A Japan-focused review of East Asian Acicnemis Fairmaire, 1849 (Coleoptera, Curculionidae) reveals cryptic diversity in a well-studied region

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
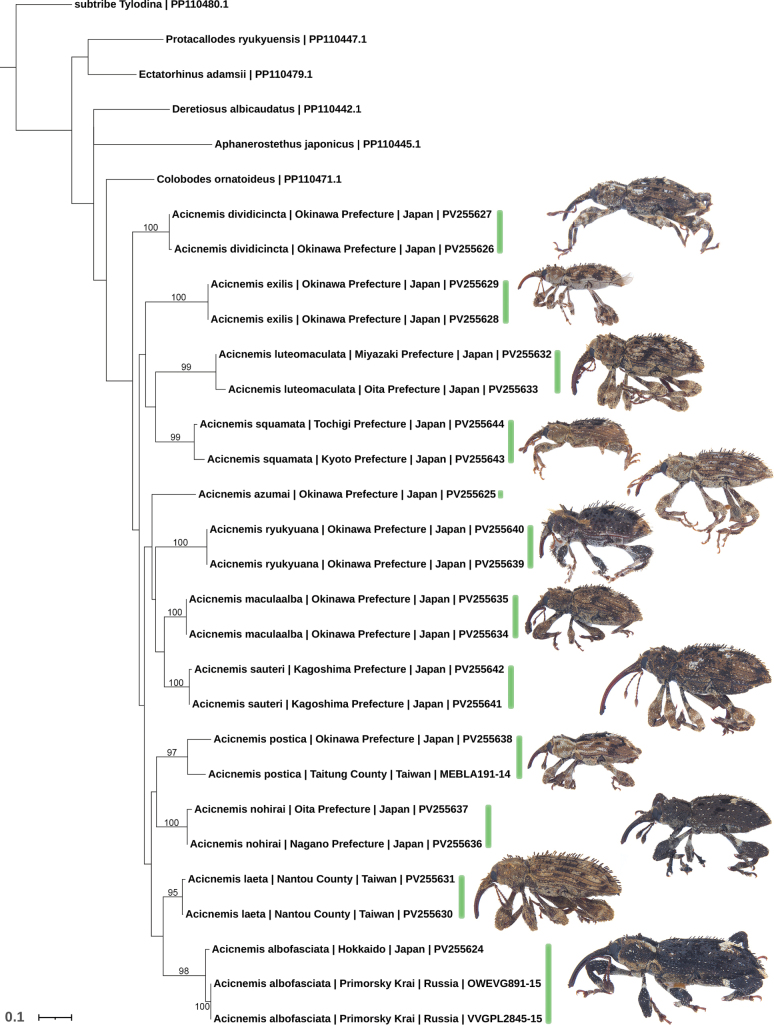 Figure 1
Figure 1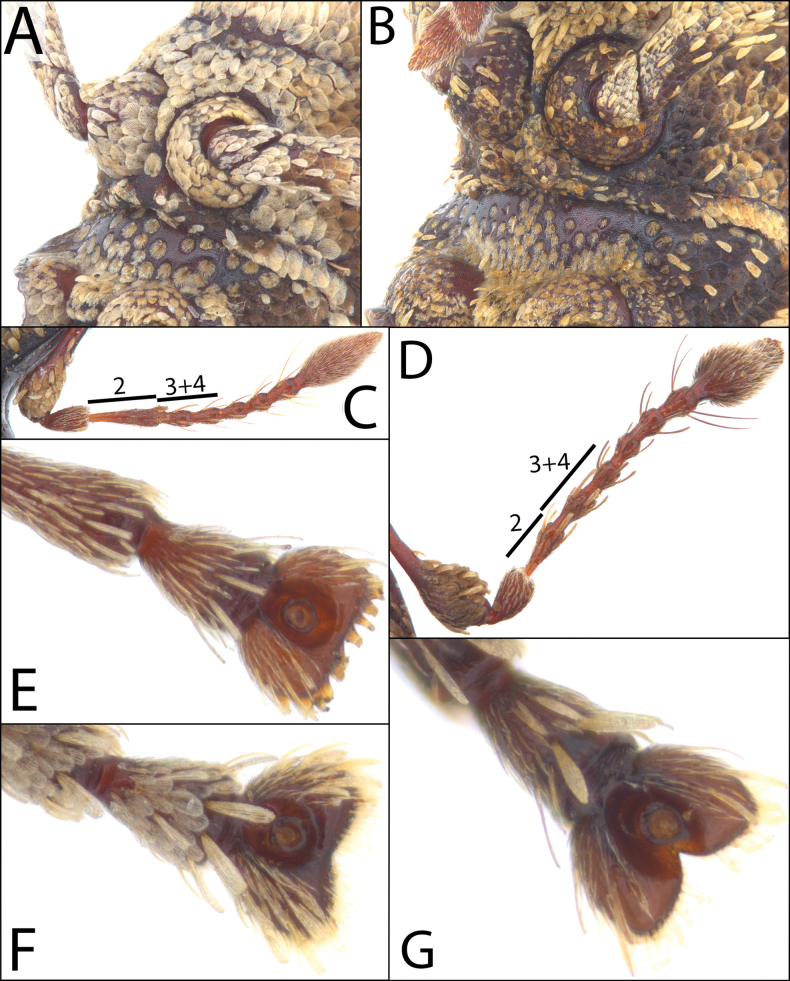 Figure 2
Figure 2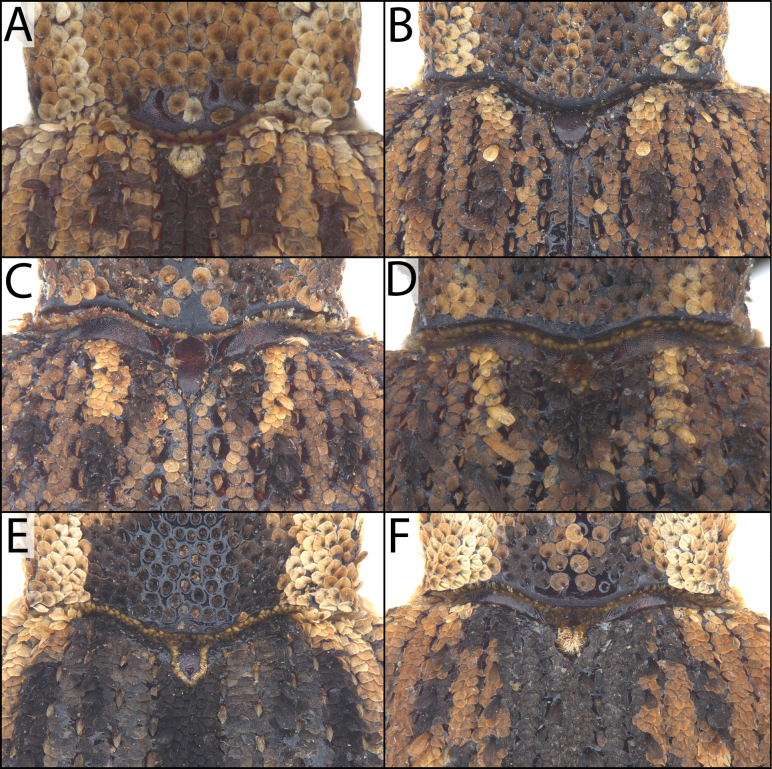 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5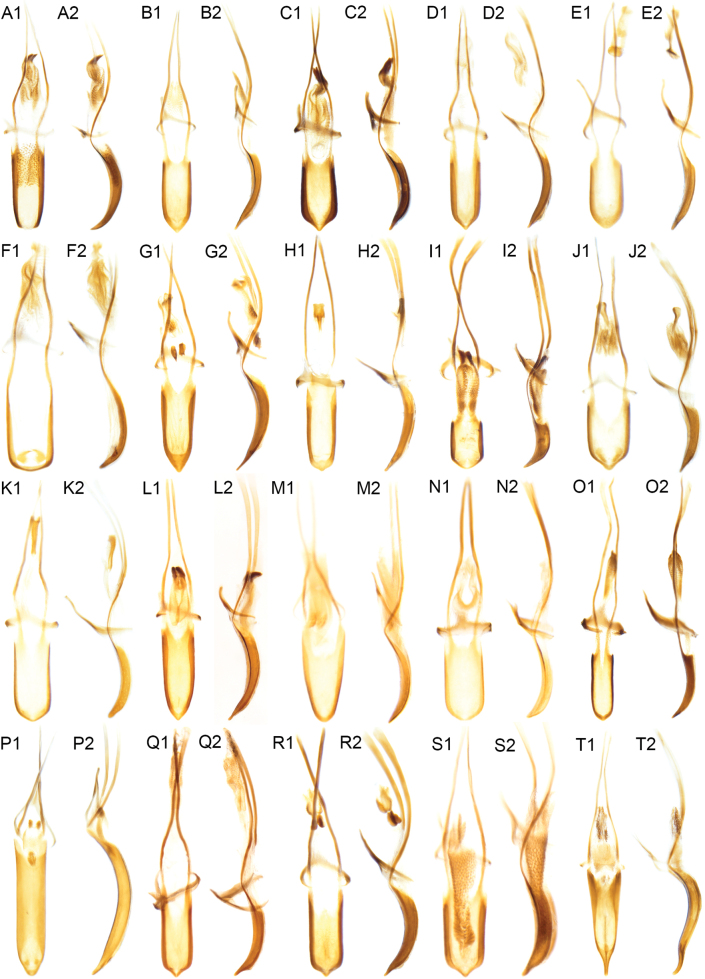 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12 Figure 13
Figure 13 Figure 14
Figure 14 Figure 15
Figure 15 Figure 16
Figure 16 Figure 17
Figure 17 Figure 18
Figure 18 Figure 19
Figure 19 Figure 20
Figure 20 Figure 21
Figure 21 Figure 22
Figure 22 Figure 23
Figure 23 Figure 24
Figure 24 Figure 25
Figure 25 Figure 26
Figure 26 Figure 27
Figure 27 Figure 28
Figure 28 Figure 29
Figure 29 Figure 30
Figure 30 Figure 31
Figure 31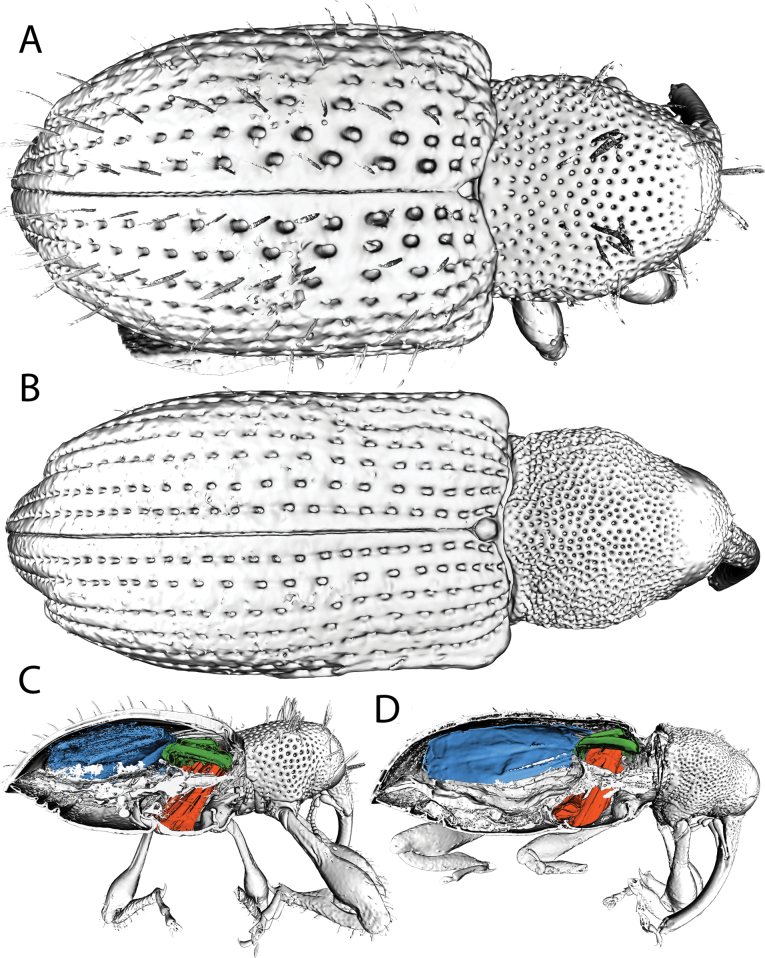 Figure 32
Figure 32| 1 | Third tarsomeres truncate or weakly emarginate (Fig. |
|
| – | Third tarsomeres strongly emarginate (Fig. |
|
| 2 | Second and fourth elytral intervals with erect scales |
|
| – | Second and fourth elytral intervals without erect scales (may have small erect scales at elytral apex) |
|
| 3 | Erect scales on odd elytral intervals elongate, hair-like (distinctly longer than antennal club) |
|
| – | Erect scales on odd elytral intervals stubby, round (shorter than antennal club) |
|
| 4 | Sclerolepidia inconspicuous, low and rounded; third tarsomeres truncate; scales covering body predominantly brown |
|
| – | Sclerolepidia distinct and protruding; third tarsomeres weakly emarginate; scales covering body predominantly gray |
|
| 5 | Second elytral interval with erect scales |
|
| – | Second elytral interval lacking erect scales (may have small erect scales at elytral apex) |
|
| 6 | Scutellum distinctly large, wide, and lacking scales (Fig. |
|
| – | Scutellum slender and largely covered in scales dorsally (e.g., Fig. |
|
| 7 | Internal sac of aedeagus protruding but with roughened musculature proximal to pedon (Fig. |
|
| – | Internal sac of aedeagus protruding but with roughened musculature located at distance from pedon (Fig. |
|
| 8 | Even-numbered elytral intervals costate |
|
| – | Elytral intervals not costate |
|
| 9 | Posterior edge of prosternum with prominent projections contiguous with the fore-coxae (Fig. |
|
| – | Posterior edge of prosternum lacking prominent projections contiguous with the fore-coxae (e.g., Fig. |
|
| 10 | Cuticle distinctly naked (lacking scales) across much of body; pronotum roughly sculptured and carinate |
|
| – | Cuticle largely covered in scales (sometimes specimens are denuded); pronotum punctate but not roughly sculptured or carinate (except at base of pronotum in |
|
| 11 | Pronotum with two exposed, bare, red tubercles medially at the base (Fig. |
|
| – | Pronotum lacking tubercles at base (Fig. |
|
| 12 | Scutellum distinctly shiny and bare dorsally (Fig. |
|
| – | Not with the above combination of characters |
|
| 13 | Pedon distinctly tapered apically into a fine point (Fig. |
|
| – | Not with the above combination of characters |
|
| 14 | Small species (3.0–3.8 mm) with brown scaling and distinct white scaled V-shaped band in posterior 1/2 of elytra; pedon tube-shaped and internal sac with a small, sclerotized structure basally, and 2 prong-like structures anteriorly at its apex (Fig. |
|
| – | Size and color variable ; aedeagus not as above |
|
| 15 | Small species (3.0–4.4 mm) with pale and dark brown scales, and shallow, white-scaled, V-shaped band in posterior 1/2 of elytra; internal sac of aedeagus distinctly scaly in appearance and protruding; apex of internal sac ending in two scaly feather-like tips; apex of pedon with distinct sinuation and pointed (Fig. |
|
| – | Size and color variable; aedeagus not as above |
|
| 16 | Large species (5.1–7.0 mm) with pale brown, dark gray, and black scales, and a tan-colored, V-shaped, transverse band across middle of elytra; scutellum covered in erect scales (Fig. |
|
| – | Size and color variable; aedeagus not as above |
|
| 17 | Exceptionally small species (2.1–3.0 mm), covered in pale brown, white, and gray scales, with 2 white-scaled spots in the apical 1/2 of the elytra; pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a blunt, rounded point; internal sac without tuberculate or roughened musculature inside the aedeagus, but with a non-hooked protruding structure (unmodified in posterior 2/3, sclerotized in anterior 1/3) that reaches the tip of the temo (Fig. |
|
| – | Larger species (3.2–7.0 mm), variable in color; aedeagus not as above |
|
| 18 | Elytra with distinct white-scaled, X-shaped band across elytra (extremely bright in some specimens); pedon with lateral edges approx. parallel in dorsal view, but sinuating in distal 1/2 into a distinct point; internal sac protruding and clear across basal 2/3; with two bean-shaped and U-shaped sclerotized structure anteriorly at its apex (Fig. |
|
| – | Elytra with white-scaled V-shaped or transverse band, but never an X-shaped band; aedeagus not as above |
|
| 19 | Brightly colored species, with dull orange, white, and brown scales; pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a blunt, rounded point; internal sac not visible inside aedeagus, but protruding basally and with 2 sclerotized cylinder-shaped structures posteriorly at the base (Fig. |
|
| – | Dark scaled species, with black and gray scales covering most of body; aedeagus not as above |
|
| 20 | Internal sac with tuberculate (roughened) musculature, visible inside of aedeagus; internal sac protruding from aedeagus and weakly hooked anteriorly (Fig. |
|
| – | internal sac not with tuberculate musculature visible inside of aedeagus (only in protruding portion of sac in some species); body size variable |
|
| 21 | internal sac with hook-shaped protruding structure and roughened musculature at the base (Fig. |
|
| – | Internal sac with modified protruding structure but not with roughened musculature and weakly hooked shape; pronotum never with impunctate longitudinal midline; size variable |
|
| 22 | Internal sac without club-shaped structure anteriorly at its apex (Fig. |
|
| – | Internal sac with club-shaped structure anteriorly at its apex; pronotum nearly uniform in color, with black and dark gray scales (no prominent lateral bands) |
|
| 23 | Club of internal sac surrounded closely by roughened musculature (Fig. |
|
| – | Club of internal sac not surrounded closely by roughened musculature (Fig. |
|
- —Canadian Museum of Nature 100014378 http://doi.org/10.13039/100014378
- —Natural Sciences and Engineering Research Council of Canada 501100000038 https://ror.org/01h531d29 http://doi.org/10.13039/501100000038
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsColeoptera Taxonomy and Distribution · Coleoptera: Cerambycidae studies · Forest Insect Ecology and Management
Introduction
Weevils (Coleoptera: Curculionoidea) are one of the most diverse (ca 62,000 species) and economically important animal groups, but remain poorly studied throughout much of Asia (Oberprieler et al. 2014; Sprick and Floren 2018). The diverse genus Acicnemis Fairmaire, 1849 (Curculionidae: Molytinae) contains more than 220 described species distributed across the Eastern Palearctic, Oriental, and Australasian Regions (Hubenthal 1919a, b; Zimmerman 1967; Kojima and Morimoto 2004; Pullen et al. 2014). Members of Acicnemis are usually cryptically colored and often bear erect scales dorsally that enhance their disruptive patterning. The genus Acicnemis is placed in the tribe Trachodini with four other genera, Karekizo Morimoto, 1962 (2 spp.), Ochodontus Desbrochers des Loges, 1897 (1 sp.), Semelima Pascoe, 1872 (1 sp.), and Trachodes Germar, 1824 (ca 30 spp.), which are also all Old World genera (Morimoto and Miyakawa 1995; Alonso-Zarazaga and Lyal 1999; Alonso-Zarazaga et al. 2023). The core genera of Trachodini, Acicnemis and Trachodes, have a complex taxonomic history (see Hubenthal 1919a; Zimmerman 1967), and have typically been distinguished based on genital (in part) and hindwing morphology (fully developed hindwings in Acicnemis, reduced in Trachodes) (Morimoto and Miyakawa 1995). Hubenthal (1919a, b) compiled the most comprehensive key to Acicnemis and is responsible for describing many species; however, those works lack illustrations and are outdated as many new synonyms and species have since been discovered. Subsequent works have largely been disjunct, single species descriptions and rarely include keys, illustrations, or useful comparative notes (e.g., Kleine 1924; Voss 1941; Zimmerman 1943; Ter-Minasian 1953); however, a few more recently published reviews do include comprehensive illustrative keys for restricted regions, such as South Korea (Park et al. 2009) and Japan (Morimoto and Miyakawa 1995).
