Co-expression of a retrotransposon and re-targeted restriction factor impairs yeast growth
Sean L Beckwith

TL;DR
A study shows that when a yeast retrotransposon and a modified version of its restriction protein are both active, yeast growth is impaired.
Contribution
The novel finding is that co-expression of a retrotransposon and a re-targeted restriction factor leads to impaired yeast growth.
Findings
Co-expression of Ty1' retrotransposon and a re-targeted restriction factor variant impairs yeast growth.
The restriction factor was engineered to target a different transposon subfamily.
This effect suggests functional interactions between retrotransposons and restriction factors impact cellular processes.
Abstract
Retrotransposons and retroviruses shape genome evolution and can negatively impact genome function. Saccharomyces cerevisiae minimizes replication of Ty1 family retrotransposons using an inhibitory protein identical to the C-terminal domain of the transposon’s capsid protein. We previously elucidated the molecular basis of restriction and subfamily specific targeting and engineered a re-targeted restriction factor. Here, we report that co-expression of the Ty1' retrotransposon and a variant of its restriction factor mutated to re-target a related transposon subfamily impairs yeast growth.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
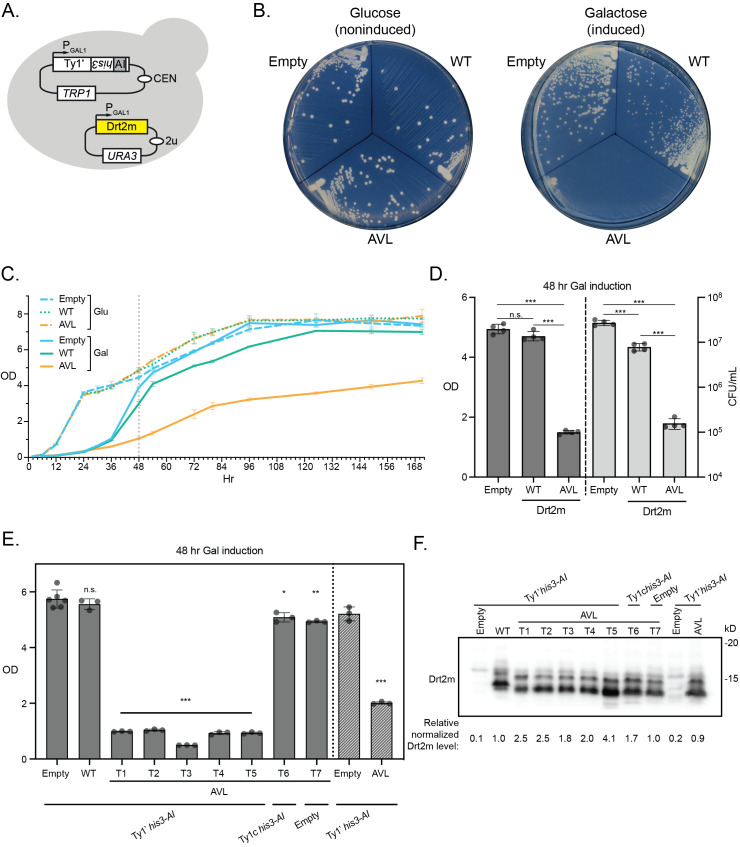 Figure 1
Figure 1|
Yeast strains used in this study. | |||||||
|
Strain |
Alias |
Label |
Genotype |
Plasmids |
|
|
Source |
|
SLBY431 |
DG4576 |
Empty |
a |
pBDG1697, pBDG1293 |
pGTy1'
|
empty pYES2 |
Beckwith et al., 2025 |
|
SLBY435 |
DG4578 |
WT |
SLBY431 |
pBDG1697, pBDG1758 |
pGTy1'
|
Drt2m |
Beckwith et al., 2025 |
|
SLBY433 |
T1 |
SLBY431 |
pBDG1697, pBDG1818 |
pGTy1'
|
Drt2m-AVL |
This study | |
|
SLBY446 |
DG4580 |
AVL (T2) |
SLBY431 |
pBDG1697, pBDG1818 |
pGTy1'
|
Drt2m-AVL |
Beckwith et al., 2025 |
|
SLBY452 |
T3 |
SLBY431 |
pBDG1697, pBDG1818 |
pGTy1'
|
Drt2m-AVL |
This study | |
|
SLBY453 |
T4 |
SLBY431 |
pBDG1697, pBDG1818 |
pGTy1'
|
Drt2m-AVL |
This study | |
|
SLBY459 |
T5 |
SLBY431 |
pBDG1818, pBDG1697 |
pGTy1'
|
Drt2m-AVL |
This study | |
|
SLBY434 |
DG4575 |
T6 |
SLBY431 |
pBDG1534, pBDG1818 |
pGTy1c
|
Drt2m-AVL |
Beckwith et al., 2025 |
|
SLBY458 |
T7 |
SLBY431 |
pBDG1818, pBDG637 |
empty pRS414 |
Drt2m-AVL |
This study | |
|
SLBY470 |
Empty |
b |
pBDG1697, pBDG1293 |
pGTy1'
|
empty pYES2 |
This study | |
|
SLBY471 |
AVL |
SLBY470 |
pBDG1697, pBDG1818 |
pGTy1'
|
Drt2m-AVL |
This study | |
|
a
| |||||||
|
b
