with the highest WSC content observed in the sour soup treatments (p < 0.05). Both L. acidophilus and sour soup treatments decreased LA content, with no differences observed between CK and sour soup treatments (p > 0.05), but significantly lower LA content was observed in L. acidophilus treatments compared to CK (p < 0.05). S-treated silage significantly reduced NH_3_-N content compared to L. acidophilus silage when compared with CK (p < 0.05).Fig. 1. Fermentation quality and chemical composition in Broussonetia papyrifera silage (DM, dry matter; CP, crude protein; ADF, acid detergent fiber; NDF, neutral detergent fiber; WSC, water-soluble carbohydrates; lactic acid, LA; AA, acetic acid. CK, control treatment; LAB, Broussonetia papyrifera inoculated with Lactobacillus acidophilus treatment; S, Broussonetia papyrifera inoculated with sour soup treatment). Different letters indicate significant differences. Data following a normal distribution were analyzed using parametric ANOVA, while data not meeting the normality assumption were analyzed using the nonparametric Kruskal–Wallis (KW) test
Bacterial community of silage
As shown in Fig. 2 A, the variance of bacterial community was observed by principal component analysis (PCA) on the OTUs levels. The bacterial community in control were clearly separated from the other groups, which suggested that bacterial community changed significantly after the addition of L. acidophilus and sour soup (p < 0.05).Fig. 2. Principal coordinate analysis (PCA, A) and the relative abundance of bacterial phyla (B) and genus (C) of the Broussonetia papyrifera silage. (CK, control treatment; LAB, Broussonetia papyrifera inoculated with Lactobacillus acidophilus treatment; S, Broussonetia papyrifera inoculated with sour soup treatment)
The bacterial community at the phylum and genus levels are shown in Fig. 2B and C, respectively. At the phylum level, Firmicutes and Proteobacteria were the top two dominant phyla in all silages, accounting for more than 80% of the phylum-level composition. Firmicutes was the most dominant phylum in all treatments, comprising more than 50% of the composition in sour soup-treated silage. At the genus level, Lactobacillus was the dominant genus in all silage treatments, particularly in S-treated and LAB-treated silages, where it was significantly more abundant than other genera (p < 0.05). The proportion of Lactobacillus in CK, L. acidophilus, and sour soup was 22.42%, 31.00%, and 37.61%, respectively. The abundance of Lactobacillus in sour soup-treated silage was significantly higher than in CK. The Shannon diversity indices of CK, L. acidophilus, and sour soup were 6.32, 6.23, and 6.06, respectively. The Chao1 indices of CK, LAB, and S were 362.51, 355.34, and 274.87, respectively. The Shannon and Chao1 indices of sour soup-treated silage were significantly lower than those of CK (p < 0.05).
The alkaloids content of silage fermentation
A total of 55 alkaloids were detected in the silage feed, mainly classified into 4 categories according to secondary classification, as illustrated in Fig. 3. There were no significant differences between CK and the addition treatments in terms of pyrrole alkaloids (p > 0.05), plumerane, and others (e.g., benzylphenylethylamine alkaloids, phenolamine, piperidine alkaloids). However, L. acidophilus treatment significantly increased the levels of alkaloids (such as alanine betaine, betaine, and broussonetinine A and B) compared to CK and sour soup treatments (p < 0.05).Fig. 3. The alkaloids content of silage, mainly classified into 4 categories according to secondary classification (A, B, C, D). (CK, control treatment; LAB, Broussonetia papyrifera inoculated with Lactobacillus acidophilus treatment; S, Broussonetia papyrifera inoculated with sour soup treatment)
The relationship between chemical composition and alkaloids
All treatments of silage exhibited a negative correlation between alkaloids and pH (p = 0.05). The heatmap results showed that certain alkaloids, such as betaine, alanine betaine, and broussonetinine A, were significantly correlated with Chao1, Shannon index, pH, AA, and CP (p < 0.05), indicating a close association with the fermentation quality of silage (see Fig. 4B). However, certain alkaloids such as broussonetinine B, zarzissine, and m-aminophenylacetylene were not closely associated with the fermentation quality of silage (p > 0.05).Fig. 4. The relationship between alkaloids and pH in silage (A), correlation analysis with heatmap shown the secondary classification (alkaloids) of every alkaloids and fermentation quality and diversity, * p < 0.05,** p < 0.01 (B). (CK, control treatment; LAB, Broussonetia papyrifera inoculated with Lactobacillus acidophilus treatment; S, Broussonetia papyrifera inoculated with sour soup treatment)
pH drive betaine and CP of fermentation silage