The genus Acicnemis is greatly in need of an exhaustive revision, and such a project will undoubtedly uncover many new species and synonyms. However, this would be an enormous undertaking given the high diversity in the genus (> 220 described species), lack of modern treatments of the group, and the costly necessity of viewing and dissecting type material that is deposited widely in natural history collections across Europe and Asia. In response to these challenges, we develop a preliminary, Japan-focused review of the East Asian species of Acicnemis, building on the work of Morimoto and Miyakawa (1995) that covered only the Japanese species. In addition to Japan, we now also include species from Taiwan, the Korean Peninsula, and China, as a foundation for future work on species in Southeast Asia, the epicenter of Trachodini diversity. We review important external morphological and genital characters, present a key to the East Asian species, and use DNA (CO1) barcoding to supplement our morphology-based species hypotheses.
Materials and methods
Specimen acquisition and general methodology
Specimens were examined from the following collections:
NHMUK Natural History Museum, London, England
CMIC Natural History Museum and Institute, Chiba, Japan
CMNC Canadian Museum of Nature, Gatineau, Canada
CNCI Canadian National Collection of Insects, Ottawa, Canada
ELKU Entomological Lab of Kyushu University, Fukuoka, Japan
HUM Hokkaido University Museum, Sapporo, Japan
KPMNH Kanagawa Prefectural Museum of Natural History, Odawara, Japan
KUM Kyushu University Museum, Fukuoka, Japan
LBM Lake Biwa Museum, Kusatsu, Japan
NHMB Natural History Museum Basel, Basel, Switzerland
NMNS National Museum of Nature and Science, Tsukuba, Japan
OIST Okinawa Institute of Science and Technology, Tancha, Japan
RUMC Ryukyu University Museum Collection, Nishihara, Japan
SDEI Senckenberg Deutsches Entomologisches Institut, Müncheberg, Germany
SNSD Senckenberg Naturhistorischen Sammlungen Dresden, Dresden, Germany
TUA Tokyo University of Agriculture, Atsugi, Japan
USNM National Museum of Natural History, Washington, U.S.A.
ZIN Zoological Institute of Russian Academy of Sciences, St. Petersburg, Russia
ZMH Zoologisches Museum Hamburg, Hamburg, Germany
We first created an exhaustive list of all described Acicnemis by reviewing the literature and examining as many major specimen collections as possible. We also examined name-bearing type material for all species presented here, as well as > 90% of the Acicnemis species not treated here. We base our taxonomic decisions on the phylogenetic species concept (Wheeler and Platnick 2000), and provide unique combinations of both morphology and molecular (CO1) characters that define the species covered in this paper. Examined specimens without institutional unique specimen identifier (USI) labels were assigned labels that read in the form: JHL_AREV_###, JHL_ACITAI_###, or JHL_DNA_###. In some older collections, we encountered that multiple specimens were glued to the same point with a single data label; in these cases we assigned a single USI to represent the entire lot of specimens on the same pin. Specimens were relaxed and dissected by removing the abdomen, and genitalia were cleared in a solution of KOH and water. The genitalia were placed in a small vial of glycerin and attached to the same pin as their respective dissected specimens. Images of genitalia were taken with a Nikon DS-Fi3 camera through a Nikon SMZ18 stereomicroscope using NIS-Elements D v. 5.41.00 (Nikon Corporation, Yokohama, Japan). All other images were taken under a Leica M205 C microscope with a Leica DMC 5400 camera and stacked using Leica Application Suite (Leica Microsystems, Wetzlar, Germany). We follow the morphological terminology of Oberpreiler et al. (2014). X-ray μCT is used to visualize cryptic morphology in a few species; we use the same methodology as Lewis et al. (2024) with scan settings as follows for the two specimens: A. ryukyuana Lewis, 2023 (OKENT0053498) [Mag: 4X, Exposure: 1.1 sec., Source dist.: 20.19 mm, Detector dist.: 8.65 mm, Voltage: 50 kV, Power: 4 W] and A. albofasciata (Ter-Minasian, 1953) (JHL_AREV_782) [Mag: 0.4X, Exposure: 3 sec., Source dist.: 15.04 mm, Detector dist.: 113 mm, Voltage: 50 kV, Power: 4 W].
DNA barcoding
The widely used barcode gene cytochrome c oxidase subunit I (CO1) has been successfully used to delineate weevils, and even short fragments of this gene can be informative in species delineation (e.g., Lewis et al. 2024). We sequenced a short ~300 bp long fragment of cytochrome c oxidase subunit I (CO1) from twelve Acicnemis species (A. albofasciata, A. azumai Morimoto & Miyakawa, 1995, A. dividicincta Morimoto & Miyakawa, 1995, A. exilis Morimoto & Miyakawa, 1995, A. laeta Hubenthal, 1919, A. luteomaculata Morimoto & Miyakawa, 1995, A. maculaalba Roelofs, 1875, A. nohirai Morimoto & Miyakawa, 1995, A. postica Hubenthal, 1919, A. ryukyuana, A. sauteri Hubenthal, 1919, and A. squamata sp. nov.) to illustrate the utility of this shortened CO1 fragment in weevil delineation, as well as to complement our morphology-based species hypotheses. In total, we sequenced 24 Acicnemis specimens and six outgroup taxa. For each Acicnemis species, we made efforts to include at least two specimens from different localities to account for intraspecific CO1 variation.
DNA was extracted using the non-destructive methods of Oberacker et al. (2019). The forward MCO-IntF (GGWACWGGWTGAACWGTWTAYCCYCC) (Leray et al. 2013) and reverse Fol-degen-rev (TANACYTCNGGRTGNCCRAARAAYCA) (Yu et al. 2012) primers were used in a PCR mix with the following thermal profile: Initial (95 °C – 15:00 min.), Denature (94 °C – 0:30 min.), Annealing (46 °C – 1:30 min.), Extension (72 °C – 1:30), Repeat 35X, Final hold (4 °C – Infinite). The PCR products were sequenced with Illumina MiSeq using 600-cycle v3 kits. The CO1 fragments were processed and assembled in Geneious Prime (v. 11.0.14.1; Dotmatics, Boston, Massachusetts, United States of America). We used BLAST to confirm that non-target (non-weevil) DNA was not erroneously used and constructed neighbor-joining trees to ensure that members of the same species (judged by morphology) clustered together. DNA sequence data was aligned using MUSCLE (v. 3.8.425 with default settings; Edgar 2004) using default settings and quality-controlled by examining translated amino-acid alignments. MEGA 11 (v. 11.0.13; Tamura et al. 2021) was used to calculate pairwise K2P distances for all sequences and were used to calculate intra- and interspecific ranges and averages in the sequenced Acicnemis species.
For the gene-tree, ModelFinder (Kalyaanamoorthy et al. 2017) was used to select a substitution model (GTR+F+I+G4). A Maximum Likelihood (hereafter ML) tree of the twelve Acicnemis species was constructed in IQ-TREE (v. 1.6.12; Nguyen et al. 2015) with a heuristic search of 100,000 initial trees. Standard nonparametric bootstrap (hereafter BS) values were calculated from 1000 replicates. For outgroup taxa, we used the molytine weevil species Aphanerostethus japonicus Lewis & Kojima, 2024 (Cryptorhynchini Schoenherr, 1825), Colobodes ornatoideus Morimoto, 1988 (Sophrorhinini Lacordaire, 1865), Deretiosus albicaudatus Morimoto, 1988 (Sophrorhinini Lacordaire, 1865), Ectatorhinus adamsii Pascoe, 1871 (Ithyporini Lacordaire, 1865), Protacallodes ryukyuensis Morimoto, 2011 (Ithyporini Lacordaire, 1865), and Tylodina (tribe) sp. 1 (Cryptorhynchini Schoenherr, 1825). All CO1 barcodes were uploaded to GenBank (Accession numbers: PP110445.1, PP110471.1, PP110442.1, PP110479.1, PP110447.1, PP110480.1, PV255624–PV255644). Three specimens that we examined were sequenced as part of another project and their sequence data were retrieved from BOLD Systems (Accession numbers: MEBLA191-14, OWEVG891-15, VVGPL2845-15).
Results
General remarks
Based on an examination of 934 specimens from 19 major Asian, European, and North American natural history collections, we documented 25 Acicnemis species from East Asia including three hitherto overlooked new species from mainland Japan: A. cryptica Lewis & Kojima, sp. nov., A. koguma Lewis & Kojima, sp. nov., and A. squamata Lewis & Kojima, sp. nov. The former two new species (A. cryptica and A. koguma) are cryptic and sister to more widespread, previously described species (A. albofasciata and A. kiotoensis, respectively). Based on morphological considerations, we also propose the following subjective synonymies: Acicnemis cruciata Kleine, 1924 syn. nov. under Acicnemis postica Hubenthal, 1919, Acicnemis yakushimana Morimoto & Miyakawa, 1995 syn. nov. under A. dividicincta Morimoto & Miyakawa, 1995, and Acicnemis dorsonigrita Voss, 1941 syn. nov. under Acicnemis palliata Pascoe, 1872.
Within the 300 bp CO1 alignment, IQ-TREE identified 125 parsimony informative sites. In our CO1 gene tree, specimens of all twelve Acicnemis species clustered together (BS ≥ 95) as expected based on morphology (Fig. 1). Nodes beyond the species level were weakly supported (BS ≤ 55); however, this is not unexpected given that we only analyzed a single 300-bp long fragment of a relatively rapidly evolving mitochondrial marker (see Townsend et al. 2012). Although a proper phylogenetic analysis (in prep) is clearly required to make robust statements about phylogeny, the weak association (BS: 38) between species such as A. sauteri and A. maculaalba is expected as these form a species complex of brown, white-spotted species (see A. maculaalba remarks section) and are frequently confused with each other. The K2P distances (see Suppl. material 1) show little intraspecific variation within Acicnemis species (range: 0–0.10193; t-distributed 95% confidence interval: 0.0199 ± 0.0198), and a trend of greater K2P distance in species where representative specimens were collected at distant localities (i.e., different prefectures or countries such as A. postica). Interspecific K2P distances between Acicnemis species ranged from 0.10771–0.27405 (t-distributed 95% confidence interval: 0.1869 ± 0.0036), and were lowest between the related A. maculaalba and A. sauteri.
Maximum likelihood tree of twelve Acicnemis species based on a 300 base-pair long CO1 gene fragment constructed in IQ-TREE v. 1.6.12 using a GTR+F+I+G4 model. Branch support values represent standard nonparametric bootstraps (1000 replicates). Bootstrap values less than 75 are not displayed on the tree. The codes MEBLA191-14, VVGPL2845-15, and OWEVG891-15 are BOLD Systems accession numbers, whereas all other codes are GenBank accession numbers. Associated Acicnemis weevil figures on the right of the tree are approximately to scale.
Morphology
The East Asian species of Acicnemis are relatively uniform morphologically across the genus, but do differ prominently in scale color and pattern, which is highly conserved within species. They also markedly differ in male genital morphology, and examination of the genitalia is critical for identification of some cryptic species covered here (e.g., A. albofasciata and A. cryptica). There are, however, several other important external morphological characters that are critical to Acicnemis species identification that we summarize below.
Third tarsomeres
The third tarsomeres are emarginate (e.g., Fig. 2G) in most Acicnemis species (e.g., A. albofasciata, A. azumai); however, a few species covered here have truncate (A. luteomaculata (Fig. 2E) and A. koguma sp. nov.) or weakly emarginate third tarsomeres (A. ryukyuana) (Fig. 2F).
Diagnostic morphological characters in Acicnemis. A. A. azumai Morimoto & Miyakawa, 1995 (JHL_ACITAI_081) showing lack of prominent posterior prosternal projections; B. A. laeta Hubenthal, 1919 (JHL_AREV_533) showing prominent prosternal projections; C. A. kiotoensis Nakane, 1963 (JHL_AREV_982) showing antenna with the second funicular antennomere longer than antennomeres 3 + 4; D. A. koguma Lewis & Kojima, sp. nov. (JHL_SYN_141) showing antenna with the second funicular antennomere shorter than antennomeres 3 + 4; E. A. luteomaculata Morimoto & Miyakawa, 1995 (JHL_AREV_189) showing truncate third tarsomere; F. A. ryukyuana Lewis, 2023 (JHL_ACI_011) showing weakly emarginate third tarsomere; G. A. maculaalba Roelofs, 1875 (JHL_AREV_577) showing strongly emarginate third tarsomere.
Antennomeres
The proportions of funicular antennomere lengths vary between Acicnemis species. For example, in A. kiotoensis Nakane, 1963 the second funicular antennomere is longer than funicular antennomeres 3+4 (Fig. 2C); however, in the closely related species A. koguma the second funicular antennomere is shorter than funicular antennomeres 3+4 (Fig. 2D).