| |||||||
|
Yeast plasmids used in this study. | |||
|
Plasmid |
Description |
Markers |
Source |
|
pBDG1293 |
pGAL-Yes2 |
|
ThermoFisher cat. no. V82520 |
|
pBDG1758 |
pBDG1293-Drt2m-6xHis |
|
Hannon-Hatfield et al., 2024 |
|
pBDG1818 |
pBDG1758-Drt2m-V266A/T270V/F312L |
|
Beckwith et al., 2025 |
|
pBDG1697 |
pGTy1'his3-AI |
|
Hannon-Hatfield et al., 2024 |
|
pBDG1534 |
pGTy1chis3-AI |
|
Saha et al., 2015 |
|
pBDG637 |
Empty vector pRS414 |
|
Brachmann et al., 1998 |
|
Antibodies used in this study. | |||
|
Antibody |
Species |
Type |
Source |
|
anti 6x-His tag |
Mouse |
Monoclonal (clone HIS.H8) |
ThermoFisher cat. no. MA1-21315 |
|
anti mouse IgG (HRP-conjugated) |
Sheep |
Polyclonal |
Cytiva cat. no. NA931 |
- —National Institute of General Medical Sciences (United States)https://ror.org/04q48ey07
- —National Institute of General Medical Sciences (United States)https://ror.org/04q48ey07
- —National Institute of General Medical Sciences (United States)https://ror.org/04q48ey07
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChromosomal and Genetic Variations · Fungal and yeast genetics research · Genome Rearrangement Algorithms
Description
Through reverse transcription, retrotransposons create a copy of themselves which is subsequently inserted into the host genome. Each transposition event, therefore, increases total copy number and can impact genome evolution. A variety of transposon control mechanisms have evolved to prevent excessive replication, including RNAi pathways, and SAMHD1, APOBEC, and MOV10 restriction factors (Goodier, 2016). The budding yeast *Saccharomyces cerevisiae * and the closely related species S. paradoxus lack these defense mechanisms, yet transposons comprise only a small fraction of the yeast genome (Garfinkel et al., 2016). S. cerevisiae contains several related groups of long-terminal repeat (LTR)-retrotransposons, the most abundant being Ty1 in many strains (Bleykasten-Grosshans et al., 2021; Curcio et al., 2015). The Ty1 genome encodes for Gag and Pol proteins. Ty1 Gag provides both capsid (CA) and nucleic acid chaperone functions. Ty1 Pol is a polyprotein that is proteolytically processed into protease, integrase, and reverse transcriptase enzymes, each of which is required for retrotransposition. Ty1 Gag assembles cytoplasmic virus-like particles (VLPs) where Ty1 protein maturation occurs and Ty1 mRNA is reverse transcribed (Curcio et al., 2015; Garfinkel et al., 1985). Ty1 Gag contains N- and C-terminal CA domains (NTD/CTD) that assemble into Ty1 VLPs (Cottee et al., 2021).
