Surface acoustic wave hemolysis assay for evaluating stored red blood cells
Meiou Song, Colin C. Anderson, Nakul Sridhar, Julie A. Reisz, Leyla Akh, Yu Gao, Angelo D'Alessandro, Xiaoyun Ding

TL;DR
A new method called SAW-HA uses sound waves to quickly assess the quality of stored blood, revealing how donor factors like BMI affect blood storage and transfusion outcomes.
Contribution
The SAW-HA platform introduces a rapid, on-site method for evaluating stored red blood cell quality using SAW hemolysis temperature.
Findings
SAW hemolysis temperature (SAWHT) is a reproducible marker for assessing stored RBC quality up to 42 days.
High BMI and elevated RBC triglycerides correlate with increased hemolysis susceptibility in some donors.
Low SAWHT units show disrupted redox control and tryptophan metabolism, indicating metabolic age and transfusion efficacy.
Abstract
Blood transfusion remains a cornerstone of modern medicine, saving countless lives daily. Yet the quality of transfused blood varies dramatically among donors—a critical factor often overlooked in clinical practice. Rapid, benchtop, and cost-effective methods for evaluating stored red blood cells (RBCs) at the site of transfusion are lacking, with concerns persisting about the association between metabolic signatures of stored RBC quality and transfusion outcomes. Recent studies utilizing metabolomics approaches to evaluate stored erythrocytes find that donor biology (e.g., genetics, age, lifestyle factors) underlies the heterogeneity associated with blood storage and transfusion. The appreciation of donor-intrinsic factors provides opportunities for precision transfusion medicine approaches for the evaluation of storage quality and prediction of transfusion efficacy. Here we propose a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
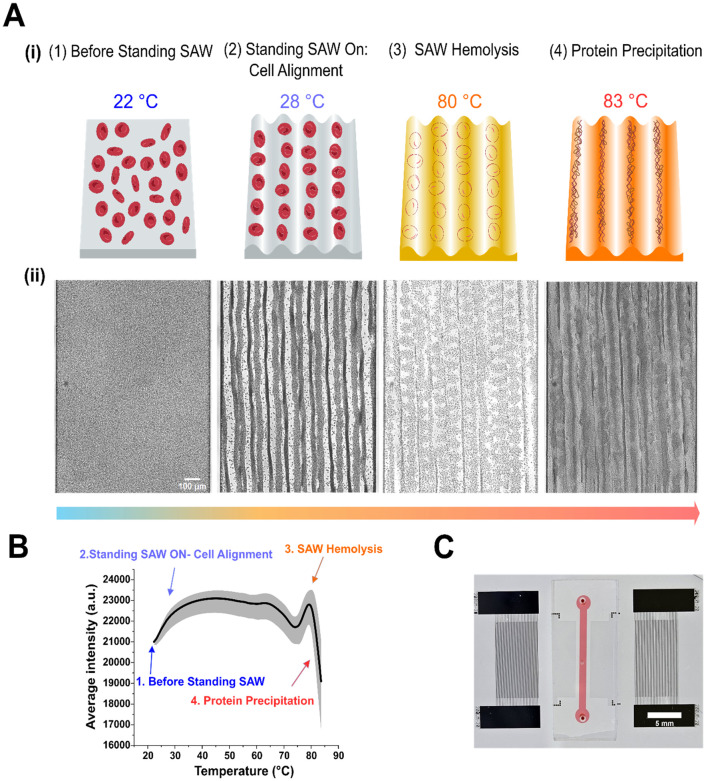 Fig. 1
Fig. 1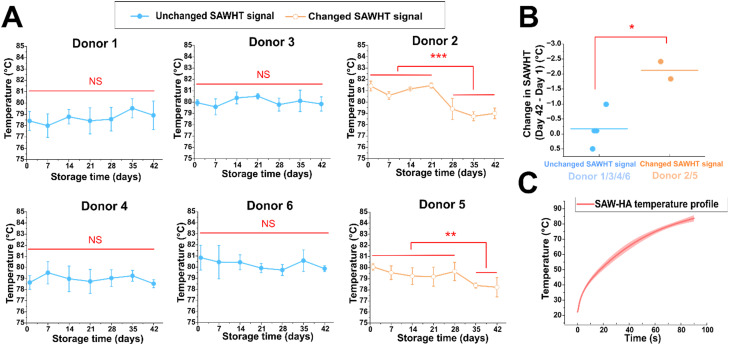 Fig. 2
Fig. 2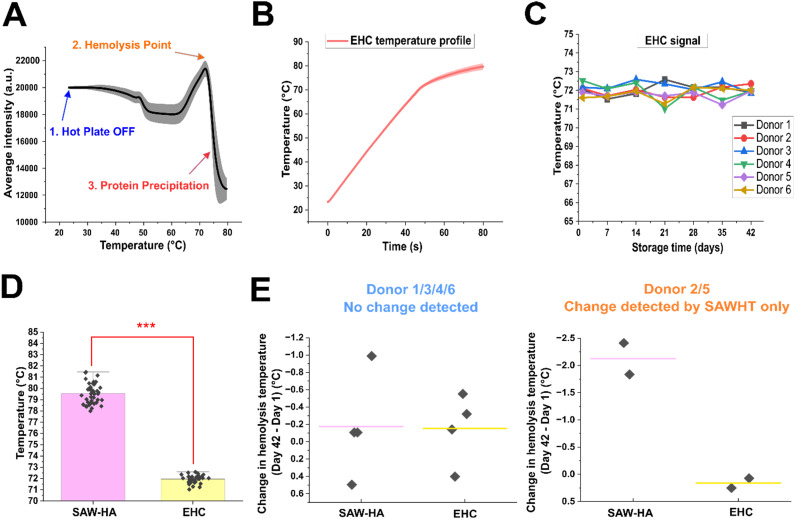 Fig. 3
Fig. 3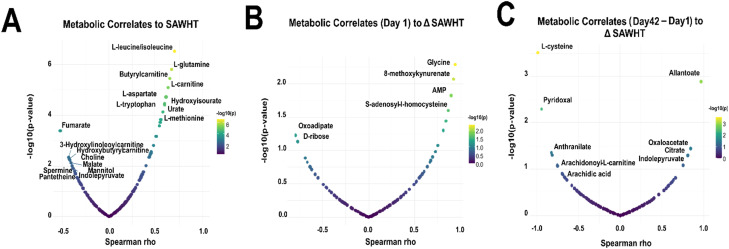 Fig. 4
Fig. 4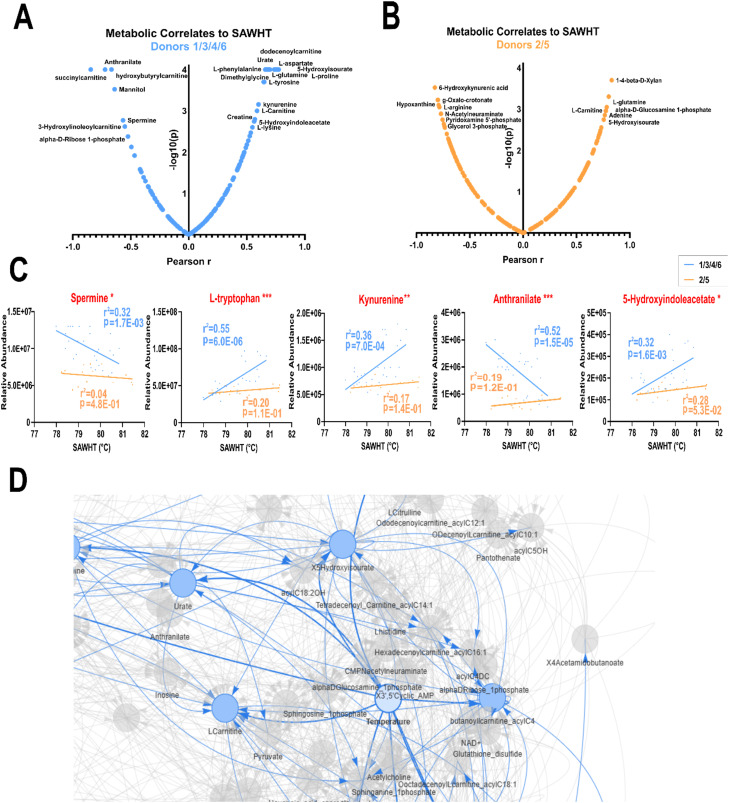 Fig. 5
Fig. 5|
|
- —National Institutes of Health10.13039/100000002
- —W. M. Keck Foundation10.13039/100000888
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBlood transfusion and management · Blood groups and transfusion · Blood donation and transfusion practices
Introduction
Blood transfusion is a life-saving medical intervention for millions of recipients worldwide. Packed red blood cells (pRBCs) – the most widely transfused blood product – are stored for up to 42 days under refrigeration in additive solutions. However, stored RBCs undergo progressive metabolic and structural changes, collectively termed storage lesions.^1^ These lesions, driven by oxidative stress and metabolic alterations, ultimately result in membrane damage, reduced deformability,^2^ and hemolysis, along with an impaired capacity to deliver oxygen,^3^ contributing to inflammatory sequelae and vascular dysfunction.^4^ Notably, RBC deterioration does not occur uniformly; donor-specific factors such as sex, age, body mass index (BMI), genetic background, and metabolism significantly influence how RBCs respond to storage.^5–9^ Current transfusion practices rely on the chronological storage age of RBCs, a metric that fails to capture metabolic age^10^—a function of RBC biophysical and biochemical integrity—which varies widely among donors. Without a benchtop assay that is easy for medical personnel to learn, use, and interpret quickly, transfusions may be non-optimized, potentially leading to poor outcomes in vulnerable patients.