Pronotum
One species covered here (A. azumai) has unscaled, baso-medial tubercles on the pronotum (Fig. 3A). These are diagnostic for this species and can be used to quickly distinguish A. azumai from the superficially similar A. laeta and much smaller species A. exilis. Furthermore, some species have an impunctate longitudinal midline on the pronotum (e.g., A. koguma, some specimens of A. albofasciata and A. cryptica).
Scutella and pronota of various Acicnemis species. A. A. azumai Morimoto & Miyakawa, 1995 (JHL_ACITAI_081) showing two bare (unscaled) tubercles baso-medially on the pronotum; B. A. kiotoensis Nakane, 1963 (JHL_AREV_982) showing a broad, unscaled scutellum; C. A. koguma Lewis & Kojima, sp. nov. (JHL_SYN_141) showing a bare, but more slender scutellum than A. kiotoensis; D. A. maculaalba Roelofs, 1875 (JHL_AREV_577) showing a small scutellum covered in scales; E. A. palliata Pascoe, 1872 (JHL_AREV_779) showing a larger scutellum that is shiny and bare dorsally, and surrounded along the perimeter by yellow scales; F. A. shigematsui Morimoto & Miyakawa, 1995 (JHL_ACITAI_052) showing a smaller scutellum covered in scales.
Prosternum
The Taiwanese species A. laeta has diagnostic prosternal projections that lie posterior to the fore-coxae (Fig. 2B). This rare character is not found in any other species covered here but is shared with a few Southeast Asian species.
Scutellum
The morphology of the scutellum is extremely variable and very important in Acicnemis species identification. Some species have a large, shiny scutellum (e.g., A. palliata Pascoe, 1872) (Fig. 3E), while others have a heavily scaled one (e.g., A. azumai) (Fig. 3A). In some instances, scutellum morphology can be used to easily differentiate similar species (e.g., A. kiotoensis vs. A. maculaalba Roelofs, 1875) (Fig. 3B, D).
Sclerolepidia
Variation in the sclerolepidia has been documented in Acicnemis previously (Lyal et al. 2006) (Fig. 4). Although we examined our two newly described cryptic species and their associated sister species (A. albofasciata – A. cryptica and A. kiotoensis – A. koguma) for variation in sclerolepidia morphology, we did not observe any significant differences. We assert that the sclerolepidia are useful primarily for broadly placing the species into groups, but not useful for separating closely related species.
Sclerolepidia in Acicnemis. A. A. kiotoensis Nakane, 1963 (JHL_AREV_982) showing low, rounded sclerolepidia; B. A. palliata Pascoe, 1872 (JHL_AREV_779) showing protruding, erect sclerolepidia.
Erect elytral scales
Many Acicnemis species have erect elytral scales that vary in length, position, and patterning. In particular, some species only have these erect scales on the odd-numbered elytral intervals (e.g., A. shigematsui Morimoto & Miyakawa, 1995), while others have erect scales on the odd-numbered intervals and the second interval (e.g., A. luteomaculata). Some East Asian species also have exceptionally long erect elytral scales (e.g., A. ryukyuana).
Pronotum punctation (cryptic morphology; requires X-ray μCT)
Pronotum punctation differs between species in some instances; however, this character is generally not visible as the pronotum is often covered by scales. We used X-ray μCT (as in Lewis et al. 2024) to strip away the pronotal scales in A. ryukyuana and A. albofasciata and found differences in puncture spacing (and pronotal sculpturing). In A. ryukyuana (Fig. 32A) the pronotum is smooth and the punctures are well spaced, whereas in A. albofasciata (Fig. 32B) the pronotum is coarsely sculptured and the punctures are closely spaced. We did not encounter any differences between closely related sister species (e.g., A. albofasciata – A. cryptica or A. kiotoensis and A. koguma), indicating that this pronotum punctation and our X-ray μCT methodology is only useful for more delineating more distantly related Acicnemis species pairs.
Metanotum + dorso-lateral flight muscles (cryptic morphology; requires X-ray μCT)
Using X-ray μCT, we visualized the metanotum and dorso-longitudinal flight muscles (DLFM), which moderately differ in proportion to the elytra in A. ryukyuana and A. albofasciata. In A. ryukyuana (Fig. 32C) the metanotum + DLFM are 30% the length of the elytra, whereas in A. albofasciata (Fig. 32D) they are 23% the length of the elytra. Although most Acicnemis species share a similar, elongate body profile to A. albofasciata, some species are more rounded and compact like A. ryukyuana. The evolution of a more compact body plan and abrupt apical elytral declivity in A. ryukyuana apparently had little effect on the relative size of the metanotum and DLFM, hence the difference in proportional length between species.
Taxonomy
Acicnemis
Taxon classificationAnimaliaColeopteraCurculionidae
Fairmaire, 1849
D31A9449-1053-581F-892C-FEBD2F63C355
Oplocnemus Dejean, 1835 (Type species: Oplocnemus mucronatus Buquet nomen nudum). Hoplocnemus Agassiz, 1846 (unjustified emendation of Oplocnemus). Acienemis Motschulsky, 1863 (incorrect subsequent spelling). Berethia Pascoe, 1872 (Type species: Berethia medinotata Pascoe, 1872). Acicnemus Haly, 1890 (incorrect subsequent spelling).
Type species.
Acicnemis variegata Fairmaire, 1849 (by subsequent designation).
Gender.
Feminine.
Description.
Body length 2.0–12.0 mm. Scale pattern: Usually cryptically patterned with contrasting brown, black, gray, and dull yellow scales. Pronotum and elytra often with erect scales. Head: Rostrum evenly curved in most species, and variably longer in females than in males; eyes large, ovate, and non-contiguous; antennae with seven-articled funicle. Prothorax: Densely punctate; pronotum often with tufts of erect scales; prosternum lacking well defined prosternal canal. Elytra: Scutellum varied (large, plate-like, naked scutellum in some species, or small and covered in scales in others); erect scales often only on odd-numbered elytral intervals; elytra unmodified in most species, costate in a few; elytra with apical projections in some species (A. laticollis Pascoe, 1885); low tubercles along elytral intervals in some species, absent in most; volant, hind wings functional. Thorax: Sclerolepidia varied: prominent and protruding in some species, forming low, indistinct tubercles in others. Abdomen: First and second sternites variably concave medially in males (convex or flat in females). Legs: Femora all bearing a large ventral tooth (simple in most species, serrated in a few); forelegs elongate in some species (e.g., A. longimana Hubenthal, 1922); third tarsomeres emarginate (bilobed) in most species, truncate in a few; tarsal claws simple.
Distribution.
Acicnemis species are broadly distributed across the Palearctic Region (Azerbaijan, China, Georgia, Japan, Korean archipelago, Russia, Taiwan), the Oriental Region (Cambodia, India, Indonesia, Laos, Malaysia, Myanmar, Philippines, Thailand, Vietnam), the Australasian Region (Australia, Fiji, New Guinea, Samoa, Tahiti, Tonga), and apparently also the Afrotropical Region (Nigeria) (Hubenthal 1919a, b; Zimmerman 1967; Alonso-Zarazaga and Lyal 1999; Kojima and Morimoto 2004; Oziegbe and Faluyi 2010; Pullen et al. 2014; Alonso-Zarazaga et al. 2023).
Ecology and natural history.
The larvae of Trachodini mine through dead wood (Lyal 2014). Host plant associations are not well known in Acicnemis; however, a few records exist (Acicnemis palliata Pascoe, 1872 with Wisteria floribunda (Willd.) DC. and W. sinensis (Sims) DC. (Fabaceae) (Morimoto and Miyakawa 1995; Lee and Morimoto 1996; Zhang 2021); Acicnemis suturalis Roelofs, 1875 with Wisteria brachybotrys Siebold & Zucc. (Morimoto and Miyakawa 1995; Lee and Morimoto 1996); Acicnemis crassiusculus Fairmaire, 1878 with Hibiscus tiliaceus L. (1753) (Malvaceae) and Ludwigia spp. (Onagraceae) (Zimmerman 1943; Oziegbe and Faluyi 2010); the larvae of one unidentified Acicnemis species were taken from a dead stem of Castanopsis sieboldii (Makino) Hatus (Fagaceae) (Lee and Morimoto 1996). Some species are attracted to lights (e.g., A. azumai) (JHL pers. obs.).
Species profiles
Acicnemis
albofasciata
Taxon classificationAnimaliaColeopteraCurculionidae
(Ter-Minasian, 1953)
AF5DBBB6-FD29-5147-8C1E-7F8AB32201D9
Trachodes albofasciata Ter-Minasian, 1953: 317. Acicnemis nigra Nakane, 1963: 37.
Type material examined
(Russia: 8; Japan: 173; Total: 181). Lectotype (designated here). Russia • Primorsky Krai, Lyanchihe (26 km from Vladivostok), 19.V.1949, on Betula manshurica, ZIN, JHL_AREV_997. Paralectotype. Russia • Primorsky Krai, Partizansk (formerly Suchan), VIII.1925 (1, ZIN), JHL_AREV_996.
Acicnemis albofasciata (Ter-Minasian, 1953) (JHL_AREV_781). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Russia • Primorsky Krai, Benevskoye, 43.17°N, 133.76°E, 4.VII.2014, E. Jendek (2, CNCI), CNCCOLVG00007390 (BOLD accession number: VVGPL2845-15), CNCCOLVG00008936 (dissected male, BOLD accession number: OWEVG891-15); • S. Primor’ye, Kedrovaya Pad Reserve, 20.V.–2.VI.1992, T. Nakamura (3, KUM), JHL_AREV_499 (dissected male), JHL_AREV_782, JHL_AREV_787; • Saghalien, Kawakami (present day Sinegorsk), 7.VIII.1922, T. Esaki (1, KUM), JHL_AREV_899; Japan: Akita Prefecture: • Tamagawa, Tazawako-machi, 10.VII.1976, S. Miyakawa (12, TUA), JHL_AREV_674 (dissected male), JHL_AREV_811 – JHL_AREV_814, JHL_AREV_935; • Tamagawa Onsen, 10.VII.1976, S. Miyakawa (4, TUA), JHL_AREV_932 (dissected male), JHL_AREV_934; • Senboku, Nyutou-onsen, 10.VI.2015, K. Takahashi (1, KUM), JHL_AREV_887; • Takada-odake, 14.VII.1962, Y. Miyatake (1 dissected male, KUM), JHL_AREV_898; • Sengan-toge, 14.IX.1974, S. Miyakawa (2, TUA), JHL_AREV_965 (dissected male); Aomori Prefecture: • Towada, 12.VI.1955, K. Shimoyama (1 dissected male, ZMH), ZMH 837198; • Nurukawa, 12.VIII.1950, K. Shimoyama (1, ZMH), ZMH 837199; • Kobokutai, 5.VIII.1937, K. Shimoyama (1, ZMH), ZMH 837200; • Yunomata, Ohata-machi, Shimokita Penn., 14–30.VII.1956, K. Morimoto (7, KUM), JHL_AREV_543 (dissected male), JHL_AREV_793 – JHL_AREV_797, JHL_AREV_846; Fukushima Prefecture: • Tateiwa, 22–23.VI.1984, A. Saito (2, CMIC), CBM–ZI 68470 (dissected male), CBM–ZI 68471; • Minami-aizu-machi, Yaso, 24.VI.2020, T. Saeki (5, KUM), JHL_AREV_882 – JHL_AREV_886; • Shindenhara, 28.V.1979, K. Emoto (1 dissected male, KUM), JHL_AREV_850; Gifu Prefecture: • Gifu (1, NHMB), JHL_SYN_192; • Hirayu, 29.VII.–28.VIII.1959, M. Sato (2, KUM), JHL_AREV_102, JHL_AREV_865; • Yunohana, 22.VI.1956 (1 dissected male, CMNC), JHL_AREV_112; Gunma Prefecture: • Sugenuma, 10–11.VII.1975, H. Irie (1, KUM), JHL_AREV_869; • Nikamatazawa, Mt. Shirane, 16–18.VII.1975, H. Irie (1 dissected male, KUM), JHL_AREV_918; • Fujimi-toge, 4.VIII, M. Samejima (8, TUA), JHL_AREV_961 (dissected male), JHL_AREV_962, JHL_AREV_964; • Hokkaido: “Hokkaido”, “6.19.53”, “Arakawa” (1, NHMB), JHL_SYN_190; • Sounkyo, 11.VII1986, S. Nomura (3, KUM), JHL_SYN_189; • Aizankei, 29.VII.1964 (1, KUM), JHL_AREV_103; • Aizankei, Mt. Daisetsu, 27–31.VII.1955, K. Morimoto & S. Kimoto (14, KUM), JHL_AREV_501 (dissected male), JHL_AREV_798, JHL_AREV_799, JHL_AREV_801 – JHL_AREV_810, JHL_AREV_855; • Teshio, 4.VII.1916, T. Isshiki (1 dissected male, HUM), JHL_AREV_101; • Ashoromura (Tokachi), 29.VII.–23.VIII.1949, R. Matsuda (5, KUM), JHL_AREV_781, JHL_AREV_888 – JHL_AREV_891; • Mitsumata-rindo, Kamishihoro-cho, 25.VI.1986, S. Miyakawa & K. Morimoto (1, TUA; • 2, KUM), JHL_AREV_542, JHL_AREV_815, JHL_AREV_816; • Ashorobuto, Ashoro-gun, 24–31.V.1957, M. Takahashi (3, KUM), JHL_AREV_480 (dissected male), JHL_AREV_847, JHL_AREV_848; • Mt. Daisetsuzan, 26.VII.1953, Y. Kurosawa (2, NMNS), JHL_AREV_852, JHL_AREV_853; • Daisetsuzan, 1.VIII.1962 (3, KUM), JHL_ACITAI_008 (dissected male), JHL_AREV_104, JHL_AREV_105; • Jushichinosawa, Mt. Nipesotsu, 6.VIII.1976, H. Irie (1, KUM), JHL_AREV_856; • Nakayama-toge (near Sapporo), 22.VI.1986, K. Morimoto (1, KUM), JHL_AREV_857; • Bibai, 12.V.1962, K. Kamijo, collected from Populus sp. (1, KUM), JHL_AREV_858; • Oyukiyama, 31.VII.1952, Yoshida (1, KUM), JHL_AREV_859; • Aizankei, Antaroma, 31.VII.1952, T. Shirozu (1, KUM), JHL_AREV_861; • Lake Akan, Kushiro, 8.VI.1957, M. Takahashi (1, KUM), JHL_AREV_862; • Wassakanai, Sohya-shichou, 26.VII.1991, S. Ohmomo (1, KUM), JHL_AREV_863; • Nishiashoro (Tokachi), 17.VIII.1949, R. Matsuda (1, KUM), JHL_AREV_864; • Yukomanbetsu, Mt. Daisetsu, 25.VII.1955, K. Morimoto (1, KUM), JHL_AREV_866; • Horoka, Kamishihoro-cho, 17.VII.1976, H. Irie (1, KUM), JHL_AREV_867; • Kamiotoineppu (Teshio), 24.VIII.1922, T. Esaki (4, KUM), JHL_AREV_868, JHL_AREV_877 – JHL_AREV_879; • Kawakami (near Honbetsu), 17–27.VII.1953, Y. Hirashima (1, KUM), JHL_AREV_870; • Aizankei, 19.VII.1962, Y. Miyatake (1, KUM), JHL_AREV_871; • Aizankei (Ishikari), 29.VII.1952, T. Shirozu (5, KUM), JHL_AREV_872 – JHL_AREV_876; • Nakarikubetsu, Rikubetsu Town, Asyoro County, 24.VII.2017, K. Narita (2, KUM), JHL_AREV_880, JHL_AREV_881; • Meakan-dake, Akan National Park, 5.VII.1958, S. Miyamoto (4, KUM), JHL_AREV_892 – JHL_AREV_895; • Uenae, Tomakomai City, 15.VIII.1989, H. Kojima (6, TUA), JHL_AREV_922 (dissected male), JHL_AREV_910 – JHL_AREV_914; • Mt. Karibayama, Shimamaki, 24–27.VII.1989, H. Kojima (2, TUA), JHL_AREV_924, JHL_AREV_926; • Mt. Daisengen, Hakodate City, 23.VII.1989, H. Kojima (1, TUA), JHL_AREV_925; • Poroshirisanso, Hidaka, 5.VIII.1988, H. Kojima (2, TUA), JHL_AREV_927, JHL_AREV_928; • Kamishihoro-chou, Mitsumata, 17.VII.2014, S. Yamamoto, JHL_DNA_154 (GenBank accession number: PV255624) (1, KUM); Ibaraki Prefecture: • Mt. Tsukuba, 22.VIII.1977, A. Tanaka (1, HUM), JHL_AREV_854; Ishikawa Prefecture: • Mt. Hakusan, 18.V.1979, I. Togashi & D.R. Smith (1 dissected male, USNM), JHL_AREV_985; Iwate Prefecture: • Iwate, VI.1936, H. Yamamoto (2, NHMB), JHL_SYN_188, JHL_SYN_191; Miyagi Prefecture: • Nuruyu, Hanayama mura, 2–6.VIII.1984, K. Morimoto (1 dissected male, KUM), JHL_AREV_849; • Mt. Funagata, Taiwa-machi, 21.VIII.2009, J. Aoki (2, KUM), JHL_AREV_839 (dissected male); Nagano Prefecture: • Nakabusa, Shinano, 24.VII, T. Samejima (1 dissected male, TUA), JHL_AREV_673; • Bandoko, Shinano Province, 13.VII.1935, H. Kiyosawa (4, KUM), JHL_AREV_917 (dissected male), JHL_AREV_919 – JHL_AREV_921; • Shirahone, 19.VII.1956, S. Kimoto (2, KUM), JHL_AREV_593 (dissected male), JHL_AREV_916; • Shirahone, 12.VIII.1960, Y. Kimura (1 dissected male, KUM), JHL_AREV_838; • Kamikouchi, 14.VII.1929, M. Samejima (4, TUA), JHL_AREV_783 (dissected male), JHL_AREV_967 – JHL_AREV_969; Niigata Prefecture: • Sasagamine, 23.VIII.1962, H. Koike (1 dissected male, KUM), JHL_AREV_896; Tochigi Prefecture: • Yumoto, Oku-nikko, 7.VII.1950, M. Takahashi (3, KUM), JHL_AREV_902 (dissected male), JHL_AREV_906, JHL_AREV_907; • Yumoto, 30.VII.1960, K. Suga (1 dissected male, KUM), JHL_AREV_903; • Chu-zenji, 16.VII.1916, E. Gallois (3, HUM), JHL_AREV_851; • Tokura, Nikko, 16.VI, S. Miyakawa (3, TUA), JHL_AREV_960 (dissected male); Yamanashi Prefecture: • Daibosatsu, 22–23.VI.1985, K. Yoshihara (6, TUA), JHL_AREV_629 (dissected male), JHL_AREV_788, JHL_AREV_789, JHL_AREV_790 – JHL_AREV_792; • Hikawa-rindo, Mts. Daibosatsu, 22.VII.1983, S. Miyakawa (21, TUA), JHL_AREV_946 (dissected male), JHL_AREV_948 – JHL_AREV_958; • Kanayama, 1.VII.1989, T. Tanaka (1 dissected male, TUA), JHL_AREV_923; • Kanayamadaira, 28–30.VI.1991, H. Urushihara (2, TUA), JHL_AREV_930 (dissected male), JHL_AREV_933; • Koshu-shi, Enzankamiigiwara, 13.VIII.2016, S. Shimamoto (2, TUA), JHL_AREV_937 (dissected male), JHL_AREV_940; • Koshu-shi, Enzankamiigiwara, 3.V.2015, S. Shimamoto (1, TUA), JHL_AREV_939; Unknown Prefecture: • Ichi-no-hashi, 29.VII.1975, S. Ogata (1 dissected male, KUM), JHL_AREV_502.