The results of differential alkaloids analysis are shown in Fig. 5. T-test results revealed that betaine and alanine were enriched in L. acidophilus-treated silage compared to CK. Betaine was enriched in S-treated silage. Betaine content in L. acidophilus-treated silage was significantly higher than in CK silage (p < 0.05). In conclusion, betaine was enriched in additive treatments, particularly in LAB-treated silage. Furthermore, our SEM results explain 91% of the change in betaine content driven by additives. L. acidophilus and sour soup treatments had a significant and direct positive effect on betaine levels (p < 0.05). L. acidophilus-treated and S-treated silages also had an indirect effect on betaine through pH and Chao1. Specifically, additives in silage increased betaine levels by decreasing pH. Finally, additive treatments had a direct positive effect on CP. In other words, L. acidophilus and S treatments decreased pH and enhanced the CP content of silage (see Fig. 6).Fig. 5. Bar plot with significantly differential alkaloids among the Broussonetia papyrifera silage with T.test (A, CK vs LAB; B, CK vs S; C, LAB vs S). (CK, control treatment; LAB, Broussonetia papyrifera inoculated with Lactobacillus acidophilus treatment; S, Broussonetia papyrifera inoculated with sour soup treatment)Fig. 6. Structural equation model (SEM) show the direct and indirect effects of LAB and S treatments on Broussonetia papyrifera silage CP and betaine. Solid and dashed arrows, respectively, represent significant (p < 0.05) and non-significant (p > 0.05) paths. Blue and red arrows, respectively, represent positive and negative effects. Numbers adjacent to arrows represent the standardized path coefficients. R.^2^ indicates the proportion of variance explained. There was non-significant deviation of the data from the models (p = 0.85; Fisher’s C = 8.76, LAB, Broussonetia papyrifera inoculated with Lactobacillus acidophilus treatment; S, Broussonetia papyrifera inoculated with sour soup treatment)
Discussion
Previous studies have confirmed that B. papyrifera is difficult to ensile effectively due to its high moisture content, low WSC concentration, and the low content of lactic acid bacteria [30]. L. acidophilus addition are commonly used as silage additives to improve fermentation quality and ensure successful ensiling [31]. Some research has investigated the effect of L. acidophilus addition on improving the quality of paper mulberry silage [30, 32]. Our results confirm that L. acidophilus treatment mainly enhances fermentation indicators such as CP, ADF, NDF, WSC, and pH in paper mulberry feed, similar to previous studies [31], but does not have an improvement effect on LA. Even our experiment found that L. acidophilus additive reduces LA content in silage. There are two mainly reasons explain this result. Firstly, the low LA content may be due to the lower DM content in the silage [33]. Secondly, part of the LA is converted into AA during the ensiling process [34]. Our results of the AA index also confirmed that the addition of L. acidophilus significantly increases AA levels, which may also contribute to the low LA content [6], this result is consistent with the conversion of lactic acid to acetic acid. However, the data on CP, WSC, and fiber were consistent with our expectations, and the addition of L. acidophilus significantly improved the quality of B. papyrifera silage. The reasons and mechanisms for improving the quality of silage feed by adding L. acidophilus have been well-documented [31]. The number of lactic acid bacteria in natural silage materials is limited, especially after drying or storage and transportation, making acid production insufficiently rapid and resulting in a slow pH decrease [35].
Guizhou sour soup is rich in organic acids, minerals, and amino acids, making it a commonly used food additive in the local region [36]. However, there have been no reports of adding sour soup to silage feed to date. Sour soup is rich in lactic acid bacteria (i.e. L. buchneri), making it suitable for use as a microbial agent [17]. Although sour soup is a traditional food additive, its sour taste, rich amino acids can also be favored by ruminants, and more importantly, sour soup enriches the taste and nutritional value of food, which means that it also has potential uses to improve feed quality and palatability [15]. We have reported for the first time that sour soup can improve the quality of paper mulberry silage feed. Compared to L. acidophilus addition, sour soup possesses abundant organic acid characteristics, suggesting that its effect on improving silage quality may be superior to that of L. acidophilus additives [17]. Our experimental results confirm that sour soup treatment is more effective in improving quality, including DM, CP, WSC, ADF, NDF, and pH value. The addition of sufficient AA content from sour soup ensures the stability of silage feed [37]. Sour soup contains high concentrations of organic acids such as lactic acid and acetic acid, which can directly lower the pH of the silage in the early stage and inhibit the growth of spoilage bacteria. Similar to L. acidophilus addition, the addition of sour soup may convert LA into AA, resulting in high AA content in sour soup-treated silage, which may be attributed to the richness of AA in B. papyrifera raw material. Our results indicate that the effect of adding S is stronger than that of adding L. acidophilus, possibly due to the synergistic effect of lactic acid bacteria and organic acids in sour soup, which is superior to the effect of a single microbial agent [38]. This also expands the range of additive choices for the silage field, suggesting that composite synergistic additives, in addition to microbial agents, hold great potential in silage additive selection.