Recent advances in -omics technologies, especially metabolomics, have provided new insights into RBC storage quality.^7,11–13^ These approaches have identified potential biomarkers for transfusion effectiveness by characterizing metabolic changes in stored RBCs. For instance, biomarkers of oxidative stress (glutathione (GSH) and its synthetic intermediates) correlate strongly with oxidative hemolysis within stored RBCs.^14–16^ Hypoxanthine, a marker of oxidant stress, accumulates nonlinearly through the shelf-life of stored RBCs and is linked to adverse transfusion outcomes as determined by chromium-51-labeled (^51^Cr) post-transfusion recovery (PTR) studies in autologous donors (gold standard to determine RBC storage quality per European Council and US Food and Drug Administration guidelines).^11^ Energy currencies such as ATP and NAD are also reduced during the storage period, the latter as a result of the activity of enzymes such as ADP ribose hydrolases (e.g., CD38).^17^ Accumulation of lipid peroxidation products and depletion of l-carnitine pools to repair such damage via the lands cycle are also negative predictors of post-transfusion efficacy in mouse models and humans.^13,18^
Current blood bank quality control relies on hemolysis testing of only ∼1% of monthly production via visual assessment and spectrophotometric methods.^19^ Between assessment and transfusion, units may age in the refrigerator without individual evaluation, while storage lesions progress unpredictably due to donor-specific factor. This single-parameter approach inadequately correlates with post-transfusion recovery and effectiveness.^20,21^ While the ^51^Cr-labeled PTR method is the gold standard for assessing stored blood quality and measuring transfusion outcomes, it remains prohibitively expensive due to radioactive handling requirements, specialized infrastructure, and costly disposal, limiting their clinical utility.^22^ Emerging approaches include ultra-high throughput metabolomics, which has identified predictive biomarkers but has so far remained confined to research settings due to sophistication and cost. Functional assessment methods also show promise, including RBC morphology evaluation using scanning electron microscopy (SEM)^23^ and differential interference contrast (DIC) microscopy,^24^ and more recently, high-throughput imaging flow cytometry for single-cell classification.^2,25,26^ Emerging photoacoustic microscopy^27^ and Raman spectroscopy^28^ techniques assess morphological changes of blood cells with potential for non-invasive clinical applications through blood storage bags or human skin. RBC deformability has also been recognized as a sensitive indicator of RBC functionality. Microsphiltration assays have recently been used to show that the sub-populations of RBCs that accumulate the bulk of the storage lesion are also the ones that are less deformable in vitro, and most likely to be sequestered in the spleen and erythrophagocytosed upon transfusion.^2,26^ Deformability is commonly measured using micropore filtration,^29^ micropipette aspiration,^30^ and ektacytometry (e.g., LORRCA).^24,31^ However, these approaches are inadequate for widespread clinical implementation due to high cost, specialized expertise requirements, and-in some cases-low throughput. Rapid, cost-effective, bedside technologies that correlate with metabolic profiles and predict transfusion efficacy are urgently needed to enable personalized transfusion medicine and improve clinical outcomes.^5^
To improve accessibility and throughput, lab-on-chip (LOC) technologies have emerged as promising tools for RBC quality assessment. These platforms offer portability, speed, and high-throughput capabilities. Many recent LOC techniques focus on miniaturizing traditional techniques, including imaging systems integrated with machine learning to profile the morphological heterogeneity of blood products with the goal to implement precision transfusion medicine practices.^32–35^ Microfluidic adaptations of ektacytometry,^36^ velocity-based deformability tracking,^37^ ratchet-based cell sorting,^38^ and capillary-mimicking deformability assays^39,40^ have further enabled evaluations of RBC rigidity. These approaches typically generate multidimensional datasets that require expert interpretation or complex operation workflows.