Diagnosis.
Body length 5.0–6.8 mm. Covered in black and dark gray scales, with longitudinal, black-bordered white stripes across the lateral edges of the pronotum and elytral humeri (compare with A. cryptica which has the entire lateral portion of the pronotum covered in white to yellow scales). Apical 1/2 of elytra with a V-shaped patch of white scales. Pronotum evenly punctured across dorsal surface, but often with shiny, impunctate longitudinal midline. Scutellum bare over anterior 1/2 of dorsal surface, otherwise covered in pale yellowish scales. Only odd elytral intervals with erect scales (even elytral intervals may possess erect scales at the extreme elytral apices). Sclerolepidia distinct and protruding. Third tarsomeres distinctly emarginate. Pedon with lateral edges approx. parallel in dorsal view, and converging evenly at apex into a dull, rounded point. Internal sac visible in anterior 1/2 of aedeagus and tuberculate (roughed appearance); • internal sac also with hook-like protruding structure anteriorly (at the apex) and surrounded by tuberculate, roughened musculature (Fig. 6A1–2).
Aedeagi of Acicnemis. A1–2. A. albofasciata (Ter-Minasian 1953); B1–2. A. azumai Morimoto & Miyakawa, 1995; C1–2), A. cryptica Lewis & Kojima, sp. nov.; D1–2. A. dividicincta Morimoto & Miyakawa, 1995; E1–2. A. exilis Morimoto & Miyakawa, 1995; F1–2. A. kiotoensis Nakane, 1963; G1–2. A. laeta Hubenthal, 1919; H1–2. A. luteomaculata Morimoto & Miyakawa, 1995; I1–2. A. maculaalba Roelofs, 1875; J1–2. A. nobilis Hubenthal, 1919; K1–2. A. nohirai Morimoto & Miyakawa, 1995; L1–2. A. palliata Pascoe, 1872; M1–2. A. postica Hubenthal, 1919; N1–2. A. ryukyuana Lewis, 2023; O1–2. A. sauteri Hubenthal, 1919; P1–2. A. shibatai Voss, 1971; Q1–2. A. shigematsui Morimoto & Miyakawa, 1995; R1–2. Acicnemis sp. 1; S1–2. A. squamata Lewis & Kojima, sp. nov.; T1–2. A. suturalis Roelofs, 1875.
Distribution.
This species occurs in far Eastern Russia (Primorsky Krai, Saghalien) and north-central mainland Japan (from Hokkaido south to Gifu Prefecture).
Remarks.
The two A. albofasciata syntypes deposited in the ZIN Collection are apparently both females, despite the original paper describing a male (B. Korotyaev to JHL, pers. comm. 2025). Dissection of males is required to conclusively differentiate between A. albofasciata and its sister species A. cryptica. However, based on our extensive collection examinations, we found that Acicnemis albofasciata occurs in the Far East (Primorsky Krai, Russia), Hokkaido, and northern mainland Japan, while A. cryptica occurs only in mainland Japan, from Aomori Prefecture south to Kyushu (Oita Prefecture). Furthermore, examined specimens of A. albofasciata have a black-scaled, lateral stripe on the pronotum, while A. cryptica have the lateral portion of the pronotum mostly covered in yellow to white scales. We are confident that the Ter-Minasian syntypes belong to A. albofasciata (not A. cryptica) as only this species apparently occurs outside of Japan (with known records from the syntype locality (Primorsky Krai, Russia), and as the Ter-Minasian syntypes have a black-scaled lateral pronotum band. To fix the identity of A. albofasciata, we hereby designate one of the Ter-Minasian syntypes (JHL_AREV_997) as a lectotype.
Acicnemis
azumai
Taxon classificationAnimaliaColeopteraCurculionidae
Morimoto & Miyakawa, 1995
8BBE2064-DEF1-5BE5-BFE5-6BF7AC8123C5
Type material examined
(Japan: 82; South Korea: 3; Total: 84). Holotype. Japan: Okinawa Prefecture: • Kudeken, Chinen-son, 22.IV.1975, H. Irie, bears red label reading “(HOLOTYPE) Acicnemis azumai Morimoto et. Miyakawa, 1994”, JHL_AREV_113, ELKU, ELKU 2962. Paratypes. Japan: Okinawa Prefecture: • Sueyoshi, “1988-6-5”, T. Ueno (5, KUM), JHL_ACITAI_041 (dissected male), JHL_AREV_134 – JHL_AREV_137; • Yona, 9–13.VIII.1969, H. Makihara (4, KUM), JHL_AREV_108 – JHL_AREV_111; • Shuri, 14.VI.1958, O. Nakachi (1, KUM), JHL_AREV_138; • Shuri, 16–20.V.1978, H. Makihara (1, KUM), JHL_AREV_117; • Aguni Island, 24–25.VIII.1989, Teruhisa Ueno, JHL_ACITAI_081 (dissected male) (4, KUM), JHL_AREV_124 – JHL_AREV_126; • Mabuni, 11.VI.1977, H. Irie (2, KUM) JHL_AREV_127, JHL_AREV_128; • Nago, 18.VI.1984, Teiso Esaki (1, KUM), JHL_AREV_130; • Akajima Island, 29–30.VIII.1980, Teruhisa Ueno (1, KUM), JHL_AREV_129; • Mount Banna, Ishigaki Island, 30.VII.1970, I. Matoba (1, KUM), JHL_AREV_131; • Tonaki Island, 27.IX.1989, Teruhisa Ueno (2, KUM), JHL_AREV_132, JHL_AREV_133; • 18.VI.1970, H. Makihara (2, KUM), JHL_AREV_122, JHL_AREV_123; • Tamagusuku, 4.V.1962, M. Kina (1, KUM), JHL_AREV_151; • Tamagusuku, 23.VIII.1962, M. Kina (2, KUM), JHL_AREV_152, JHL_AREV_153; • Otomi, Iriomote Island, 25.IV.1969, H. Makihara (1, KUM), JHL_AREV_143; • Shimozato, Miyako Island, 1.IX.1958, T. Hidaka (1, KUM), JHL_AREV_144; • Kudeken, 31.V.1976, T. Ogasawara (1, KUM), JHL_AREV_145; • Shuri, 5–9.V.1969, H. Makihara (1, KUM), JHL_AREV_146; • Naha, 13.VIII.1952, K. Hamamatsu (1, KUM), JHL_AREV_147; • Sueyoshi, 9.VII.1987, Teruhisa Ueno (1, KUM), JHL_AREV_148.
Acicnemis azumai Morimoto & Miyakawa, 1995 (OKENT0132427). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Okinawa Prefecture: • Iriomote Island, Mt. Tedo, 28.III.2022, Y. Obae (1, KUM), JHL_AREV_511; • Ishigaki Island, Inoda, 24.III.2022, S. Imada, M. Maruyama, T. Nozaki (1, KUM), JHL_AREV_512; • Kabira Park, Ishigaki Island, 6.VI.1968, M. Takagi (1, KUM), JHL_AREV_763; • Okinawa Island, Sakukawa, 8.XI.1971, T. Terya (1, RUMC), JHL_AREV_513; • Okinawa Island, Nishihara, 16.VII.1977, M. Arasaki (1, RUMC), JHL_AREV_514; • Okinawa Island, Yona, 24.IV.1965, T. Kakinohana (2, RUMC), JHL_AREV_515, JHL_AREV_516; • Okinawa Island, Kudeken, 28.VI.1977, K. Omato (1, RUMC), JHL_AREV_517; • Okinawa Island, Shuri, 7.VII.1972, M. Kinjo (1, RUMC), JHL_AREV_518; • Miyako Island, 1.IX.1958, T. Takara (3, RUMC), JHL_AREV_519 – JHL_AREV_521; • Okinawa Island, Shuri, 16.VI.1961, O. Nakachi (1, RUMC), JHL_AREV_522; • Okinawa Island, Shuri, 24.VIII.1962, O. Nakachi (1, RUMC), JHL_AREV_523; • Okinawa Island (1, OIST), JHL_AREV_984 (GenBank accession number: PV255625); • Tancha, Okinawa Island, 4.III.2025, J.H. Lewis, to residential lights (1, OIST), JHL_AREV_995; • Tancha, Okinawa Island, 14.III.2025, J.H. Lewis, to residential lights (1, OIST), JHL_AREV_993; • Tancha, Okinawa Island, 29.VIII.2025, J.H. Lewis, to residential lights (1, OIST), JHL_AREV_2081; • Sueyoshi Park, Naha, 12–26.II.2016, (1, OIST), OKENT0132427; South Korea • Oseag Ri, Ganweondo, 4–6.VII.1984, T. Senoh (1, KUM), JHL_AREV_155; • Sam Jeong Li, Ma Cheong Meon, Hamyang Gun, Gyeongsamnamdo, 9–15.V.1991, K. Morimoto (2, KUM), JHL_AREV_154, JHL_ACITAI_079 (dissected male).
Diagnosis.
Body length 3.3–4.9 mm. Covered in pale brown scales, with alternating white and black scaled bands on odd elytral intervals. Pronotum with two exposed, bare tubercles medially at the base (one on each side) (Fig. 3A). Scutellum covered in yellow scales. Only odd elytral intervals with erect scales. Sclerolepidia distinct and protruding. Posterior edge of prosternum lacking prominent projections contiguous with the fore-coxae (Fig. 2A) (present in A. laeta). Third tarsomeres distinctly emarginate. Pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a point. Internal sac without tuberculate or roughened musculature inside the aedeagus, but with a non-hooked protruding structure (unmodified in posterior 1/2, with roughened in anterior 1/2) that is equal in length to the pedon (Fig. 6B1–2).
Distribution.
This species occurs in southern Japan (Okinawa Prefecture, Ryukyu Islands) and South Korea.
Remarks.
On Okinawa Island (Japan) this species is commonly encountered in the spring and is often attracted to residential building lights.
Acicnemis
biarcuata
Taxon classificationAnimaliaColeopteraCurculionidae
Hubenthal, 1919
BAB38C43-B84D-5AD8-A739-F31F12F84438
Type material examined
(Taiwan: 5). Holotype. Taiwan • “Kosempo”, “Formosa”, “IX.09”, “Sauter”, bears red label reading “Holotypus”, SDEI, SDEIColeoptera #303414, SDEI Muncheburg COL – 18569.
Acicnemis biarcuata Hubenthal, 1919 (JHL_AREV_628). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Taiwan • Lienhoachii, “1975.5.12”, S. Imasaka (1, TUA), JHL_SYN_184; • Mudan Township, Pingtung County, 22°08'21.71"N, 120°51'31.23"E, 9.III.2013, Y. Fujisawa (1, TUA), JHL_AREV_626; • Mudan Township, Pingtung County, 22°06'12.57"N, 120°47'33.71"E, 6.III.2013, Y. Fujisawa (1, TUA), JHL_AREV_627; • Manzhou Township, Pingtung County, 22°08'30"N, 120°51'13"E, 2.V.2019, S. Shimamoto (1, TUA), JHL_AREV_628.
Diagnosis.
Body length 4.0–5.2 mm. Covered in dark-gray scales, with two V-shaped, transverse black bands across middle of elytra. Pronotum with pale brown and yellow scales dorsally, but dark at base medially. Pronotum approx. sculptured and carinate. Scutellum shiny and bare in examined specimens. Only odd elytral intervals with erect scales across elytral disk (some erect scales on 2^nd^ interval in apical fourth of elytra). Third and fifth elytral intervals with cluster of 3–5 erect scales at elytral base. Sclerolepidia distinct and protruding. Mesosternum with distinctly raised ridge between meso-coxae, highest laterally but dipping shallowly medially (no such ridge or only weakly developed in most other species). Third tarsomeres weakly emarginate.
Distribution.
This species is currently only known from Taiwan.
Acicnemis
bickhardti
Taxon classificationAnimaliaColeopteraCurculionidae
Hubenthal, 1919
A2A84396-C691-58FD-A08B-252BE9BD89DC
Type material examined
(Taiwan: 3). Holotype. Taiwan • Kosempo, H. Sauter, 1909, bears a red label reading “Type”, SNSD, JHL_ACITAI_001.
Acicnemis bickhardti Hubenthal, 1919 (JHL_AREV_591). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Taiwan • “16.VII.20”, T. Kano (1, NMNS), JHL_AREV_591; • 20.VI.1938, unknown collector (1, TUA), JHL_AREV_592.
Diagnosis.
Body length 5.0–7.0 mm. Easily distinguished from other East Asian Acicnemis by the (1) large size (2) black to dark red cuticle largely naked, but with bright yellow scaled bands running longitudinally across the lateral edges of the pronotum and elytral humeri, as well as transversally across the apical 1/2 of the elytra.
Distribution.
This species is known only from Taiwan.
Remarks.
Many similarly patterned, large species that presumably form a distinct clade within Acicnemis are present throughout Southeast Asia (e.g., A. ibis Faust, 1896 and A. peduncularis Pascoe, 1872), and a number of these remain undescribed.
Acicnemis
cordata
Taxon classificationAnimaliaColeopteraCurculionidae
Hubenthal, 1919
2C364F23-F2EB-5514-8197-EDD143888B42
Acicnemis fukienensis Voss, 1958.
Type material examined
(Vietnam: 1). Holotype. Vietnam • “Mau-son, Tongking” (modern day northern Vietnam), “Gehr. W. Muller, Vermacht, 1909”, bears a red label reading “Typus”, SNSD, JHL_AREV_987.