S. cerevisiae restricts uncontrolled retrotransposition of Ty1 subfamilies using distinct mechanisms: canonical Ty1 (Ty1c) is inhibited by a self-encoded restriction factor, p22/p18, whereas the closely related Ty1' is inhibited by an endogenized restriction factor, Drt2 (Cottee et al., 2021; Hannon-Hatfield et al., 2024; Saha et al., 2015). The minimal inhibitory fragment of both restriction factors (p18m and Drt2m) is a conserved CA C-terminal domain (CTD) which interacts with their respective CA via an extensive hydrophobic interface termed “Dimer-1”. We recently reported that p18m and Drt2m are highly specific to their subfamilies (Beckwith et al., 2025). There are only three amino acid residues found within Dimer-1 that diverge between the subfamilies but are highly conserved within each subfamily: A266, V270, and L312 (AVL) in Ty1c and p18 compared to V266, T270, and F312 (VTF) in Ty1' and Drt2. By mutating these three residues, we re-targeted each restriction factor to the opposite transposon.
Here, we demonstrate that the re-targeted restriction factor, Drt2m-AVL, causes a growth defect specifically when co-expressed with the Ty1' retrotransposon. We expressed the Ty1' retrotransposon, marked with the his3-AI retrotranscript indicator gene (which enables monitoring of mobility frequency), on a plasmid under the GAL1 galactose-inducible promoter (Curcio & Garfinkel, 1991) ( ** Figure 1A ** ). Wildtype Drt2m or the re-targeted mutant Drt2m-AVL were tagged with a C-terminal hexahistidine tag and expressed alongside Ty1' his3-AI on a separate plasmid, also under the control of the GAL1 promoter. Experiments were performed in a S. paradoxus yeast strain lacking any full-length Ty elements (Chen et al., 2022; Cottee et al., 2021; Saha et al., 2015).
We observed that co-expression of Drt2m-AVL and Ty1' drastically impaired cell growth compared to wildtype Drt2m or empty vector. We measured this growth defect in both plate and liquid assays and across different time points. On agar plates, cell growth is not impaired when the strains are grown on glucose media which represses gene expression from our constructs controlled by the GAL1 promoter ( ** Figure 1B ** ). When expression is induced by growing cells on galactose plates, wildtype Drt2m expression causes a slight growth reduction as seen by colony size. However, Drt2m-AVL expression results in extremely reduced colony size. We further measured liquid growth of these three strains in non-inducing (glucose) and inducing (galactose) media over more than 1 week (171 hours). Yeast cells grow more quickly in glucose, their optimal carbon source, than in galactose. In glucose, growth of the three strains is not dramatically different ( ** Figure 1C ** , see Extended Data for t -test comparisons). However, when Drt2m-AVL is expressed on galactose, growth of the culture is dramatically slowed. Expression of wildtype Drt2m slightly reduces culture growth but the culture still saturates and growth plateaus by the end of the data collection period. The strain expressing Drt2m-AVL never approached a cell density close to empty vector or non-induced cultures. We selected the 48 hr time point to further examine growth in liquid media as Drt2m-AVL displayed a marked growth defect. We validated optical density (OD) as an accurate, rapid measure of cell density of the culture after 48 hr in galactose media. Drt2m-AVL expressing cultures had significantly lower OD than wildtype Drt2m and empty vector strains and had one to two orders of magnitude fewer colony forming units (CFU) per mL of culture ( ** Figure 1D ** ).