Here, we present the surface acoustic wave hemolysis assay (SAW-HA), an acoustic-integrated LOC platform that introduces a new biophysical biomarker for assessing RBC storage quality. The SAW-HA offers several practical advantages including minimal sample volume (<2 μL), rapid results (under two minutes), and straightforward quantitative output without complex data analysis. SAW technologies are known for their precise fluid control and high biocompatibility.^41–45^ Acoustic-induced heating has been used to precisely control on-chip temperatures,^46,47^ with demonstrated applications in protein-ligand interaction screening and in distinguishing healthy from sickle cell disease samples.^48^ Building on these capabilities, our assay leverages both acoustic forces and acoustic-induced heating to induce hemolysis, using the SAW hemolysis temperature (SAWHT) as a biomarker for storage-induced changes in RBC quality. Measuring SAWHT at weekly intervals during storage enables a quantitative assessment of RBC quality across a spectrum—from optimal to severely degraded units—supporting decisions on transfusion suitability. When integrated with metabolomics data, this platform provides new insights into donor-specific differences in stored RBC aging and introduces a biomarker to support the advancement of personalized transfusion medicine.
Experimental methods
Device fabrication
The SAW device was fabricated using a 500 μm thick, 76.2 mm diameter, 128° Y-cut X-propagating lithium niobate (LiNbO_3_) substrate. Two identical interdigital transducers (IDTs) were patterned on either side of a PDMS microchannel. The IDTs were fabricated using standard photolithography, starting with spin-coating a positive photoresist (S1813, Dow, USA) on the wafer. After UV exposure and development with MF319 developer (Dow, USA), layers of chrome/gold (Cr/Au, 10/100 nm) were deposited using e-beam evaporation, and excess photoresist was removed via lift-off (Remover PG, Kayaku, Japan). Each IDT comprised 30 electrode pairs with 50 μm spacing and a 10 mm aperture, yielding a frequency of ≈20 MHz for the propagating SAW.
For the polydimethylsiloxane (PDMS) channel, a negative SU8 mold was prepared by spin-coating SU8 2025 photoresist (MicroChem, USA) on a silicon wafer and patterning it with optical lithography. PDMS (Sylgard 184, Dow Corning, USA) was poured onto the mold, cured at 65 °C for 35 minutes, and punched with 0.75 mm diameter inlet/outlet holes and a 0.35 mm diameter temperature measurement hole. The PDMS microchannel (80 μm height, 1 mm width, and 10 mm long) was bonded to the LiNbO_3_ substrate using air plasma (PDC-001, Harrick Plasma, USA) and baked at 65 °C for 18 hours.
Samples and materials
Leukodepleted pRBCs in ACD-A/AS-3 were collected via apheresis at Vitalant Blood Donation Center (Denver, CO, USA) from six healthy donors (3 males, 3 females, aged 35–40) with appropriate informed consent and institutional review board (IRB) approval as specified in the Vitalant research materials agreement. All experiments were performed in accordance with the guidelines of the Declaration of Helsinki and the U.S. Federal Policy for the Protection of Human Subjects (Common Rule, 45 CFR 46), and experiments were approved by the ethics and biosafety committees at University of Colorado Boulder. Informed consents were obtained from human participants of this study. RBC units were stored at 1–6 °C and aseptically sampled weekly from day 1 to day 42. Each week, 100 μL of RBC units were frozen for future metabolic and lipidomic analysis. For weekly experiments (SAW-HA, EHC, and temperature-matched SAW-HA), RBC units were diluted 1 : 10 in PBS (v/v).
Device operation
For each set of experiments (n = 3), the PDMS channel was filled with PBS, with additional PBS droplets placed at the inlet and outlet ports. The device was then placed in a vacuum desiccator connected to a standard laboratory house vacuum line for 20 minutes for air bubble removal and temporary hydrophilicity treatment of the PDMS walls. Before each individual experiment, air bubbles were removed with ethanol, followed by manual injection of PBS into the inlet hole of the channel using a micropipette. Diluted RBC samples (∼5 μL) were injected, ensuring the channel remained bubble-free. After each experiment, the channel was cleaned with bleach followed by water washing in preparation for the next experiment. The SAW device was mounted on a custom 3D-printed holder and imaged using an inverted microscope (Eclipse Ti2, Nikon, Japan), a CMOS camera (Orca-Flash 4.0, Hamamatsu, Japan), and HCImage Live software. Videos were recorded at 2.5 fps with a 10× objective in brightfield, capturing 2048 × 2048-pixel images at 50% brightness and minimum aperture. The same channel area was imaged for consistency. Temperature inside the PDMS channel was monitored using a digital thermocouple (5TC-TT-K-36-36, Newark, USA) connected to a data acquisition (DAQ) system (cDAQ-9171 and NI TB-9212, NI, USA) and controlled via LabVIEW (NI, USA), with measurements taken for each image frame. All thermocouples were calibrated using the National Instruments data acquisition system by measuring ice-bath temperature (0 °C reference) to confirm measurement accuracy within ±0.01 °C.
An RF signal generator (33500B, Keysight, USA) and two power amplifiers (403LA, E&I, USA) were used to apply signals to the IDTs via bayonet coupling adapter (BCA) cables to printed circuit board (PCB) connectors (Fig. S1A). Three SAW devices were used throughout the experiments. For donor 1, all three devices were used at each weekly time point to establish device-to-device reproducibility (three technical replicates per time point). After confirming device consistency, one device was selected for donors 2–6 experiments. Each SAW device was individually characterized by measuring SAW resonant frequency using a network analyzer (E5061B, Keysight, USA). Resonant frequencies typically ranged from 19.56 to 19.58 MHz. For SAW-HA, the SAW was operated at about 1.2 W for 2 minutes. For temperature-matched SAW-HA, the SAW power started at 0.8 W and manually increased by 0.2 W about every 10 s to align with the heating profile of eletrical heating control (EHC). For EHC, a transparent heating plate (BT-I55D, Cell MicroControls, Norfold, VA, USA) with a microscope stage adapter (MSA-WELLP, Cell MicroControls, Norfold, VA, USA) were used (Fig. S1B). The temperature of the heating stage was controlled using a micro-temperature controller (mTC3-HT, Cell MicroControls, Norfolk, VA, USA) with the following parameters: band limit (Bnd) set to 100, final temperature set to 80 °C, with all other parameters at factory defaults.