Acicnemis cordata Hubenthal, 1919 holotype (JHL_AREV_987). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Diagnosis.
Body length 5.0 mm. Virtually indistinguishable externally from A. maculaalba (see that species). Although most A. maculaalba have the white elytral spots interrupted by brown scales along the first elytral interval, some have no interruption (as in the A. cordata holotype); therefore, this character cannot be used alone to reliably separate A. maculaalba and A. cordata.
Distribution.
This species is known from the type locality (Mau-son, Tongking, which corresponds to modern day northern Vietnam) and apparently also Kuatun, China (Voss’s specimen).
Remarks.
We examined the holotype specimen of A. cordata Hubenthal, 1919 (collected from Fujian, China; SNSD) and found it is virtually indistinguishable externally from Taiwanese and Japanese specimens of A. maculaalba. However, as the A. cordata holotype is a female, it was not possible to examine the male genitalia, which are extremely important for distinguishing members of the A. maculaalba species complex. We tentatively recognize A. cordata here, but also note that further examination of specimens from mainland Asia is necessary to work out the species in this complex.
Acicnemis
costulifera
Taxon classificationAnimaliaColeopteraCurculionidae
Hubenthal, 1919
F2EFD8DB-464C-5BB4-9665-B1AE70E0D11E
Type material examined
(Unknown country: 1). Holotype. Unknown locality: “Coll. J. Faust Ankauf 1900”, bears a red label reading “Typus”, SNSD, JHL_ACITAI_003.
Acicnemis costulifera Hubenthal, 1919 (JHL_ACITAI_003). A. Lateral; B. Oblique. Scale bar: 1 mm (applies only to A). Note that the silver region is an entomological pin.
Diagnosis.
Body length 4.8 mm. This is the only species covered here with costate elytra (on the 2^nd^, 4^th^, and 6^th^ intervals). Also, is has erect, elongate scales along the odd-numbered elytral intervals (as in A. ryukyuana).
Distribution.
Acicnemis costulifera is recorded from Taiwan and Japan in Alonso-Zarazaga et al. (2023); however, Hubenthal (1919b) states that the holotype collection locality is unknown. We examined the holotype specimen and indeed, it does lack labels with any locality data. Furthermore, no specimens of this species collected from Japan were encountered in any of the examined collections. As such, we are confident that the record of this species from Japan is erroneous, and that the record from Taiwan should be treated with suspicion until additional specimens are found.
Acicnemis
cryptica
Taxon classificationAnimaliaColeopteraCurculionidae
Lewis & Kojima sp. nov.
183AA775-DE6B-51CB-8245-8DC267CF10C2
https://zoobank.org/D117E957-E5E4-47B0-BF8A-4444EC767456
Type material
(Japan: 72). Holotype. Japan: Nagano Prefecture: • Otari Onsen, Otari-mura, 23.VII.1990, M. Hasegawa, bears a red holotype label reading “HOLOTYPE / Acicnemis cryptica / Lewis & Kojima, 2025”, JHL_AREV_672, dissected male, deposited in KUM. Paratypes. Japan: Aomori Prefecture: • Towada, 14.VI.1957, K. Shimoyama (1 dissected male, KUM), JHL_AREV_498; Fukushima Prefecture: • Yunohana, Tateiwa V., Minami-Aizu, 14.VI.1947, “Y.K.” (1 dissected male, NMNS), JHL_AREV_139; Fukuoka Prefecture: • Hikosan (Buzen), 19.V.1939, H. Hori (1 dissected male, KUM), JHL_AREV_981; Gunma Prefecture: • Mt. Buson, 13–15.VII.1975, H. Irie (2, KUM), JHL_AREV_904 (dissected male), JHL_AREV_905; Kochi Prefecture: • Mt. Tebako, 7–10.VIII.1957, K. Morimoto (13, KUM), JHL_AREV_479 (dissected male), JHL_AREV_823 – JHL_AREV_827, JHL_AREV_829 – JHL_AREV_831, JHL_AREV_833 – JHL_AREV_835, JHL_AREV_837; • Mt. Tebako, 18–21.VII.1955, K. Kojima (2, KUM), JHL_AREV_828, JHL_AREV_832; • Monobe-mura, 26–28.VIII.1959, K. Morimoto (2, KUM), JHL_AREV_836, JHL_AREV_845; Ibaraki Prefecture: • Mt. Yamizo-san, Daigo-machi, 1.XII.1995, S. Nomura (1 dissected male, KUM), JHL_AREV_901; Iwate Prefecture: • Mt. Hayachine, 2–5.VIII.1982, H. Makihara (8, KUM), JHL_AREV_594 (dissected male), JHL_AREV_817, JHL_AREV_818, JHL_AREV_822, JHL_AREV_841 – JHL_AREV_844; Nagano Prefecture: • Otari Onsen, Otari-mura, 23.VII.1990, M. Hasegawa (1, TUA), JHL_AREV_819; Oita Prefecture: • Mt. Sobo, 29.VII.1953, T. Matsuda (1 dissected male, KUM), JHL_AREV_544; Shizuoka Prefecture: • Amagi-toge, Hacchoike Izu-hanto, 14.V.1980, J. Okuma (4, TUA), JHL_AREV_947 (dissected male), JHL_AREV_959; • Tokyo Metropolis: Nippara, 19.IX,1971, H. Takizawa, JHL_AREV_118 (1, HUM); • Ogawadani, Okutama, “1966.8.6”, T. Goh (1 dissected male, HUM), JHL_AREV_140; • Mt. Takao, 27.V.1917, E. Gallois (3, HUM) JHL_AREV_114 (dissected male), JHL_AREV_141, JHL_AREV_142; • Mt. Takao, 10.V.1914, E. Gallois (1, HUM), JHL_AREV_115; • Mt. Takao, Hachioji, 22.VIII.2024, 35.62972°N, 139.2525°E, R. Anderson, beating in mixed maple / beech / cypress forest, “2024-102” (2, CMNC), JHL_AREV_2035, JHL_AREV_2026; • Takao Musashi, V.1912, H. Takabayashi (1, HUM), JHL_AREV_116; Yamanashi Prefecture: • Daibosatsu, 19.V.1974, M. Tao (1, TUA), JHL_AREV_778; • Koganezawa-rindo, Ohtsuki-shi, 30–31.V.1980, J. Okuma (5, TUA), JHL_AREV_630 (dissected male), JHL_AREV_820, JHL_AREV_821; • Koganezawa-rindo, 23.VI.1974, S. Miyakawa (8, TUA), JHL_AREV_941 – JHL_AREV_945.
Acicnemis cryptica Lewis & Kojima, sp. nov. (JHL_AREV_778). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Diagnosis.
Body length 4.9–7.0 mm. Covered in dark to pale gray and black scales. Pronotum covered in black scales medially, but with white to yellow scales laterally (compare with A. albofasciata which also has white lateral pronotal bands, but bordered by black scales at the lateral-most extreme of the pronotum). Apical 1/2 of elytra with a V-shaped patch of white scales. Pronotum evenly punctured across dorsal surface, but often with shiny, impunctate longitudinal midline. Scutellum bare over anterior 1/2 of dorsal surface, otherwise covered in pale yellowish scales. Only odd elytral intervals with erect scales (even elytral intervals may possess erect scales at the extreme elytral apex). Sclerolepidia distinct and protruding. Third tarsomeres distinctly emarginate. Pedon with lateral edges approx. parallel in dorsal view, but sinuating into an acute point (Fig. 6C1–2). Internal sac without tuberculate or roughened musculature inside the aedeagus (compare with A. albofasciata, which has a tuberculate internal sac visible inside the aedeagus), but with hook-shaped protruding structure surrounded by tuberculate, roughened musculature.
Distribution.
This species is endemic to mainland Japan (excluding Hokkaido), north from Aomori Prefecture and south to Kyushu (Oita Prefecture, Fukuoka Prefecture).
Etymology.
The specific name cryptica is a reference to the remarkable similarity of this species in external morphology to A. albofasciata and also explains why it was overlooked for decades. We also suggest the Japanese common name ハイイロカレキゾウムシ [Haiiro-kareki-zoumushi], which translates in English to “Gray-dead-wood-weevil”.
Remarks.
This cryptic species was previously assumed to be a pale-color morph of A. albofasciata (see Morimoto and Miyakawa 1995: 50, fig. 11), and indeed only differs externally from A. albofasciata in overall color (pale gray in A. cryptica; black to dark gray in A. albofasciata) and weakly in scale pattern (entire lateral portion of pronotum white to yellow scaled in A. cryptica; isolated white to yellow band, bordered by black lateral pronotal band in A. albofasciata). The male genitalia of A. cryptica (which were apparently not examined previously), however, are completely distinct from A. albofasciata and differ in the shape of the aedeagus as well as in internal sac morphology (Fig. 6C1–2).
Acicnemis
dividicincta dividicincta
Taxon classificationAnimaliaColeopteraCurculionidae
Morimoto & Miyakawa, 1995
0B173776-B892-5A96-AF78-7090D19F92E6
Acicnemis yakushimana Morimoto & Miyakawa, 1995, syn. nov.
Type material examined
(Japan: 34). Holotype. Japan: Okayama Prefecture: • Mt. Gagyuzan, Takahashi City, 19–20.V.1975, H. Irie, bears red label reading “(HOLOTYPE) Acicnemisdividicincta Morimoto et. Miyakawa, 1994”, JHL_SYN_120, ELKU, ELKU 2957. Paratypes. Fukuoka Prefecture: • Mt. Hiko, 5.IX.1988, K. Morimoto (1, KUM), JHL_SYN_118; Kochi Prefecture: • Mt. Tebako, 7–10.VIII.1957, K. Morimoto (2, KUM), JHL_SYN_114, JHL_SYN_115; Nagasaki Prefecture: • Unzen, 18.VII.1946, M. Moritsu & Y. Kurosawa (3, KUM), JHL_SYN_102 (dissected male), JHL_SYN_113, JHL_SYN_119; Okayama Prefecture: • Mt. Gagyuzan, Takahashi City, 19–20.V.1975, H. Irie (3, KUM), JHL_SYN_111, JHL_SYN_112, JHL_DNA_142 (dissected male); Wakayama Prefecture: • Mt. Ootoh, 7.VIII.1980, I. Matoba (1, KUM), JHL_SYN_116.
Acicnemis dividicincta dividicincta Morimoto & Miyakawa, 1995 (JHL_AREV_772). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Other type material.
Kagoshima Prefecture: • Miyanoura, Yakushima Island, 26.VII.1974, T. Mikage, bears red label reading “(HOLOTYPE) Acicnemis yakushimana Morimoto et. Miyakawa, 1994”, JHL_SYN_180 (1, ELKU), A. yakushimana holotype specimen, ELKU 2959; • Miyanoura, Yakushima Island, 26–29.VII.1974, T. Mikage (13, KUM), JHL_SYN_122 – JHL_SYN_124, JHL_SYN_127 – JHL_SYN_130, JHL_SYN_132, JHL_SYN_133, JHL_SYN_138, JHL_SYN_139, JHL_DNA_141 (dissected male); • Shiratani, Yakushima Island, 24–26.VII.1974, T. Mikage (2, KUM), JHL_SYN_121, JHL_SYN_131; • Mt. Tachudake, Yakushima Island, 20.VII.1971, H. Irie (4, KUM), JHL_SYN_125, JHL_SYN_126, JHL_SYN_134, JHL_SYN_135; • Ambo – Kosugidani, Yakushima Island, 29.VII.1929, Hiroshi Hori (2, KUM), JHL_SYN_136, JHL_SYN_137; • Shiratani-rindo, Yakushima Island, 20.VII.1976, M. Kaneda (1, KUM), JHL_AREV_663.
Non-type material.
Ehime Prefecture: • Omogo, Iyu, 19.VIII.1969, T. Nakane, (1, HUM), JHL_AREV_772; • Tokyo Metropolis: Hachijo-jima, Mitsune, VI.2001, T. Kinoshita (1 dissected male, KPMNH), JHL_AREV_936; • Hachijo-jima, Noboryou-toge, 9.VI.1996, I. Hirai (1, KUM), JHL_AREV_938.
Diagnosis.
Body length 4.2–7.0 mm. Covered in black and dark gray scales, with two V-shaped, transverse white and black bands across middle of elytra. Pronotum with weakly patterned gray, black and faint yellow scales. Scutellum covered loosely in yellow scales (denuded in many specimens). Only odd elytral intervals with erect scales across elytral disk (some erect scales on 2^nd^ interval in apical fourth of elytra). Sclerolepidia distinct and protruding. Third tarsomeres distinctly emarginate. Pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a moderately acute point. Internal sac not visible inside aedeagus, but with a non-hooked protruding structure (unmodified in posterior 1/2, with roughened in anterior 1/2) that is clearly longer than pedon (Fig. 6D1–2).
Distribution.
This subspecies occurs in mainland Japan (central Japan south to Yakushima Island) and adjunctly on Hachijo Island (Tokyo Metropolis).
Remarks.
Acicnemis yakushimana was distinguished from A. dividicincta by Morimoto and Miyakawa (1995) on the basis of purported differences in the relative proportions of the funicular antennomeres and minor differences in color. Furthermore, A. yakushimana also reportedly possessed a sclerite in the internal sac at the gonopore. After dissecting several specimens of each species across their geographic range, we did not observe any differences in genital morphology between A. yakushimana and A. dividicincta. Furthermore, no observable differences in antennal funicle proportions were observed. As such, we hereby treat A. yakushimana syn. nov. as a junior subjective synonym of A. dividicincta.
Acicnemis
dividicincta okinawana
Taxon classificationAnimaliaColeopteraCurculionidae
Morimoto & Miyakawa, 1995
2923444F-AAB0-5781-AB11-91A0E92145B5
Type material examined
(Japan: 13). Holotype. Japan: Okinawa Prefecture: • Hiji, Kunigami-son, Okinawa Island, 8.VII.1974, T. Mikage, bears red label reading “(HOLOTYPE) Acicnemis dividicincta okinawana Morimoto et. Miyakawa, 1995”, JHL_SYN_110, ELKU, ELKU 2958. Paratypes. Japan: Kagoshima Prefecture: • Torigamine, Amami Island, 30.VI.1976, N. Morishimo (1, KUM), JHL_SYN_104; Okinawa Prefecture: • Hiji, Kunigami-son, Okinawa Island, 3–9.VII.1974, T. Mikage (5, KUM), JHL_SYN_101 (dissected male), JHL_SYN_103, JHL_SYN_105 – JHL_SYN_107.
Acicnemis dividicincta okinawana Morimoto & Miyakawa, 1995 (JHL_SYN_106). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Kagoshima Prefecture: • Mt. Yuwandake, Amami-Oshima, 5–9.VII.1968, K. Sakai (1, CMIC), CBM-ZI 68475; Okinawa Prefecture: • Tancha, 1–7.VI.2022, J.H. Lewis (1, OIST), JHL_DNA_126 (GenBank accession number: PV255626); • Tancha, Onna-son, Okinawa Island, 2.VI.2022, sweeping coastal vegetation, J.H. Lewis (1, OIST), JHL_SYN_109; • Yona, Okinawa Island, 19.X.1973, S. Azuma (1, RUMC), JHL_SYN_108; • Tancha, Onna-son, Okinawa Island, 29.VIII.2025, J.H. Lewis, to residential lights (1, OIST), JHL_AREV_2082; • Oku, Okinawa Island, 26.83604°N, 128.27191°E, 5–19.VIII.2016 (1, OIST), OKENT0089525 (GenBank accession number: PV255627).
Diagnosis.
Body length 4.8–6.5 mm. Identical to A. dividicincta dividicincta above, differing from that species only in possessing somewhat paler gray scales.
Distribution.
This subspecies occurs on Amami-Oshima (Kagoshima Prefecture) and Okinawa Island (Okinawa Prefecture).
Remarks.
We continue to recognize this subspecies as it is significantly isolated from the mainland populations and therefore presumably does represent a distinct intraspecific lineage.
Acicnemis
exilis
Taxon classificationAnimaliaColeopteraCurculionidae
Morimoto & Miyakawa, 1995
A9AD03D2-B69C-5CFD-A239-305F33D36AA1
Type material examined
(Japan: 21; Taiwan: 3; Total: 24). Holotype. Japan: Kagoshima Prefecture: • Amami-Oshima, Naze, 27.V.1978, T. Tsutsumi, bears red label reading “(HOLOTYPE) Acicnemis exilis Morimoto et. Miyakawa, 1994”, ELKU, ELKU 2963.