We next tested the growth defect of Drt2m-AVL co-expressed with Ty1' in several different strain constructions by measuring OD after 48 hr of growth in galactose media. The growth defect was observed in strains generated through four independent transformations (T1-T4) and a strain generated by transforming in the Drt2m-AVL and Ty1' his3-AI plasmids in the opposite order (T5). Co-expression of Drt2m-AVL with Ty1c his3-AI (T6) or with an empty vector (T7) did not dramatically impair growth ( ** Figure 1E ** ). Furthermore, the growth defect was reproducible in a distinct strain background of Ty-less S. paradoxus ( ** Figure 1E ** , hatched bars on right). Finally, we assessed Drt2m and Drt2m-AVL protein expression by western blot. All strains expressed protein well with some variability in protein level (see Extended Data for quantification of total protein levels). Protein expression does not appear to explain the striking growth defect especially when Drt2m-AVL and Ty1' are co-expressed.
In summary, we have shown a marked growth defect in yeast co-expressing Drt2m-AVL and Ty1' both on agar plates and liquid media. This defect is not an artifact of plasmid construction as it is only observed in galactose media that induces gene expression. Wildtype Drt2m expression causes a slight reduction in growth seen qualitatively by colony size on agar plates but inconsistently in liquid media measured by OD 660 at 48 hr. The dramatic growth impairment caused by Drt2m-AVL expression is consistently observed on solid and liquid media and is reproducible in several strain constructions and in two S. paradoxus backgrounds. Differences in protein expression do not explain the impaired cell growth. The severe growth defect is only observed when Drt2m-AVL and Ty1' are both expressed, not when Ty1' is expressed with wildtype Drt2m nor when Drt2m-AVL is expressed with Ty1c or in the absence of any transposon. The mechanism behind this growth defect remains to be determined but may involve cellular products that modulate the process of Ty1 retrotransposition (Curcio et al., 2015).
Methods
Yeast strains, plasmids, and media
Yeast strains and plasmids used in this study are included under Reagents . All yeast strains are Ty-less Saccharomyces paradoxus . SLBY470 and SLBY471 were generated by transforming the indicated plasmids into strain DG2204 (Beckwith et al., 2023). Yeast was grown in synthetic complete (SC) media lacking uracil (-U) and tryptophan (-T) and containing the indicated sugar at 2% w/v. For induction in liquid media, starter cultures were grown overnight at 30 °C in synthetic media containing raffinose, diluted 1:20 into media containing either galactose or glucose, and grown at 22 °C. Optical density (OD) readings were measured at 660 nm; readings greater than 1.0 were diluted 1:10 in water and the diluted value was multiplied by 10. Colony forming units (CFU) per mL of culture were calculated by diluting culture in sterile water, plating on SC-U-T glucose plates, and counting colonies after four days of growth at 30 °C (see Extended Data for dilution factors and volumes). All graphs show data from at least three replicates and colony growth on plates are representative images of three replicates.
Protein extraction and western blotting
Western blotting was performed on total protein extracted from galactose-induced yeast by trichloroacetic acid (TCA) precipitation. Cells were broken by vortexing in the presence of glass beads in 20% TCA for 5 min, cooling on ice for 3 min, and vortexing again for 5 min. Glass beads were washed in 5% TCA; the liquid was transferred to a new tube and centrifuged at 9,391 x g for 10 min. The pellet was resuspended in 150 μL of 4x Laemmli Sample Buffer (Bio-Rad cat. no. 1610747) containing 2-mercaptoethanol and boiled for 5 min. Proteins were separated on AnyKD Stain-Free Protein Gels and visualized according to supplier specifications (Bio-Rad cat. no. 4568126). PVDF membranes were immunoblotted with monoclonal mouse hexa-histidine antibody clone HIS.H8 (ThermoFisher cat. no. MA1-21315) diluted 1:3000 in 2.5% milk-TBST and then peroxidase-conjugated anti-mouse IgG (Cytiva cat. no. NA931) diluted 1:5,000 in TBST. Immune complexes were detected with WesternBright enhanced chemiluminescence (ECL) detection reagent (Advansta cat. no. K-12049-D50). Imaging was performed using a ChemiDoc MP (Bio-Rad). Total protein normalization and band quantification was done using Image Lab software (Bio-Rad). Precision Plus Kaleidoscope protein standards (Bio-Rad cat. no. 1610395) were used to estimate molecular weights. The western blot shown is representative of three replicates.