Metabolomics/lipidomics by UHPLC/MS
To extract metabolites, either cold 5 : 3 : 2 MeOH : ACN : H_2_O (v/v/v) solution (metabolomics) or cold MeOH (lipidomics) was added in a 10 : 1 ratio to 5 μL of stored RBCs. Samples were vortexed vigorously for 30 minutes at 4 °C, then centrifuged for 10 minutes at 18 213 rcf. Using 10 μL injection volumes, the supernatants were analyzed by ultra-high-pressure-liquid chromatography coupled to mass spectrometry (UHPLC-MS – Vanquish and Orbitrap Exploris 120, Thermo). Metabolites were resolved across a 1.7 μm, 2.1 × 150 mm Kinetex C18 column using a 5 minute gradient previously described.^49^ Using 10 μL injection volumes, non-polar lipids were resolved using UHPLC coupled to ddMS^2^ using a 5 minute gradient method as previously described.^50^
Following data acquisition, .raw files were converted to .mzXML using RawConverter. Metabolites were then annotated based on intact mass, ^13^C natural isotope pattern and retention times in conjunction with the KEGG database and an in-house standard library. Peaks were integrated using El-Maven (Elucidata). Quality control was assessed as using technical replicates run at the beginning, end, and middle of each sequence as previously described. Lipidomics data were analyzed using LipidSearch 5.0 (Thermo Scientific), which provides lipid identification on the basis of accurate intact mass, isotopic pattern, and fragmentation pattern to determine lipid class and acyl chain composition.
Metabolomics and lipidomic data were analyzed using R Package (R Core Team) utilizing the following packages: shing, dplyr, plotly, ggplot2, circlize, and zip.
Image capture and analysis
All video and image processing were carried out using ImageJ.^51^ For image analysis, a single rectangular region of interest (ROI) measuring 1400 × 2000 pixels was selected to cover the entire channel area. Cell lysis and protein denaturation curves were generated by utilizing the “plot z-axis profile” function in ImageJ, which calculated the average grayscale intensity within the ROI for each frame throughout the image stack. The SAWHT of the RBCs was identified by aligning the rightmost peak of the curve with the corresponding temperature (refer to Fig. 1).
Working mechanism of SAW hemolysis assay (SAW-HA). (A) (i) Schematic representation of the four sequential phases of RBC behavior in the microfluidic chamber under the standing SAW application: dispersed RBCs (22 °C), cell alignment (28 °C), RBC lysis (80 °C), and protein aggregation (83 °C). Cell and protein illustrations were created using BioRender. (ii) Representative high-contrast images correspond to the key phases outlined in (i). The scale bar represents 100 μm. Raw, unaltered images used for analysis are included in the supplementary materials (Fig. S2A). (B) Quantitative analysis of average grayscale intensity as a function of temperature from RBC samples of six donors using SAW-HA (n = 126). The curve highlights distinct phases of cell behavior. Points corresponding to images in (A) are annotated. Note that the temperature at the third point corresponds to the biomarker SAW hemolysis temperature (SAWHT). (C) Photograph of the experimental microfluidic device used for the SAW-HA. The device features integrated SAW transducers and sample chamber with thermocouple probe hole in the middle, with a scale bar of 5 mm.
Statistical analysis
Statistical analyses were performed using Excel (Microsoft, USA), OriginPro (OriginLab, USA), and GraphPad Prism (GraphPad Software, USA) software. For comparisons between datasets, a two-sided, unpaired student's t-test was used. Significance levels were set as follows: *P < 0.05, **P < 0.01, ***P < 0.001.
Results and discussion
Device design and working principle
The schematic and working principle of the SAW-HA are shown in Fig. 1. The system consists of a PDMS microfluidic channel bonded onto a piezoelectric LiNbO_3_ substrate. Two gold IDTs are patterned symmetrically on either side of the 10 mm long, 80 μm high, and 1 mm wide channel. The device is placed in a custom-built holder fitted to a microscope stage and connected to BCA cables via a custom PCB connector (Fig. S1A). Upon activation with radio frequency (RF) signal, the IDTs generate SAWs that propagate toward each other, forming a one-dimensional standing SAW field.^52^ As the standing SAW interacts with the fluid inside the channel, acoustic energy radiates into the fluid due to the mismatch in sound velocity between the fluid and the substrate. This interaction induces pressure waves and acoustic heating effects caused by the viscous dissipation of acoustic energy into the fluid.^46^ The pressure waves create alternating pressure nodes (regions of minimum pressure) and antinodes (regions of maximum pressure). Particles suspended in the fluid are driven toward these nodes or antinodes depending on acoustic contrast factors – their density and compressibility relative to the surrounding medium. The heating profile induced by the acoustic heating effect can be precisely tuned by altering parameters such as the standing SAW frequency, power input, duty cycle, and phase.
The standing SAW was applied to induce lysis of RBCs suspended in the microfluidic channel. The lysis process was tracked using high-resolution imaging and thermocouple-enabled temperature measurements, with grayscale intensity extracted from video data (Movie S1). The intensity was plotted as a function of temperature to quantitatively represent the process (Fig. 1B). When the standing SAW was applied at 19.5 MHz and 1.2 W, RBCs aggregated at the pressure nodes due to acoustic radiation forces. This aggregation led to a noticeable increase in image grayscale intensity, which corresponded to the initial steep rise in the intensity-temperature graph. As the temperature increased from approximately 40 °C to 75 °C, the structural and mechanical properties of the RBC membranes began to change.^53^ These changes included alterations in cell density, compressibility, and acoustic contrast factors. The intensity-temperature graph showed oscillations during this phase, which could potentially be attributed to morphological transitions in the cells. The peak in the graph at approximately 80 °C represents the point of maximum image intensity, corresponding to the rupture of RBC membranes. We define this temperature as the SAW hemolysis temperature (SAWHT), which serves as a biomarker for RBC storage quality assessment. Following membrane rupture, intracellular proteins (predominantly hemoglobin) undergo denaturation, aggregation, and precipitation, which is reflected in the declining portion of the curve.