Acicnemis exilis Morimoto & Miyakawa, 1995 (OKENT0065625). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Kagoshima Prefecture: • Jinushi-daimyojin, Satomura, Nakano-jima Island, 7–9.VII.2019, S. Imada (2, KUM), JHL_AREV_595, JHL_AREV_596; Okinawa Prefecture: • Hikawa, Yonaguni Island, 3.V.2016, N. Tsuji (1, KUM), JHL_AREV_597; • Banna-koen, Ishigaki Island, 1.IX.2015, J. Aoki (9, KUM), JHL_SYN_145 (dissected male), JHL_AREV_598, JHL_AREV_599, JHL_AREV_601 – JHL_AREV_605; • JHL_AREV_724; • Mt. Omoto-dake, 15–17.IV.2014, S. Yamamoto (2, KUM), JHL_AREV_606, JHL_AREV_607; • Southeast Botanical Gardens, Okinawa Island, 10–24.II.2016 (1 dissected male, OIST), OKENT0109018; • Chatan, Ireibaru, 26.32468°N, 127.75918°E, 11–25.XI.2015 (1, OIST), OKENT0105900 (GenBank accession number: PV255629); • Nanjo, Tamagusuku, 26.32468°N, 127.75918°E, 9–23.IX.2015, OKENT0056923 (GenBank accession number: PV255628) (1, OIST); • Sefa-utaki, Nanjo, 15–29.VII.2016, (1, OIST), OKENT0065625; near Hiji, Okinawa Island, 16.III.2024, J.H. Lewis (1, OIST), JHL_AREV_998; • Banna-dake, Ishigaki Island, 20.VII.1973, H. Takizawa (1, HUM), JHL_ACITAI_086; Taiwan • Wulai (near Taipei), 27.V.1965, K. Morimoto (1 dissected male, KUM), JHL_AREV_624; • Kotosho Island, 13–15.VIII.1968, H. Makihara (2, KUM), JHL_AREV_757, JHL_AREV_758.
Diagnosis.
Body length 2.1–3.0 mm. Covered in pale brown, white, and gray scales, with two white-scaled spots in the apical 1/2 of the elytra. Scutellum covered in yellow scales. Only odd elytral intervals with erect scales. Sclerolepidia small but protruding. Third tarsomeres distinctly emarginate. Pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a rounded point. Internal sac without tuberculate or roughened musculature inside the aedeagus, but with a non-hooked protruding structure (unmodified in posterior 2/3, sclerotized in anterior 1/3) that reaches the tip of the temo (Fig. 6E1–2).
Distribution.
This small species occurs throughout the Ryukyu Islands (southern Kagoshima Prefecture and Okinawa Prefecture, Japan) as well as Taiwan.
Remarks.
This species appears to be closely allied with the similarly minute, long hind-legged Australian species Acicnemis meriones Pascoe, 1872, A. arachnopus Hubenthal, 1919, and the Sumatran species A. ambigua Hubenthal, 1919; however, phylogenetic analysis is required to confirm this. Preliminary observations suggest that some of the names in this group are synonyms.
Acicnemis
kiotoensis
Taxon classificationAnimaliaColeopteraCurculionidae
Nakane, 1963
CE8417E5-0CC2-5A60-A947-04F37FED721C
Type material examined
(Japan: 73; Taiwan: 4; Total: 77). Holotype. Japan: Kyoto Prefecture: • Kurama, 17.VI.1954, T. Nakane, bears red label reading “HOLOTYPE”, bears pink label reading “185-5”, Syst. Ent. Hokkaido Univ Japan SEHU 0000005125, HUM. Paratype. Japan: Kagoshima Prefecture: • Yakushima Island, Miyanoura, 28.IV.1954, Y. Kurosawa, bears red label reading “PARATYPE” (1, HUM).
Acicnemis kiotoensis Nakane, 1963 (JHL_AREV_775). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Kagoshima Prefecture: • Kirishima, 18.IX.2007, J. Aoki (2, KUM), JHL_AREV_161, JHL_AREV_162; • Mt. Kirishima, 22.VII.1956, J. Nagao, (1, KUM), JHL_AREV_775; • Amami Oshima, Hatsuno, 25.V.1960, T. Shibata (1, KUM), JHL_AREV_177; • Chuorindo, Amami Oshima, 27.VI.1980, A. Izumi (1, LBM), LBM 5117; • Chuorindo, Amami Oshima, 30.VI.1980, Yoshikawa (1, LBM), LBM 5119; • Hatsuno, Amami Oshima, 1–8.VII.1973, M. Ito (1, LBM), LBM 5130; • Yuwan, Amami Oshima, 7.VIII.1964, M. Nagai (1, HUM), JHL_AREV_645; • Amami Oshima, 21.VII.1955, T. Shirozu (1, KUM), JHL_AREV_660; • Ishihara-rindo, Amami Oshima, 9–11.VI.1978, R. Noda (1, KUM), JHL_AREV_661; • Nonigawa, Amami Oshima, 17.VI.1963, J. Nagao (1, KUM), JHL_AREV_662; • Hatsuno, Amami Oshima, 27.V.1960, T. Shibata (1, ZMH), ZMH 837205; • Yudomari-rindo, Yakushima Island, 26.VII.1982, A. Seki (2, LBM), LBM 5120, 5121; • Miyanoura, Yakushima Island, 17.VII.1968, H. Nomura (1, LBM), LBM 5124; • Miyanoura, Yakushima Island, 21–22.VII.1974, K. Emoto (2, LBM), LBM 5126, 5127; • Miyanoura, Yakushima Island, 15–26.VII.1972, H. Huzita (2, LBM), LBM 5129, 5131; • Miyanoura, Yakushima Island, 26–29.VII.1974, T. Mikage (5, KUM), JHL_AREV_648 – JHL_AREV_650; • Miyanoura, Yakushima Island, 29.VIII.1952, Takeya & Hirashima (1, KUM), JHL_AREV_653; • Miyanoura, Yakushima Island, 18.VI.1975, Y. Kurosawa (1, NMNS), JHL_AREV_679; • Okawa-rindo, Yakushima Island, 1–2.VIII.1974, T. Mikage (2, KUM), JHL_AREV_646, JHL_AREV_647; • Okawa, Yakushima Island, 22.VII.1975, H. Akiyama (1 dissected male, NMNS), JHL_AREV_157; • Shiratani, Yakushima Island, 26–30.VII.1979, M. Miura (8, HUM), JHL_AREV_634 – JHL_AREV_641; • Shiratani, Yakushima Island, 24–25.VII.1974, T. Mikage (5, KUM), JHL_AREV_657 – JHL_AREV_659; • Kosugidani, Yakushima Island, 19.VII.1974, T. Mikage (1, KUM), JHL_AREV_656; • Kosugidani, Yakushima Island, 23.VII.1950, T. Shirozu (1, KUM), JHL_AREV_655; • Kosugidani, Yakushima Island, 9.–10.VII.1952, Y. Kurosawa (4, NMNS), NSMT-I-C 10267–10269, NSMT-I-C 10301; • Hanayama Natural Forest, Yakushima Island, 21.VIII.1983, Sk. Yamane (3, HUM), JHL_AREV_642 – JHL_AREV_644; • Mt. Hoyoshi, Minamiosumi-cho, 24–25.III.2018, R. Ito (1, KUM), JHL_AREV_840; • Mt. Yuwan, Amami-Oshima, 30.VII.1963, L. Gressitt (1 dissected male, KUM), JHL_SYN_152; Kyoto Prefecture: • Mt. Kurama, 30.V.1982, K. Masaki (1, LBM), LBM 5118; • same locality, 8.X.1978, K. Masaki (1, LBM), LBM 5122; Miyazaki Prefecture: • Miyakonojo-shi, Natsu-cho, Miike, 22.VI.2019, R. Ito (1, KUM), JHL_AREV_175; • Mt. Aoidake, 7.VIII.1974, K. Morimoto (1 dissected male, KUM), JHL_DNA_147; Nagasaki Prefecture: • Mt. Taterasan, Izuhara, Tsushima Island, 24–27.VII.1985, A. Saito (15, CMIC), CBM – ZI 68476–68490; • Mt. Tatera, Tsushima Island, 18–20.V.1961, H. Kamiya (2, KUM), JHL_AREV_651, JHL_AREV_652; Nara Prefecture: • Kasugayama, 25.VII.1981, K. Masaki (1, LBM), LBM 5116; • Kasugayama, 10.V.1964, K. Kinugasa (1, LBM), LBM 5133; • Kasugayama, 20.V.1951, K. Sawada (1, KUM), JHL_AREV_654; • Mt. Kasuga, 9.V.1952, K. Sawada (1 dissected male, KUM), JHL_SYN_153; Okinawa Prefecture: • Okinawa Island, Yonahadake, 8–29.VI.1977, H. Irie (2, KUM), JHL_AREV_172, JHL_AREV_173; • Iriomote Island, Shirahama, 23–24.VI.1970, H. Makihara (2, KUM), JHL_AREV_158, JHL_AREV_159; • same locality, 31.VIII–5.IX.1969, H. Makihara (6, KUM), JHL_AREV_163 – JHL_AREV_168; • same locality, 26.VII.1963, Y. Miyatake (1, KUM), JHL_AREV_169; • Okinawa Island, Yona, 9–13.VIII.1969, H. Makihara (1, KUM), JHL_AREV_176; • Iriomote Island, Nakara River, 25–28.VI.1970, H. Makihara (2, KUM), JHL_AREV_170, JHL_AREV_171; • Iriomote Island, Taketomi-cho, Mt. Tedo, 2.X.2017, K. Narita (1, KUM), JHL_AREV_174; • Iriomote Island, 28.VI.1978, M. Kinjo (1, RUMC), JHL_AREV_524; • Iriomote Island, Funaura, 8.X.1977, M. Arasaki (1, RUMC), JHL_AREV_525. Taiwan • Mt. Hsin Kao, 1.VII.1961, S. Ueno, JHL_AREV_160 (1, KUM); • Meifeng, 15.V.1983, H. Townes (1, CMNC), JHL_AREV_667; • Anmashan, Taichung Hsien, 11–15.V.1992, A. Smetana (2, CMNC), JHL_AREV_707, JHL_AREV_708.
Diagnosis.
Body length 4.5–7.0 mm. Covered in pale and dark brown scales, with two obliquely shaped pale brownish, black-bordered spots across middle of elytra. Second funicular antennomere longer than funicular antennomeres 3 + 4 (Fig. 2C) (compare with A. koguma, which has the second funicular antennomere shorter than funicular antennomeres 3 + 4). Pronotum lacking impunctate longitudinal midline (present in A. koguma). Scutellum diagnostic: large, shiny, and lacking scales (Fig. 3B). Odd and 2^nd^ elytral intervals with erect scales (scales beginning at middle of elytra on 2^nd^ interval; from elytral base on odd intervals). Sclerolepidia indistinct, forming low, rounded, non-protruding tubercles (Fig. 4A). Third tarsomeres emarginate (not truncate as in A. koguma). Pedon with lateral edges approx. parallel in dorsal view but converging evenly into a wide, blunt point. Internal sac not visible inside the aedeagus, but with a non-hooked protruding structure (unmodified in posterior 1/2, with roughened in anterior 1/2) that is clearly longer than the pedon (Fig. 6F1–2).
Distribution.
This distinctive species occurs in Japan (mainland, throughout the Ryukyu Islands) and Taiwan.
Acicnemis
koguma
Taxon classificationAnimaliaColeopteraCurculionidae
Lewis & Kojima sp. nov.
6FA0DC04-D964-5807-AD2E-8B6E193B52DE
https://zoobank.org/78DAC1B8-51AE-4ABC-9E44-27A21760C117
Type material examined
(Japan: 3). Holotype. Japan: Wakayama Prefecture: • Mt. Otoh, 20.VII.1980, I. Matoba, bears a red holotype label reading “HOLOTYPE / Acicnemis koguma / Lewis & Kojima, 2025”, JHL_DNA_143, female, deposited in KUM. Paratypes. Japan • Tokyo Metropolis: Mt. Takao, 12.VIII.1969, H. Fujita, LBM accession # 5128 (1, LBM), JHL_SYN_141; • Mt. Takao, 3–4.VII.1965, K. Sakai (1, CMIC), CBM-ZI 68474.
Acicnemis koguma Lewis & Kojima, sp. nov. (JHL_SYN_141). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Diagnosis.
Body length 4.5–7.0 mm. Covered in pale and dark brown scales, with two obliquely shaped pale brownish, black-bordered spots across middle of elytra. Second funicular antennomere shorter than funicular antennomeres 3 + 4 (Fig. 2D) (longer in A. kiotoensis). Pronotum with impunctate longitudinal midline (absent in A. kiotoensis). Scutellum large (somewhat narrower than A. kiotoensis) and bare dorsally but bordered laterally by pale yellow scales (Fig. 3C). Only odd elytral intervals with erect scales (some erect scales on 2^nd^ interval in apical fifth of elytra) (A. kiotoensis with erect scales on the 2^nd^ elytral interval). Sclerolepidia indistinct, forming low, rounded, non-protruding tubercles. Third tarsomeres truncate (emarginate in A. kiotoensis).
Distribution.
This rare species is only known from two localities in mainland Japan, Mt. Takao (Tokyo Metropolis) and Mt. Otoh (Wakayama Prefecture).
Etymology.
The specific name koguma means little bear in Japanese, and is a reference to the apparent mountain-loving preference and small, brown nature of this rarely observed weevil species. We also suggest the Japanese common name コグマカレキゾウムシ [Koguma-kareki-zoumushi], which in English means “Little-bear-dead-tree-weevil”.
Remarks.
This cryptic species was clearly overlooked due to its remarkable similarity with A. kiotoensis in overall color and appearance, but differs consistently in the five characters listed in the diagnosis. The male remains unknown.
Acicnemis
laeta
Taxon classificationAnimaliaColeopteraCurculionidae
Hubenthal, 1919
BBBD59AA-596D-5FC7-A6EC-A6D76421B4F8
Type material examined
(Taiwan: 71). Lectotype (designated here). Taiwan • Fuhosho, “IX.07–09”, Sauter, SDEI, SDEI Muncheberg Col 20204, bears the original red label reading “Syntypus” as well as a new, red lectotype label reading “LECTOTYPE / Acicnemis laeta / Hubenthal, 1919 / Det. Lewis & Kojima, 2025”. Paralectotypes. Taiwan • Fuhosho, “IX.07–09”, Sauter (22, SDEI), SDEI Muncheberg Col 18617–18623 & 20199–20203 & 20205–20214, all bear a red label reading “Syntypus” and new, yellow paralectotype labels; • Hoozan, “08–10”, Sauter (18, SDEI), SDEI Muncheberg Col 18624–18632 & 20215–20223, all bear a red label reading “Syntypus” and new, yellow paralectotype labels; • Kosempo, Sauter (2, SDEI), SDEI Muncheberg Col 18633–18634, bear a red label reading “Syntypus” and new, yellow paralectotype labels.
Acicnemis laeta Hubenthal, 1919 (JHL_AREV_773). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Taiwan • Lingui, 18.VI.1979, S. Nakamura, W. Chen (1, KUM), JHL_AREV_526; • Pi Shan Spa, Tai Tung Hsein, 2.VI.1987, K. Baba (2, KUM), JHL_AREV_527, JHL_AREV_528; • Guandaoxi, I.VI.1973, K. Kojima (2, KUM), JHL_AREV_529, JHL_AREV_530; • Jihyueh-tan, Nantou Hsein, 12.V.1977, H. Fujita (1, KUM), JHL_AREV_531; • Kenting Park, Pingtung Hsein, 11.V.1970, M. Takagi (1, KUM), JHL_AREV_532; • Nanshanchi, Nantou Hsein, 7.V.1977, T. Niisato (1, KUM), JHL_AREV_533; • Liu Kui, 28.III.–10.IV.1986, K. Baba (2, KUM), JHL_AREV_534, JHL_AREV_535; • Kenting, Pingtung Hsein, 2.IV.1965, S. Ueno (2, KUM), JHL_AREV_536, JHL_AREV_537; • Nanshanchi, 30.IV.1965, T. Nakane, T. Shirozu (2, KUM), JHL_AREV_538, JHL_AREV_539; • Nanshanchi, Nantou Hsein, 12.VII.1966 (1, KUM), JHL_AREV_540; • Shyk Shan, near Liu Kui, 25.IV.1986, K. Baba (1 dissected male, KUM), JHL_AREV_541; • Nanshanchi, 3.V.1979, K. Emoto (1, KUM), JHL_AREV_664; • Lienhoachi, 12.V.1975, S. Imasaka (1, TUA), JHL_AREV_665; • “community park”, 28.X.1976 (1, TUA), JHL_AREV_666; • Nantou, 24.V.1983, K. Ra, (1, HUM), JHL_AREV_773; • Wushe, 29.IV.–3.V.1983, H. Townes (3, CMNC), JHL_AREV_669 – JHL_AREV_671; • Liukei, Kaohshiiung, 29.IV.–8.V.1982, H. Takizawa (3, HUM), JHL_AREV_675 – JHL_AREV_677; • near Liukuei, Kaohsiung Hsien, 5–9.IV.1995, H. Kojima (1, TUA), JHL_AREV_690; • “Formosa”, T. Kano (1, NMNS), JHL_AREV_680; • Kao; • Nanshan Riv. Trail, Ren’ai Township, Nantou County, 18–19.X.2017, K. Narita (2, KUM), JHL_DNA_124 (GenBank accession number: PV255630), JHL_AREV_125 (GenBank accession number: PV255631).