The Chao1 and Shannon indices of the bacterial community were evaluated based on alpha diversity (Fig. 7). As expected, sour soup treatment exhibited lower Chao1 and Shannon indices, suggesting that the sour soup treatment reduced bacterial diversity. Our PCA results indicate that the bacterial community of B. papyrifera during ensiling could be substantially influenced by sour soup and L. acidophilus treatments. Generally, Enterococcus and Lactococcus in silage are considered to be lactic acid-producing bacteria at the initial stage of fermentation [31]. It is well known that lactobacilli grow and multiply rapidly in the early stages, accumulating a large amount and lowering the pH value by producing lactic acid [39]. Therefore, lactobacilli are often used as microbial additives to improve the fermentation quality of silage [40]. Usually, Enterobacter are considered undesirable bacteria because they can compete with lactic acid, leading to a slower accumulation of non-protein nitrogen [41, 42]. In the present study, fewer Enterobacter were observed in all treatments, including CK, indicating lower attachment of Enterococcus to the B. papyrifera. Aerobic Gram-negative bacteria Kosakonia are often detected in tree construction, which is consistent with our findings [43, 44]. In summary, our results indicate that the content of undesirable bacteria in the silage is relatively low, contributing to the successful silage and high fermentation quality observed under L. acidophilus and sour soup additions. However, the diversity of sour soup-treated was not shown distinct relative to non-additions silage, which means high quality of sour soup-treated silage may caused by the enriched organic matter, such as Lactobacillus and NH_3_-N [15]. Also our bacterial community found some harmful microorganisms, such as Proteobacteria (e.g., Escherichia coli), while the relative abundance of harmful pathogenic microorganisms of additions silage was similar compared to non-addition, which confirmed that both two additions did not impose a threat of fermentation process [45].Fig. 7. Alpha diversity (A, B) and the relative abundance of lactobacillus (C). (CK, control treatment; LAB, Broussonetia papyrifera inoculated with Lactobacillus acidophilus treatment; S, Broussonetia papyrifera inoculated with sour soup treatment)
Alkaloids are a class of specialized metabolites characterized by their low molecular weight and nitrogen content, found in approximately 20% of plant species [25]. Among the approximately 12,000 structurally elucidated alkaloids, many exhibit significant biological activity, including antibacterial properties, and have been developed for therapeutic purposes [46]. However, certain alkaloids, such as colchicine and pyrrolizidine alkaloids, are toxic, and their accumulation in feed may pose a risk to animals [21, 47, 48]. We have reported, for the first time, the changes in alkaloid content in B. papyrifera silage. In our study, we observed a decrease in some harmful secondary metabolites under additive treatments, although not significant (Fig. 3B), indicating that additives not only improve the quality of silage feed but also mitigate the risk of toxin accumulation. The addition of sour soup and L. acidophilus to silage can safely feed ruminant animals. Furthermore, our results indicate that L. acidophilus treatment significantly increased the content of beneficial alkaloids (Fig. 3A). For instance, betaine and alanine betaine have antioxidant, antibacterial, and anti-inflammatory effects, as well as improving lactation performance in cows, and are often used as feed additives [49]. However, there are few reports that focus on the content of betaine in feed. The increase in beneficial alkaloids, such as betaine, in feed can promote increased milk production in ruminant animals [50]. Our results indicate that both L. acidophilus and sour soup treatments enhanced the content of betaine. Among them, the L. acidophilus treatment exhibited the highest content of betaine. In other words, the L. acidophilus treatment showed superior effects on both fermentation quality and betaine content. Many studies found that alkaloids have high medicinal and feeding value, which favors the content of beneficial alkaloids that could enhance animals’ production performance. With the advancement of silage technology, there is increasing attention to fermentation mechanisms and processes. The dynamics of quality and secondary metabolite succession during the fermentation process of silage are the focus of future research [51].
We employed SEM to analyze the pathways of betaine and protein changes in tree silage. SEM is commonly applied in natural ecology [52], including grassland and farmland ecosystems, to analyze mechanisms and processes. Our SEM results indicate that additives induce pH changes in silage, thereby reducing diversity and regulating betaine levels. Protein changes are also implicated in pH, consistent with previous research findings [53]. In other words, L. acidophilus and sour soup additions influence fermentation quality and betaine content by regulating pH levels. SEM provides robust evidence from a data-correlation perspective, elucidating the reasons for the increase in CP and betaine content.
Conclusion
Sour soup and L. acidophilus treatment can improve fermentation quality and increase the level of lactobacillus of B. papyrifera. L. acidophilus treatment increased the content of beneficial alkaloids in silage. The difference analysis of alkaloids showed that betaine was significantly enriched by the two additives, and L. acidophilus treatment had better promoting effect than sour soup treatment. Finally, the two additives drove the content of betaine in B. papyrifera silage by reducing pH. In summary, our results indicate that sour soup can be used as an additive for the application of B. papyrifera silage under anaerobic storage, it is possible to increase the study that uses edible sour soup as an alternative silage additive to apply other king of raw materials. However, its regulatory effect on bioactive substances (such as alkaloids) still needs further research.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lee EJ, Hagel J, Facchini P. Role of the phloem in the biochemistry and ecophysiology of benzylisoquinoline alkaloid metabolism. Front Plant Sci. 2013;4.10.3389/fpls.2013.00182 PMC 367809823781223 · doi ↗ · pubmed ↗
- 2Du, Seishi Y, Tetsuji O, Yimin C. Cellulase–lactic acid bacteria synergy action regulates silage fermentation of woody plant. Biotechnol Biofuel Bioprod. 2023;16.10.1186/s 13068-023-02368-2PMC 1040384237542284 · doi ↗ · pubmed ↗