SAWHT as a biomarker evaluating RBC storage quality change
We quantified the SAWHT of each donor's RBCs as a function of storage time (Fig. 2A). Two distinct trends are apparent. Four donors (donors 1, 3, 4, and 6) maintained an unchanged acoustic signal over the 42 day storage period, with SAWHT showing minimal variation. In contrast, two donors (donors 2 and 5) exhibited significant decreases in SAWHT. For the unchanged acoustic signal group (donors 1, 3, 4, and 6), the average change in SAWHT over 42 days was −0.19 ± 0.61 °C (Fig. 2B). In contrast, the changed acoustic signal group (donors 2 and 5) displayed significant declines in SAWHT starting at specific time points. Donor 2 showed a noticeable decrease from day 28 onward, while donor 5 experienced a similar reduction starting from day 35. Both donors exhibited stabilization in SAWHT after the initial decrease, with no further significant changes observed through day 42. The average decrease in SAWHT for this group was −2.12 ± 0.41 °C, significantly greater than that of the unchanged group (Fig. 2B).
*SAW hemolysis temperature (SAWHT) serves as a biomarker for detecting donor-specific, storage-induced changes in RBC quality. (A) Weekly tracking of SAWHT for six biological samples (individual donors) throughout 42 days of cold storage. To establish device-to-device reproducibility, donor 1 was measured using all three SAW devices at each weekly time point (n = 3 technical replicates per time point). After confirming device consistency, donors 2–6 were measured using a single device. Each data point represents the mean ± s.d. of three technical replicates. (B) Comparison of SAWHT changes during storage between two donor groups. Each data point represents one donor's change in SAWHT from day 1 to day 42 (ΔSAWHT = day 42 − day 1). The horizontal bar represents the mean of each group. Unchanged acoustic signal group: donors 1, 3, 4, 6; changed acoustic signal group: donors 2, 5. (C) The average temperature profile of the SAWHT across all six donors, demonstrating consistent temperature rise over time and its reproducibility (n = 126). The shaded region represents the standard deviation. The student t-tests of independence were performed in the above figures. *P < 0.05, **P < 0.01, and **P < 0.001; two-sided, unpaired t-test. NS, not significant.
It is well established that the melting temperatures of proteins are highly sensitive to heating rates, particularly in traditional thermal shift assays.^54,55^ To ensure the reliability and comparability of results across storage periods and between donors, it is crucial to maintain a consistent heating rate during each trial. This approach minimizes variability arising from heating rate dependencies and allows for accurate assessment of stability changes in RBCs. Fig. 2C illustrates the heating profiles of RBC samples obtained from six donors over a 42 day storage period. The profiles remained consistent across different weeks for individual donors, confirming that heating conditions were precisely controlled and uniform.
Acoustic effects play an important role in detecting RBC storage quality change
To isolate the role of acoustic waves beyond thermal contributions, we first conducted an electrical heating control (EHC) experiment. This control employed electrical heating under identical experimental conditions to the acoustic method, including channel dimensions, RBC concentrations, and video parameters (Fig. S1B and S3A, & Movie S2). The purpose of this experiment was to determine whether heating alone could account for the observed variations in SAWHT and to evaluate the effects of heating alone on detecting RBC storage quality change. The intensity versus temperature profiles obtained from the EHC (Fig. 3A) revealed distinct thermal transitions compared to the SAW-HA. An intensity decrease around 49 °C likely reflects morphological changes in RBCs due to denaturation of the cytoskeleton spectrin.^56^ The rightmost peak in these profiles corresponds to the peak hemolysis point induced by electrical heating, which is followed by protein denaturation and precipitation. The average hemolysis temperature measured by the SAW-HA was significantly higher than that of the EHC (79.55 °C vs. 71.95 °C; Fig. 3D). Moreover, while donors 2 and 5 showed storage-dependent reductions in SAWHT using the SAW-HA, no such trend was observed in the EHC (Fig. 3E). This contrast highlights the importance of acoustic effects—beyond heating alone—in detecting storage-induced changes in RBC quality.
*Electrical heating control (EHC) shows that acoustic effects are critical for detecting changes of RBCs quality during cold storage. (A) Grayscale intensity versus temperature analysis of RBC samples from six donors under electrical heating reveals two key phases: (1) hemolysis point and (2) subsequent protein denaturation and precipitation (n = 126). Representative images with enhanced contrast for these phases are included in Fig. S3. Raw, unaltered images used for analysis are included in Fig. S2B. (B) Average temperature profile of EHC from all six donors, demonstrating consistent and controlled temperature progression over time (n = 126). The shaded region represents the standard deviation. (C) Weekly tracking of hemolysis temperatures across 42 days of cold storage for six biological samples (individual donors) under EHC. Each data point represents the mean of three technical replicates for each donor at each time point. Individual donor plots with mean ± s.d. and statistical significance tests are shown in Fig. S3B. (D) Comparison of the hemolysis temperatures across all donors and weeks between SAW-HA and EHC (n = 42). (E) Comparison of the change in hemolysis temperature (day 42 minus day 1) between SAW-HA and EHC for two donor groups: donors 1/3/4/6 (unchanged acoustic signal) and donors 2/5 (changed acoustic signal). Each data point represents one donor's change in SAWHT/EHC hemolysis temperature from day 1 to day 42, and the horizontal bar represents the mean value. The student t-test of independence was performed. **P < 0.001; two-sided, unpaired t-test.
To further confirm the role of acoustic waves and rule out potential confounding factors, we conducted an additional control experiment (Fig. S4). In this experiment, the heating profile of the SAW-HA was adjusted to closely replicate that of the EHC (Fig. S4A). By gradually increasing the acoustic power by 0.2 W every 10 seconds, we ensured that the temperature rise in this temperature-matched-to-EHC SAW-HA mirrored the slower heating profile observed in the EHC. The only key difference between this temperature-matched SAW-HA and the EHC is the presence of acoustic waves. The intensity versus temperature curves for the temperature-matched SAW-HA closely resembled those of the SAW-HA (Fig. S4C). Both acoustic assays exhibited higher hemolysis temperatures (∼80–82 °C) compared to the EHC (approximately 72 °C). Moreover, both assays detected storage-dependent shifts in hemolysis temperature for donors 2 and 5, a trend absent in the EHC. This finding demonstrates that the temperature difference in the initial of the heating profile or heat-shock effects (above 37 °C effect to cells) does not account for the observed donor-specific changes by the SAW-HA; rather, the presence of acoustic waves is essential for detecting these shifts.