Diagnosis.
Body length 4.1–5.8 mm. Covered in pale brown scales, with alternating white and black scaled bands on odd elytral intervals. Scutellum covered in yellow to brown scales. Only odd elytral intervals with erect scales. Sclerolepidia distinct and protruding. Posterior edge of prosternum with prominent projections contiguous with the fore-coxae (Fig. 2B). Third tarsomeres distinctly emarginate. Pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a moderately acute point in apical 1/2. Internal sac protruding and with two bead-shaped sclerotized structures in basal 1/2, and a vase-shaped structure in apical 1/2 with roughened musculature (Fig. 6G1–2).
Distribution.
We only encountered this species from Taiwan. The records from Japan and the Kuril Islands (Russia / Japan) noted in Alonso-Zarazaga et al. (2023) are almost certainly erroneous and this species may have been confused with A. azumai. This species may plausibly occur in mainland Asia (China) (listed as occurring there by Alonso-Zarazaga et al. (2023)), but we have yet to examine any specimens of A. laeta from this region.
Remarks.
This species is superficially quite similar to A. azumai, but differs in pronotal, prosternal, and genital morphology.
Acicnemis
luteomaculata
Taxon classificationAnimaliaColeopteraCurculionidae
Morimoto & Miyakawa, 1995
DDDC7371-7A1C-5A5B-942A-07135233633F
Type material examined
(Japan: 45). Holotype. Japan: Fukuoka Prefecture: • Hiko-san, 13–14.VI.1957, K. Morimoto, bears red label reading “(HOLOTYPE) Acicnemis luteomaculata Morimoto et. Miyakawa, 1994”, ELKU, ELKU 2961. Paratypes. Japan: Ehime Prefecture: • Hojo-shi, Mt. Takanawa, 18.V.2003, T. Kurihara (1, KUM), JHL_AREV_420; Fukuoka Prefecture: • Hiko-san, 8.VI.–13.VII.1957, K. Morimoto (11, KUM), JHL_AREV_181 – JHL_AREV_190, JHL_ACITAI_090 (dissected male); • same locality, 4.VI.1958, K. Morimoto (2, KUM), JHL_AREV_192, JHL_AREV_193; • same locality, 10.VII.1938, Hori, Kawahara & Yasumatsu (1, KUM), JHL_AREV_194; • Mt. Wakasugi, 18.XII.1955, K. Oshima (1, KUM), JHL_AREV_120; • Wakasugi, 28.VII.1940, T. Shirozu (2, KUM), JHL_AREV_405, JHL_AREV_406; Fukushima Prefecture: • Mt. Shizukura-yama, Mishima Town, 37.3756°N, 139.6266°E, 11–20.VII.2018, K. Narita (1, KUM), JHL_AREV_412; Kochi Prefecture: • Mt. Tebako, 7–10.VIII.1957, K. Morimoto (2, KUM), JHL_AREV_196, JHL_AREV_197; Nara Prefecture: • Mt. Odaigahara, 2.VII.1978, H. Hiramatsu (1, KUM), JHL_AREV_198; • Kasugayama, 25.VII.1959, T. Hozumi (1, KUM), JHL_AREV_121; • Kasuga, 5–31.V.1959, K. Ueda (3, KUM), JHL_AREV_401 – JHL_AREV_403; • Kasuga, 30.IV.1961, K. Ueda (1, KUM), JHL_AREV_404; Okayama Prefecture: • Takahashi City, Mt. Gagyuzan, 19–20.V.1975, H. Irie (3, KUM), JHL_AREV_178 – JHL_AREV_180; Wakayama Prefecture: • Mt. Gomadan, 28.VII.1979, I. Matoba (1, KUM), JHL_AREV_191.
Acicnemis luteomaculata Morimoto & Miyakawa, 1995 (JHL_AREV_409). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Ehime Prefecture: • Hojo-shi, Mt. Takanawa, 18.V.2003, T. Kurihara (1, KUM), JHL_AREV_420; • Ishizuchi Mt., Omogo Valley, 18–25.VIII.1980, S. & J. Peck (1, CMNC), JHL_AREV_668; Fukuoka Prefecture: • Hiko-san, 23–26.VII.1968, M. Chujo (1, KUM), JHL_AREV_195; Hiroshima Prefecture: • Sandankyo, 12–13.V.1969, M. Chujo (1, KUM), JHL_AREV_418; Miyazaki Prefecture: • Miyakonojo-shi, Natsuo-cho, Miike, 22.VI.2019, R. Ito (6, KUM), JHL_AREV_407 – JHL_AREV_411, JHL_DNA_117 (GenBank accession number: PV255632); Nara Prefecture: • Kasuga, 10.V.1959, T. Shibata (1, KUM), JHL_ACITAI_091; Oita Prefecture: • Mt. Kuro-dake, 28.V.1990, S. Ogata (4, KUM), JHL_AREV_413 – JHL_AREV_416; • Tsukahara, Yufuin-cho, Yufu-shi, 21.XI.2016, R. Ito (1, KUM), JHL_AREV_721; • Tsukahara, Yufuin-cho, Yufu-shi, 27.V.2018, R. Ito (1, KUM), JHL_DNA_123 (GenBank accession number: PV255633); Wakayama Prefecture: • Komoridani, 21.VIII.1941, K. Sakaguchi (1, HUM), JHL_AREV_678; Yamagata Prefecture: • Tozawa-mura, Mt. Haguro, 23.VI.2003, J. Aoki (1, KUM), JHL_AREV_417.
Diagnosis.
Body length 4.0–5.4 mm. Covered in pale brown scales, with two obliquely shaped tan-colored, black-bordered spots across middle of elytra. Pronotum lacking impunctate longitudinal midline (present in A. koguma). Scutellum sparsely covered in pale yellow scales. All elytral intervals with erect scales. Sclerolepidia indistinct, forming low, rounded, non-protruding tubercles. Third tarsomeres distinctly truncate (Fig. 2E) (compare with the superficially similar A. kiotoensis, A. maculaalba, and A. sauteri, which all have emarginate third tarsomeres). Pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a blunt, rounded point. Internal sac not visible inside the aedeagus, but with a basal projection that bears a distinctive rectangular sclerite anteriorly at its apex (Fig. 6H1–2).
Distribution.
This species is found in mainland Japan (except Hokkaido) and has also been reported from South Korea (Park et al. 2009).
Acicnemis
maculaalba
Taxon classificationAnimaliaColeopteraCurculionidae
Roelofs, 1875
87491871-1E50-5A9B-AD08-F51AD234E18C
Type material examined
(Japan: 50). Holotype. Japan • unknown locality, “1910–320”, G. Lewis, bears a circular white, red-bordered label reading “Type H.T.”, NHMUK015024192.
Acicnemis maculaalba Roelofs, 1875 (JHL_AREV_777). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Fukuoka Prefecture: • Mt. Homan, 15.V.1975, H. Irie (1, KUM), JHL_AREV_583, misidentified A. luteomaculata paratype; Kagoshima Prefecture: • Sata (Osumi), Magome-Odomari, 28.V.1952, Esaki & Hirashima (1, KUM), JHL_AREV_556; • Kuchinoshima, 26.VI.–3.VII.1969, H. Makihara (3, KUM), JHL_AREV_557 – JHL_AREV_559; • Akuseki Island, 30.VII.1969, M. Sakai (4, KUM), JHL_AREV_560 – JHL_AREV_563; • Nakanoshima, 25.IV.–2.V.1975, H. Irie (4, KUM), JHL_AREV_564 – JHL_AREV_567; • Amami-Oshima, Hatsuno, 12.VI.1963, J. Nagao (1, KUM), JHL_AREV_570; • Yakushima Island, Miyanoura, 26.VII.1974, T. Mikage (1, KUM), JHL_AREV_571; • Funayuki, Yakushima Island, 26.VII.1989, M. Satoshi (1, CMIC), CBM ZI 159703; Kochi Prefecture: • Okinoshima (Tosa Province), 28.VI.–2.VIII.1953, K. Morimoto & K. Sugimoto (5, KUM), JHL_AREV_551 – JHL_AREV_555; Mie Prefecture: • Mie University Forest, Ichishi-gun, 11.VI.1956, M. Ichihashi (2, KUM), JHL_AREV_546, JHL_AREV_547; Miyazaki Prefecture: • Mt. Aoidake, 6.V.1976, K. Morimoto, Z. Kuranaga, A. Iwasaki, N. Yoshida (1, KUM), JHL_AREV_590; Okinawa Prefecture: • Komi, Iriomote Island, 14.III.1997, M. Chujo (1, KUM), JHL_SYN_143; • Iriomote Island, 28.V.1962, K. Kojima (1, KUM), JHL_AREV_545; • Yonahadake, 8–29.VI.1977, H. Irie (2, KUM), JHL_AREV_568, JHL_AREV_569; • Hiji, Okinawa Island, 3.VII.1974, T. Mikage (2, KUM), JHL_AREV_572, JHL_AREV_573; • Yona, Okinawa Island, 15.VI.1970, H. Makihara (1, KUM), JHL_SYN_142; • Yona, Okinawa Island, 29.VI.1976, H. Makihara (1, KUM), JHL_AREV_574; • Mt. ufudaki, Kohama Is., 29.V.2022, S. Imada, (1, KUM), JHL_AREV_777; • Mantabaru, Yonaguni Island, 25–27.V.2022, S. Imada (3, KUM), JHL_AREV_575 – JHL_AREV_577; • Omoto-dake, Ishigaki Island, 30.VI.1974, T. Mikage (1, KUM), JHL_AREV_578; • Mt. Banna, Ishigaki Island, 23–26.V.1990, K. Morimoto (1, KUM), JHL_AREV_579; • Shirahama, Iriomote Island, 31.VIII.–5.IX.1969, H. Makihara (3, KUM), JHL_AREV_580 – JHL_AREV_582; • Funaura, Iriomote Island, 1.X.1978, S. Azuma (1, RUMC), JHL_AREV_713; • Nakagusuku Park, Okinawa Island, 26.28557°N, 127.79491°E, 3–17.II.2016, OKENT0068361 (GenBank accession number: PV255635) (1, OIST); • Yona Field, Okinawa Island, 26.73974°N, 128.23598°E, 8–22.VII.2016, OKENT0050489 (GenBank accession number: PV255634) (1, OIST); • Toyko District: Oji-ike, Miyake Island, 5–7.IV.2010, J. Aoki (6, KUM), JHL_AREV_584 – JHL_AREV_589; • Sato, Mt. Kurosa – Kitakao-san, Mikura-jima Island, 2.VI.1979, J. Okuma (1 dissected male, KUM), JHL_AREV_426; Wakayama Prefecture: • near Mt. Ohtoh, 13.VI.–7.VIII.1980, I. Matoba (2, KUM), JHL_AREV_548, JHL_AREV_549; • Mt. Ohtoh, 16–19.VII.1982, H. Makihara (1, KUM), JHL_AREV_500.
Diagnosis.
Body length 2.6–5.0 mm. Covered in pale and dark brown scales, with two obliquely shaped pale brownish, black-bordered spots across middle of elytra. Pronotum lacking impunctate longitudinal midline (present in A. koguma). Scutellum small, bordered by pale, yellow scales, and weakly bare dorsally or covered in scales (Fig. 3D) (compare with A. kiotoensis which possesses a large, broad, bare scutellum). Odd, 2^nd^, and 4^th^ elytral intervals with erect scales. Sclerolepidia indistinct, forming low, rounded, non-protruding tubercles. Third tarsomeres emarginate (Fig. 2G) (as opposed to truncate, as in A. koguma). Pedon with lateral edges approx. parallel in dorsal view but converging in distal 1/2 evenly into a blunt point. Pedon stout (pedon length / total aedeagus length including temo = 0.2). Internal sac protruding and with roughened musculature across length; with two bent, tube-like sclerotized structures anteriorly at its apex (Fig. 6I1–2).
Distribution.
This species occurs broadly throughout Japan (mainland Japan and Ryukyu Islands) and Taiwan and is also recorded from China by Alonso-Zarazaga et al. (2023).
Remarks.
Acicnemis maculaalba forms a species group with A. sauteri. The association of A. maculaalba and A. sauteri is supported by morphology and weakly (BS: 38) by our CO1 gene tree. See also A. cordata Remarks section.
Acicnemis
nobilis
Taxon classificationAnimaliaColeopteraCurculionidae
Hubenthal, 1919
3A98A048-6003-545B-A301-8AA6A9CF6822
Type material examined
(Taiwan: 10). Holotype. Taiwan • Hoozan, “H. Sauter 1910”, bears small yellow label reading “1911 14”, bears red label reading “Typus”, SNSD.
Acicnemis nobilis Hubenthal, 1919 (JHL_AREV_496). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Taiwan • Penpuchi, Nantou Hs., 13.VII.1966, H. Sasaji (2, KUM), JHL_SYN_151 (dissected male), JHL_AREV_492; • Sangkang, Nantou Hs., 14.VII.1966, H. Sasaji (3, KUM), JHL_SYN_150 (dissected male), JHL_AREV_491, JHL_AREV_493; • Lushan, Nantou Hsein, 8.VI.1976, H. Makihara (1, KUM), JHL_AREV_494; • Nanshanchi, Wenchuan Hs., 27.IV.–4.V.1977, T. Mikage (1, KUM), JHL_AREV_495; • Hsi-Leng, 3.V.1981, S. Tsuyuki (1, TUA), JHL_AREV_496; • Kaohsiung, 5.VIII.1978, T. Niisato (1, TUA), JHL_AREV_497.
Diagnosis.
Body length 4.1–5.2 mm. Covered in dark gray and black scales, with a pale-yellow scale, V-shaped, transverse band across middle of elytra. Basal patch of black scales surrounding scutellum approx. V-shaped and extending prominently onto the second elytral intervals (as opposed to the similar A. nohirai which has an elongate I-shaped patch of black scales on the first elytral interval that barely extends on the second elytral interval basally). Pronotum lacking impunctate longitudinal midline (present in A. koguma). Scutellum covered in pale scales. Elytra with erect scales only on odd intervals. Sclerolepidia distinct and protruding. Third tarsomeres distinctly emarginate. Pedon with lateral edges approx. parallel in dorsal view, but sinuating into a blunt, rounded point. Internal sac protruding from aedeagus, with roughened musculature posteriorly and a club-shaped sclerotized structure anteriorly at its apex (Fig. 6J1–2).
Distribution.
This species is currently only known from Taiwan.
Acicnemis
nohirai
Taxon classificationAnimaliaColeopteraCurculionidae
Morimoto & Miyakawa, 1995
99B40C3B-45FA-56D2-8318-1925B0964925
Type material examined
(Japan: 23). Holotype. Japan: Gifu Prefecture: • Hirayu, 19.VI.1963, T. Nohirai, bears a red label reading “(Holotype) Acicnemis nohirai Morimoto et Miyakawa, 1994”, ELKU, ELKU 2956. Paratypes. Japan: Aomori Prefecture: • Kawauchi, Shimokita-gun, 25.VII.1984, S. Yamauchi (1, KUM), JHL_AREV_435; Ehime Prefecture: • Omogokei, 15.VI.1981, S. Naomi (1, KUM), JHL_AREV_437; Gifu Prefecture: • Hirayu, 19.VI.1963, T. Nohirai (5, KUM), JHL_AREV_429 – JHL_AREV_433; • Hokkaido: Horoka, Kamishihoro-cho, 17.VII.1976, H. Irie (1, KUM), JHL_AREV_434; • Uchida, 18.V.1930 (1, HUM), JHL_AREV_454; • Sapporo, “3.VIII.80” (1, KUM), JHL_AREV_460; Nagano Prefecture: • Tobira Spa, Matsumoto, 7.VIII.1973, T. Mikage (1 dissected male, KUM), JHL_SYN_147; Niigata Prefecture: • Kurokawa, 30.VII.1962, K. Baba (1 dissected male, KUM), JHL_SYN_146; Oita Prefecture: • Kurasame, Kuju, 15.VI.1975, H. Irie (2, KUM), JHL_AREV_438, JHL_AREV_439; Tochigi Prefecture: • Chuzenji-Yumoto, Shimotsuke, 22.VII.1923, T. Esaki (1, KUM), JHL_AREV_436; Yamanashi Prefecture: • Fuji-rindo, Mt. Fuji, 10–12.VIII.1976, H. Irie (1, KUM), JHL_AREV_440; • Mt. Kaikoma, 13.VIII.1962, K. Oshima (1, KUM), JHL_AREV_441; • Masutomi, Nirasaki, 16.VII.2006, H. Takizawa (1, HUM), JHL_AREV_461.