Reagents
**: **
**: **
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Beckwith Sean L. Cottee Matthew A. Hannon-Hatfield J. Adam Newman Abigail C. Walker Emma C. Romero Justin R. Stoye Jonathan P. Taylor Ian A. Garfinkel David J. 2025109 Probing the molecular determinants of Ty 1 retrotransposon restriction specificity in yeast PLOS Genetics 21101553-7404 e 1011898 e 101189810.1371/journal.pgen.101189841066520 PMC 12530519 · doi ↗ · pubmed ↗
- 2Beckwith Sean L. Nomberg Emily J. Newman Abigail C. Taylor Jeannette V. Guerrero-Ferreira Ricardo C. Garfinkel David J. 2023717 An interchangeable prion-like domain is required for Ty 1 retrotransposition Proceedings of the National Academy of Sciences 120300027-842410.1073/pnas.2303358120 PMC 1037261337459521 · doi ↗ · pubmed ↗
- 3Bleykasten-Grosshans Claudine Fabrizio Romeo Friedrich Anne Schacherer Joseph 2021611 Species-Wide Transposable Element Repertoires Retrace the Evolutionary History of the Saccharomyces cerevisiae Host Molecular Biology and Evolution 38101537-17194334434510.1093/molbev/msab 17134115140 PMC 8476168 · doi ↗ · pubmed ↗
- 4Baker Brachmann Carrie Davies Adrian Cost Gregory J. Caputo Emerita Li Joachim Hieter Philip Boeke Jef D. 1998130 Designer deletion strains derived from Saccharomyces cerevisiae S 288C: A useful set of strains and plasmids for PCR-mediated gene disruption and other applications Yeast 1420749-503X 11513210.1002/(sici)1097-0061(19980130)14:2<115::aid-yea 204>3.0.co;2-29483801 · doi ↗ · pubmed ↗
- 5Chen Jingxuan Mc Queary Holly Hall David W. Philippsen Peter Garfinkel David J. Bergman Casey M. 2022120 Genome Assembly of the Ty 1-Less Saccharomyces paradoxus Strain DG 1768 Microbiology Resource Announcements 1112576-098X 10.1128/mra.00868-21PMC 877260235049349 · doi ↗ · pubmed ↗
- 6Cottee Matthew A. Beckwith Sean L. Letham Suzanne C. Kim Sarah J. Young George R. Stoye Jonathan P. Garfinkel David J. Taylor Ian A. 2021922 Structure of a Ty 1 restriction factor reveals the molecular basis of transposition copy number control Nature Communications 1212041-172310.1038/s 41467-021-25849-0PMC 845837734552077 · doi ↗ · pubmed ↗
- 7Curcio M J Garfinkel D J 199121 Single-step selection for Ty 1 element retrotransposition.Proceedings of the National Academy of Sciences 8830027-842493694010.1073/pnas.88.3.936PMC 509291846969 · doi ↗ · pubmed ↗
- 8Curcio M. Joan Lutz Sheila Lesage Pascale 201542 The Ty 1 LTR-Retrotransposon of Budding Yeast, Saccharomyces cerevisiae Microbiology Spectrum 322165-049710.1128/microbiolspec.mdna 3-0053-2014 PMC 439924225893143 · doi ↗ · pubmed ↗