Metabolite correlations with SAWHT can differentiate storage duration and donor phenotypes in RBCs
The relative abundance of metabolites central to energy and redox metabolism across all samples were correlated with storage duration (Fig. S5). Similar to previous reports, we observed accumulations of RBC storage biomarkers hypoxanthine, 5-oxoproline, and lactate over the storage duration,^11,12^ and glycolysis intermediates strongly negatively correlated with storage duration, in keeping with the body of literature on the metabolic changes of aging RBCs in vitro.^57^ We first correlated the change in metabolite abundance with SAWHT across all storage days (Fig. 4A and S6A). The top positive correlates were several essential amino acids (leucine, glutamine) and energy metabolism substrates (AMP). Negative correlates include dicarboxylates (citrate, fumarate) and hydroxybutyrylcarnitine (acyl-C4:OH).
A Spearman correlation analysis was performed between metabolite peak intensity (A.U.) and SAWHT. Metabolic correlates were graphed against p-value. Top correlates are labelled for (A) metabolite abundance versus SAWHT (N = 42), (B) correlates of day 1 metabolite abundance to change in SAWHT over storage duration (day 42 − day 1, N = 6), and (C) correlates of the change in metabolite abundance (day 42 − day 1) to changes in SAWHT over storage duration (day 42 − day 1, N = 6). Selected metabolite scatter plots for top correlates in A, B, and C are shown in Fig. S6.
Next, we correlated the abundance of metabolites on day 1 with the SAWHT over storage duration to identify features that predict decreased lysis temperatures on day 42 (Fig. 4B and S6B). Here, positive correlates represent metabolites that stabilize SAWHT and include glycine, AMP, S-adenosylhomocysteine (SAH), and 8-methoxykynurenate. Negative correlates include ribose and biliverdin but are not as strongly correlated.
We then correlated the change in abundance of metabolites (day 42 minus day 1) with the SAWHT over storage duration (Fig. 4C and S6C). This analysis identified cysteine as the top negative correlate, with the two donor samples that showed decreased lysis temperature accumulating cysteine. Pyridoxal, an antioxidant and the active form of vitamin B6, is also strongly negatively correlated. Anthranilate, a tryptophan metabolite like kynurenine and marker of oxidative stress, was also negatively associated, as well as long-chain fatty acids (FAs). Allantoate, a purine metabolite, showed the strongest positive correlation. In this sense, positive correlation demonstrates metabolites that are decreased in donors with decreased lysis temperature. Citrate, a carboxylic acid that abounds in citrated-anticoagulants, was also identified positively correlated along with fumarate, a catabolite of purine salvage and malate/aspartate metabolism.^58,59^
To further interrogate the metabolic profiles during storage between the SAWHT-defined two groups of donors, we performed separate correlations of metabolite and lipid levels against change in SAWHT (Fig. 5A and B and S8). These analyses revealed several key pathways that are selectively altered in donors with reduced SAWHT (Fig. 5C and S7). Donors 2 and 5 showed loss of correlation in glutamine and its downstream metabolites such as GSH, while the other donors showed positive correlations and stabilization of SAWHT glutamate and GSH. Spermine, a critical polyamine that negatively correlates with SAWHT in other donors, shows a lack of correlation in donors 2 and 5. Of major importance, tryptophan metabolism is drastically altered across multiple nodes in donors 2 and 5, with kynurenine, anthranilate, and serotonin showing reduced abundance and association with SAWHT. Furthermore, these donors show elevated triglycerides (TGs) concentration across all temperatures. Together, these results suggest that BMI may be tightly correlated with the SAWHT depression over time, as elevated TGs and disrupted tryptophan metabolism are associated with high BMI.^60–62^
*A Pearson correlation analysis was independently performed between metabolite peak intensity (A.U.) and SAWHT within the two donor groups. Top correlates are labelled for (A) donors 1, 3, 4, and 6 (N = 28) and (B) donors 2 and 5 (N = 14). (C) Select features were graphed with significance marked for features with significantly different slopes between two donor groups by simple linear regression (p < 0.05 *, p < 0.01 **, p < 0.001 **). Additional metabolites and lipids correlates of SAWHT are shown in Fig. S7. (D) A neural network analysis centered around metabolites that correlate to SAWHT (N = 42).
Current FDA guidelines permit refrigerated RBC storage up to 42 days, requiring a mean 24 hour autologous transfusion recovery rate of at least 75% across tested samples.^22^ However, statistical modeling of existing recovery data suggests only 67.3% of blood unit samples would meet this standard at 42 days, revealing significant variability in stored RBC quality that remains undetected in clinical practice due to the high cost and specialized expertise required for PTR testing.^63^ A major contributor is donor-to-donor differences, with some individuals consistently producing better-storing RBCs than others.^64^ Factors impacting the heterogeneity in the quality and post-transfusion efficacy of stored blood products include genetics,^8,65^ biology (sex, age, ethnicity),^6^ BMI,^7^ and recreational, professional, environmental, or other exposures.^66^ Such variability poses significant risks for transfusion-dependent patients—such as those with thalassemia, sickle cell anemia, or trauma—who require frequent or massive transfusions. RBC units from poor storers can undergo rapid clearance, leading to iron overload,^4^ which in turn promotes inflammatory complications such as acute respiratory distress syndrome and cardiorenal dysfunction^67,68^ and increases susceptibility to infections by siderophilic bacteria.^69^ Identifying high-quality RBC units based on donor characteristics may improve transfusion strategies, patient outcomes, and reduce healthcare costs.
Several technologies have been developed to assess RBC quality in high throughput and reveal donor–donor differences, including metabolic biomarkers,^7,13,64,70^ morphology analysis,^25,26,32,33^ and deformability-based microfluidic tests.^37,38^ While emerging methods have demonstrated success in research settings, they face distinct challenges for clinical translation. Morphology-based methods using imaging flow cytometry or automated microscopy systems require sophisticated optical setups and complex computational algorithms that generate multiple output metrics (e.g., morphological index, spherocyte percentage, echinocyte percentage), requiring expert interpretation of multidimensional data. Deformability-based microfluidic approaches have demonstrated high sensitivity but are susceptible to channel clogging (particularly with stored blood containing debris and microparticles) and involve complex operation protocols with multiple pressure adjustments and manual calibration steps.
One of the key innovations of our method is the introduction of the SAW hemolysis temperature as a biomarker for RBC storage quality. Unlike traditional assays where lysis is a preprocessing step for intracellular content analysis,^71^ our results demonstrate that the temperature at which RBCs lyse by acoustics provides valuable insights into membrane integrity and biomechanical resilience. In contrast to existing approaches, our SAW hemolysis assay offers several practical advantages: (1) simple operation with single quantitative output that does not require expert interpretation; and (2) potential for integration into cost-effective miniaturized PCB-based devices supporting simultaneous operation of multiple SAW-HA units with on-board power management for portable, point-of-care deployment. This multi-device platform could address throughput requirements for routine blood quality monitoring. Additionally, sterile sampling innovations using interconnected primary and aliquot bag systems could enable representative testing without compromising transfusable unit integrity. These features position SAW-HA as a clinically viable platform for high-throughput blood product quality assessment in blood banking applications.
After confirming here that donor RBCs stored in a manner consistent with the literature (Fig. S5), our correlation analyses identified several SAW-specific metabolites associated with major changes in SAWHT (Fig. 4A and S6A). Metabolites strongly positively correlated to SAWHT represent potential biomarkers for healthy membrane stability. This list includes many amino acids, each contributing to a multitude of vital cellular functions. For instance, glutamine is a conditionally essential amino acid and player in erythropoiesis, as well as the precursor to the vital endogenous antioxidant glutathione.^72,73^ Tryptophan, the amino acid precursor to kynurenine and serotonin, was also observed to maintain healthy membrane stability.^74^ Coincidentally, tryptophan metabolism has been shown to be dysregulated in obesity and metabolic dysfunction across multiple tissues.^75–77^ Additionally, the tryptophan metabolite kynurenine is a top metabolic correlate to osmotic stress and hemolysis despite a lack of storage-mediated changes.^12^ In the realm of fatty acid metabolism, membrane stabilizer carnitine and hydroxylated acyl-carnitines were strong negative correlates, a potential double-hit to FA equilibrium.^78^
We identified several metabolites as potential predictors of decreased lysis temperature over the storage duration (Fig. 4B and S6B) including ribose and oxoadipate. While oxoadipate is an intermediate of tryptophan metabolism, d-ribose is a protein-glycation substrate, another critical process for membrane proteins as well as a player in oxidative stress and energy metabolism.^79^ Inversely, we identified a few potential predictors of stable lysis temperature over the storage duration. AMP was decreased in the donor samples with reduced SAWHT. ATP is associated with erythrocyte membrane response to high pressure through alterations in phosphorylation and dephosphorylation of membrane proteins.^80^ Glycine was the top correlated metabolite, most likely due to its role in glutathione synthesis and regulation of oxidative signalling and damage.^81^
Finally, we highlight metabolites that correlate with changes in SAWHT (Fig. 4C and S6C). Cysteine shows the strongest negative correlation, with the two donors with decreased SAWHT accumulating cysteine over the storage duration. While cysteine has been implicated in blood fluidity in vitro, it is most likely a marker of impaired glutathione synthesis.^82,83^ Thus, it is unsurprising that features involved in redox balance are also implicated, such as pyridoxal, anthranilate, and allantoate. Pyridoxal, an aldehyde of vitamin B6, is implicated in FA synthesis and acts as an ROS scavenger.^84^ Anthranilate is another tryptophan metabolite and observed marker of donor age in stored RBCs.^85^ Purine metabolite allantoate is also a biomarker of donor age and an established marker of oxidative stress.^86^
Separation of the donor groups based on SAWHT over storage duration reveals that tryptophan and arginine metabolism over time are significantly altered over several nodes (Fig. 5A–C and S7). Additionally, glutamate, glutathione, and glutathione disulfide show downward trends. These pathways appear to be the most influential when predicting SAW hemolysis performance. A neural network analysis further identified l-carnitine, pyruvate and hydroxylated acyl-carnitines as associated players (Fig. 5D). Urate metabolism was identified as a nearby node to SAWHT, which has been shown to be associated with high BMI and increased hemolysis.^87^
Although these pathways are important in blood aging, donor statistics also play an important role in the stability of the blood sample. Donors 2 and 5 have the highest BMI among the group (Table 1). There is a significant overlap between the biomarkers associated with hemolysis and those associated with high BMI. For instance, the high correlation of SAWHT with abundance of leucine and isoleucine could indicate insulin resistance associated with high BMI.^88^ Additionally, TGs are among the lipid correlates to SAWHT (Fig. S7 and S8B). Elevated TGs are not only observed in obesity and associated with high BMI but are also a critical component of the RBC membrane.^60,61^ Upon further review of donor statistics, it was discovered that donors 2 and 5 were of Hispanic heritage (Table 1). Ethnicity has been shown to be a significant factor in RBC membrane-lipid profile, median BMI, and hemolysis in larger population studies. One such study in over 16 000 US Hispanic/Latinos found an overall prevalence of dyslipidemia in Hispanics at 65% and noted high incidences of elevated low-density lipoprotein cholesterol (LDL-C) and TGs.^89^ When evaluating ethnicity as a variable in the determination of insulin resistance via TG levels, Mexican Americans had higher prevalence of syndromes associated with insulin resistance than African Americans despite similar occurrence of obesity, hypertension, and diabetes between the two groups.^90^ Of interest to our SAW-HA, Hispanic blood donors exhibited higher markers of oxidative hemolysis than other donor populations.^91^ However, we report an altered metabolic profile for this SAW hemolysis when compared to either oxidative or osmotic hemolysis markers previously observed.^12^ Thus, with the interplay of ethnicity, BMI, lipid profile, and hemolysis; we cannot single out one factor as the main contributor to our observed decrease in SAW-hemolysis temperature. Rather, we can conclude that these factors may combine to alter membrane stability during storage, and our method can identify donor samples that may have decreased efficacy as transfusion medicine. Finally, deficient tryptophan metabolism and reduced serotonin are also associated with high BMI, potentially explaining the lack of correlation in our donor set.^62,75^
The SAW-HA provides a uniquely sensitive method for evaluating subtle changes in RBC membrane stability during cold storage. The mechanisms underlying this capability can be explained by the complex interactions between acoustic forces and cellular structures. Standing SAW imposes cyclic mechanical stresses (shear stresses, coupling with rapidly changing acoustic pressure fields and radiation pressures^47^) that actively probe membrane fragility, revealing donor-specific mechanical vulnerabilities masked under thermal conditions alone. Our data demonstrates that acoustic methods produce significantly higher hemolysis temperatures than heat-only controls (79.55 °C vs. 71.95 °C), suggesting that acoustic forces fundamentally alter the cells' response to thermal stress. This effect likely occurs through multiple pathways: 1) acoustic forces may transiently reorganize lipid packing, as molecular dynamics simulations show that ultrasound can induce oscillatory pressure changes that alter lipid tail ordering;^92,93^ 2) mechanical stimulation may modify membrane protein conformations through hydrophobic mismatch mechanisms, where small changes in bilayer thickness alters proteins toward new functional states;^94^ and 3) acoustic perturbations likely engage the spectrin-actin cytoskeletal network, potentially strengthening membrane-skeleton coupling through mechanosensitive channel activation (e.g., Piezo1) and calcium-dependent pathways.^95^
Our data highlights several metabolic correlates with the new SAWHT metric described. However, high BMI and the associated metabolic phenotype seem to be an influential factor in our limited donor group. Still, the LOC protocol identified donor samples with decreased membrane stability, thus predicted to have decreased transfusion efficacy. As the transfusion field moves toward personalized medicine, assessing donor-specific factors such as BMI may be critical for optimizing blood storage and recipient outcomes. Our SAW-integrated device provides a scalable, rapid, and accessible approach to RBC quality assessment, offering a promising on-site tool for improving transfusion practices while addressing critical concerns about donor variability. Future objectives include larger scale, follow-up studies in combination with current non-bedside assays of transfusion efficacy.
Conclusions
In summary, we have developed the surface acoustic wave hemolysis assay (SAW-HA), a microfluidic platform that introduces SAW hemolysis temperature (SAWHT) as a quantitative biomarker for evaluating stored red blood cell quality. The SAW-HA differentiated donor-specific storage behaviours across six donors over 42 days, identifying two distinct phenotypes: donors with stable SAWHT (−0.19 ± 0.61 °C) and those with declining SAWHT (−2.12 ± 0.41 °C). Integration with metabolomics revealed that donors with decreased SAWHT were characterized by disrupted redox control, deficient tryptophan metabolism, and elevated triglycerides—metabolic biomarkers indicative of decreased membrane stability—with high BMI emerging as a potential predictor of poor storage quality. Even in such a small sample group, SAWHT measurement was able to separate donor samples displaying markers of decreased membrane stability independently of donor ethnicity or metabolomics profile. This proof-of-concept platform addresses critical limitations in transfusion medicine by providing rapid (<2 minutes), sample-efficient (<2 μL), and cost-effective point-of-care analysis. It should, however, be noted that clinical translation requires validation through larger studies correlating SAWHT with gold-standard post-transfusion recovery methods, and metabolic correlations need confirmation across diverse populations to establish broader applicability. We believe the SAW-HA platform holds significant promise for advancing personalized transfusion medicine through rapid bedside quality assessment. Future applications may extend to diagnosing blood diseases, detecting pathological conditions that alter cellular membranes, and conducting fundamental membrane biophysics studies, potentially revolutionizing blood banking practices and improving patient outcomes.
Author contributions
M. S., C. C. A., A. D., and X. D., designed the study. C. C. A., J. A. R., A. D. performed omics analyses. X. D. contributed to the conceptualization. M. S., N. S., performed experiments. M. S., C. C. A. wrote the first draft of the manuscript, which was reviewed and finalized by all co-authors.
Conflicts of interest
A patent application based on this work is filed.
Supplementary Material
LC-026-D5LC00652J-s001
LC-026-D5LC00652J-s002
LC-026-D5LC00652J-s003
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zimring J. C. Established and theoretical factors to consider in assessing the red cell storage lesion Blood 20151252185219010.1182/blood-2014-11-56775025651844 PMC 4383795 · doi ↗ · pubmed ↗
- 2Peltier S. Marin M. Dzieciatkowska M. Dussiot M. Roy M. K. Bruce J. et al., Proteostasis and metabolic dysfunction characterize a subset of storage-induced senescent erythrocytes targeted for post-transfusion clearance J. Clin. Invest.20251359 e 18309910.1172/JCI 18309940067362 PMC 12043093 · doi ↗ · pubmed ↗
- 3Rabcuka J. Blonski S. Meli A. Sowemimo-Coker S. Zaremba D. Stephenson D. et al., Metabolic reprogramming under hypoxic storage preserves faster oxygen unloading from stored red blood cells Blood Adv.202265415542810.1182/bloodadvances.202200777435736672 PMC 9631703 · doi ↗ · pubmed ↗
- 4Hod E. A. Zhang N. Sokol S. A. Wojczyk B. S. Francis R. O. Ansaldi D. et al., Transfusion of red blood cells after prolonged storage produces harmful effects that are mediated by iron and inflammation Blood 20101154284429210.1182/blood-2009-10-24500120299509 PMC 2879099 · doi ↗ · pubmed ↗
- 5Isiksacan Z. D'Alessandro A. Wolf S. M. Mc Kenna D. H. Tessier S. N. Kucukal E. et al., Assessment of stored red blood cells through lab-on-a-chip technologies for precision transfusion medicine Proc. Natl. Acad. Sci. U. S. A.2023120 e 211561612010.1073/pnas.211561612037494421 PMC 10410732 · doi ↗ · pubmed ↗
- 6Kanias T. Lanteri M. C. Page G. P. Guo Y. Endres S. M. Stone M. et al., Ethnicity, sex, and age are determinants of red blood cell storage and stress hemolysis: results of the REDS-III RBC-Omics study Blood Adv.201711132114110.1182/bloodadvances.201700482029034365 PMC 5638435 · doi ↗ · pubmed ↗
- 7Hazegh K. Fang F. Bravo M. D. Tran J. Q. Muench M. O. Jackman R. P. et al., Blood donor obesity is associated with changes in red blood cell metabolism and susceptibility to hemolysis in cold storage and in response to osmotic and oxidative stress Transfusion 20216143544810.1111/trf.1616833146433 PMC 7902376 · doi ↗ · pubmed ↗
- 8Page G. P. Kanias T. Guo Y. J. Lanteri M. C. Zhang X. Mast A. E. et al., Multiple-ancestry genome-wide association study identifies 27 loci associated with measures of hemolysis following blood storage J. Clin. Invest.202113113 e 14607710.1172/JCI 14607734014839 PMC 8245173 · doi ↗ · pubmed ↗