Acicnemis nohirai Morimoto & Miyakawa, 1995 (JHL_AREV_774). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Gifu Prefecture: • Hirayu, 19.VI.1963, T. Nohirai (1, KUM), JHL_AREV_442; Nagano Prefecture: • Otaki-mura, 31.V.2010, K. Takahashi (1, KUM), JHL_DNA_119 (PV255636); Oita Prefecture: • Yuhu-shi, Yuhuin-chou, Tsukahara, 18.IX.2016, R. Ito (1, KUM), JHL_DNA_122 (PV255637); Yamanashi Prefecture: • Fuji-rindo, Mt. Fuji, 3.X.1992, T. Mikage (1, KUM), JHL_AREV_443; • Mt. Daibosatsu, Hagiwara, 12.VIII.2005, T. Kurihara (1, KUM), JHL_AREV_720; • Masutomi, 24–25.V.1968, H. Takizawa, (1, HUM), JHL_AREV_774.
Diagnosis.
Body length 4.9–6.0 mm. Covered in dark gray and black scales, with a white-scaled, V-shaped, transverse band across middle of elytra. Elytra with elongate I-shaped patch of black scales on the first elytral interval (surrounding the scutellum) that barely extends onto the second elytral interval. Pronotum lacking impunctate longitudinal midline (present in A. koguma). Scutellum covered in dark brown scales. Elytra with erect scales only on odd intervals. Sclerolepidia distinct and protruding. Third tarsomeres distinctly emarginate. Pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a blunt, rounded point. Internal sac not visible inside aedeagus, but with protruding basal structure that bears a prong-shaped sclerite anteriorly at its apex (Fig. 6K1–2).
Distribution.
This species is apparently endemic to mainland Japan and occurs from Hokkaido south to Kyushu (Oita Prefecture).
Acicnemis
palliata
Taxon classificationAnimaliaColeopteraCurculionidae
Pascoe, 1872
2D859A73-57DE-559B-94B2-4FCD616645CF
Acicnemis dorsonigrita Voss, 1941, syn. nov.
Type material examined
(China: 2; Japan: 20: Total: 22). Holotype. Japan: Nagasaki Prefecture: • Nagasaki, Pascoe Coll. 93–60, bears a circular white, red-bordered label reading “Type H.T.”, NHMUK015026632. Paratypes. China • Tienmuschan, bears a red label reading “Type” (1 dissected male, NHMB), Acicnemis dorsonigrita type specimen, JHL_ACITAI_010; • Tienmuschan, bears a red label reading “Cotypus Acicnemis dorsonigrita” (1, ZMH), ZMH 841827.
Acicnemis palliata Pascoe, 1872 (JHL_AREV_779). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Chiba Prefecture: • Ichihara-shi, Kongouuchi, 28.V.1989, M. Satoshi (2, CMIC), CBM-ZI 159709, CBM-ZI 159710; Ehime Prefecture: • Higashino, Matsuyama-shi, 1–22.VII.2016, Y. Hisasue (2, KUM), JHL_AREV_484, JHL_AREV_485; Fukuoka Prefecture: • Chikuzen, Kashii, Kusaba Kosen, 5.VI.1935, Esaki, Hori, Ho, Fujino, Yasumatsu, Nomura & Kawahara (1, KUM), JHL_ACITAI_046; • Mt. Homan, 2.VII.1978, R. Noda (1 dissected male, KUM), JHL_ACITAI_007; • Nokonosima, 8–29.V.2005, H. Hirano (2, KUM), JHL_AREV_486, JHL_AREV_487; Fukushima Prefecture: • 1.VII.2000, K. Takahashi (1, KUM), JHL_AREV_483; • Kaneyama-machi, Okuriyama, 2.VI.2012, J. Aoki (3, KUM), JHL_AREV_488; • Aizu, Kaneyama, 20–21.VII.2012, J. Aoki (4, KUM), JHL_AREV_489; Kagawa Prefecture: • Takamatsu-shi, 15.V.1961, M. Chujo (1, KUM), JHL_ACITAI_051; • Miki-cho, 16.VI.1961, M.Chujo (1, KUM), JHL_ACITAI_050; • Kinuki-shi, 25.V.2015, J. Aoki (1, KUM), JHL_AREV_490; • Kita-gun, Hirai-cho, 13.V.1949, K. Iwata (1, ZMH), ZMH 837201; Nagasaki Prefecture: • Tsushima, Izuhara, 15.V.1971, H. Irie (1, KUM), JHL_ACITAI_006; Nara Prefecture: • Kasuga, 4.VI.1961, K. Ueda (1, KUM), JHL_ACITAI_049; Oita Prefecture: • Tsukahara, Yufuin-machi, Yufu-shi, 18.VI.2016, R. Ito, (1, KUM), JHL_AREV_779; Osaka Prefecture: • Mt. Iwawaki, 18.VI.1966, H. Kimura (1, KUM), JHL_ACITAI_048.
Diagnosis.
Body length 6.1–7.0 mm. Covered in pale brown, dark gray, and black scales, with a tan-colored, V-shaped, transverse band across middle of elytra. Pronotum lacking impunctate longitudinal midline (present in A. koguma). Scutellum bordered by pale brown scales, but conspicuously bare and shiny dorsally (Fig. 3E). Elytra with erect scales only on odd intervals. Sclerolepidia distinct and protruding (Fig. 4B). Third tarsomeres distinctly emarginate. Pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a blunt, rounded point. Internal sac not visible inside aedeagus, but with a protruding basal structure that bears two hook-like structures anteriorly at the apex that are curved 90 degrees inward (Fig. 6L1–2).
Distribution.
This species occurs in Japan (mainland) and China, and has also been reported from South Korea (Park et al. 2009) and the Kuril Islands (Alonso-Zarazaga et al. 2023).
Remarks.
An examination of the holotype of A. palliata (NHMUK), the holotype of A. dorsonigrita (NHMB), a paratype of A. dorsonigrita (ZMH), as well as comparisons of the original descriptions of these two species revealed that A. dorsonigrita syn. nov. is a junior synonym of A. palliata. In particular, the holotypes of A. palliata and A. dorsonigrita (1) both have scutella which are bare dorsally, bordered by scales around the perimeter (2) are more than 5 mm in length (measured from the apex of the elytra to the base of the rostrum), and (3) bear the same characteristic pattern of scales dorsally. Furthermore, the original descriptions of these species certainly appear to refer to the same species, and notably the body length measurements given for A. palliata (“3 lin.” = 6.25 mm) and A. dorsonigrita (5–5.8 mm) are approximately equal. The much smaller species (at most 4.4 mm) previously referred to as A. dorsonigrita by Morimoto and Miyakawa (1995) is new and is described below as A. squamata.
Acicnemis
postica
Taxon classificationAnimaliaColeopteraCurculionidae
Hubenthal, 1919
C5483005-1F16-5B59-A4CF-C62B2EA1A456
Acicnemis cruciata Kleine, 1924, syn. nov.
Type material examined
(Japan: 31; Taiwan: 5; Total: 36). Lectotype (designated here): Taiwan • “Formosa Is.”, “Coll. Pape”, SDEI Müncheberg Col – 18570 (1, SDEI), bears red label reading “Syntypus” and a new, red lectotype label reading “LECTOTYPE / Acicnemis postica / Hubenthal, 1919 / Det. Lewis & Kojima, 2025”. Paralectotypes. Taiwan • Fuhosho, “IX-7–09”, H. Sauter, SDEI Müncheberg Col – 18571 (1, SDEI), bears red label reading “Syntypus” and a new, yellow paralectotype label; • Kankau (Koshun), VI. 1912, H. Sauter, SDEI Müncheberg Col – 18572 (1, SDEI), bears red label reading “Syntypus” and a new, yellow paralectotype label.
Acicnemis postica Hubenthal, 1919 (JHL_AREV_780). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Other type material.
• Kosempo, “VII–19”, H. Sauter, Acicnemis cruciata holotype (1, SDEI), SDEIColeoptera #303422, SDEI Müncheberg Col – 18568.
Non-type material.
Taiwan • Taoyuan Hsein, Paling, 24.V.1977, H. Fujita (1, KUM), JHL_ACITAI_057; • Wenchuan Hsein, Lushan, 4.VII.1978, T. Mikage (1, KUM), JHL_ACITAI_058; • Ping Tung Hsein, Kenting, 2.IV.1965, S. Ueno (1, KUM), JHL_ACITAI_060; • Kenting, 31.III.1970, Y. Miyake (2 dissected males, KUM), JHL_ACITAI_082, JHL_SYN_179; • Chihpen Wenchuan, Taitung Hsien, 7. VI.1980, H. Makihara (1, KUM), JHL_AREV_735; • Jiuyuehtan, 31.V.1978, K. Kusama (1, KUM), JHL_AREV_736; • Taidong, Lijia road, 22.8062°N, 121.0310°E, 19.VIII.2013, V. Grebennikov (1, CNCI), CNCCOLVG00006901 (BOLD accession number: MEBLA191-14); Japan: Kagoshima Prefecture: • Shinokawa, Setouchi-cho, Amami Oshima, 10.VII.2008, K. Ozaki (1, CMIC), CBM ZI 136615; • Nakanoshima Island, 1–2.V.1975, H. Irie (1, KUM), JHL_AREV_750; • Nagata, Yakushima Island, 17.V.1960, J. Nagao (1, KUM), JHL_AREV_754; • Mt. Kindake, Nakano Is., 7–9.VI.2019, S. Imada, (1, KUM), JHL_AREV_780; Okinawa Prefecture: • Ishigaki Island, Mt. Omoto, 18–22.VI.1991, K. Morimoto (1, KUM), JHL_ACITAI_055; • Mt. Omoto, 21.V.1977, H. Irie (1, KUM), JHL_AREV_755; • Mt. Omoto, 18–21.IV.1975, H. Irie (1 dissected male, KUM), JHL_ACITAI_062; • Ushikumori, Iriomote Island, 7.X.1963, K. Morimoto (1, KUM), JHL_AREV_756; • Yonaguni Island, Urabu-dake, 25–26.VI.1974, T. Mikage (6, KUM), JHL_ACITAI_056, JHL_AREV_751 – JHL_AREV_753; • Okinawa Island, Hiji, 9.VII.1974, T. Mikage (2, KUM), JHL_ACITAI_059; • Fusato, Okinawa Island, 5.VI.1970 (1, KUM), JHL_AREV_762; • Omotodake, Ishigaki Island, 8.VII.1968, M. Takagi (1, KUM), JHL_AREV_761; • Funaura, Iriomote Island, 2.X.1978 (2, RUMC), JHL_AREV_709, JHL_AREV_710; • Banna, Ishigaki Island, 14.III.1964, S. Azuma (1, RUMC), JHL_AREV_712; • Haneji, Okinawa Island, 10.VIII.1979, T. Kohama (1, RUMC), JHL_AREV_711; • Mt. Inbi-take, Yonaguni Island, 2.VI.2021, S. Imada (1, KUM), JHL_AREV_723; • Kuburadake, Yonaguni Island, 15.VI.2001, L. Hirano (1, KUM), JHL_AREV_725; • Shirahama, Iriomote Island, 31.VIII–5.IX.1969, H. Makihara (9, KUM), JHL_AREV_726 – JHL_AREV_734; • Shirahama, Iriomote Island, 4.X.1963, S. Ueno (1, KUM), JHL_AREV_737; • upper Nakara, Iriomote Island, 12.III.1964, Y. Miyatake (1, KUM), JHL_AREV_738; • Komi, Iriomote Island, 12.VII.1963, Y. Miyatake (1, KUM), JHL_AREV_739; • Sonai, Yonaguni Island, 25–29.VIII.1969, H. Makihara (10, KUM), JHL_AREV_740 – JHL_AREV_749; • Okinawa Island (1, OIST), JHL_AREV_983 (GenBank accession number: PV255638); • Tancha, Okinawa Island, 18.V.2024, J.H. Lewis, sweeping vegetation (1, OIST), JHL_AREV_994.
Diagnosis.
Body length 3.2–5.2 mm. Covered in orange, white, dark and pale gray scales. Elytra with inner five or six intervals black and gray; outer intervals with pale brown to orange scales; with V-shaped band of white scales along middle three or four intervals in center of elytra. Pronotum lacking impunctate longitudinal midline (present in A. koguma). Scutellum small and covered in erect, hair-like yellow scales. Elytra with erect scales only on odd intervals. Sclerolepidia distinct and protruding. Third tarsomeres emarginate. Metatibia evenly curved, not sinuate as in most other species. Pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a blunt, rounded point. Internal sac not visible inside aedeagus, but protruding and with two sclerotized cylinder-shaped structures posteriorly near the base (Fig. 6M1–2).
Distribution.
This species occurs commonly throughout southern Japan (Ryukyu Islands) and Taiwan.
Remarks.
Examination of the A. cruciata Kleine, 1924 syn. nov. holotype (SDEIColeoptera #303422) revealed that this is a junior synonym of A. postica as it possesses all of the key characteristics that define this distinctive species (same scale pattern, evenly curved metatibiae, small, scaled scutellum). We designate one of the Hubenthal A. postica syntypes (SDEI Müncheberg Col – 18570) as a lectotype to fix the identity of this species.
Acicnemis
ryukyuana
Taxon classificationAnimaliaColeopteraCurculionidae
Lewis, 2023
9B388526-9B1E-5202-BB6E-58F07231B225
Figs 1, 2F, 6N1–6, 2, 25, 32A, C
Type material examined
(Japan: 28). Holotype. Japan: Okinawa Prefecture: • Ishigaki, Omoto-dake, 2.VII.1991, K. Hidetada, bears black-bordered red label reading “HOLOTYPE / Acicnemis ryukyuana / Lewis, 2023”, JHL_ACI_004, dissected male, deposited in KUM. Paratypes. Japan: Okinawa Prefecture: • Ishigaki Island, Yarabu-dake, 28.V.1998, K. Takahashi (1, KUM), JHL_ACI_001; • Ishigaki Island, Omoto-dake, 2.VII.1991, K. Hidetada (9, KUM), JHL_ACI_002, JHL_ACI_003, JHL_ACI_005–JHL_ACI_011; • Ishigaki Island, Mt. Yarabudake, 17–20.IV.1998, S. Ohmomo (1, KUM), JHL_ACI_012; • Ishigaki Island, Omotodake, 30.VI.1974 (1, KUM), JHL_ACI_013; • Okinawa Island, Kunigami, Yambaru National Park, Yona Field (26.73972°N, 128.23630°E), 4–18. IX.2015, A. Miyagi, Y. Tamaki, I. Maehira, L. Iha, S. Iriyama, T. Yoshida (2, OIST), OKENT0063788, OKENT0063789 (dissected male); • Okinawa Island, Kunigami, Yambaru National Park, Yona Field (26.73972°N, 128.23630°E), 7–21.VIII.2015, M. Yoshimura, M. Ogasawara (1, OIST), OKENT0053190 (GenBank accession number: PV255639); • Okinawa Island, Kunigami, Yambaru National Park, Yona Field (26.73894°N, 128.23720°E), 21.VIII–4. IX.2015, Y. Tamaki, T. Yoshida (2, OIST), OKENT0055037, OKENT0053264; • Okinawa Island, Kunigami, Yambaru National Park, Yona Field (26.73972°N, 128.23630°E), 18.IX–16.X.2015, A. Miyagi, I. Maehira, T. Yoshida (1, OIST), OKENT 0053498 (GenBank accession number: PV255640); • Okinawa Island, Hiji, 8.VII.1974, T. Mikage (1, KUM), JHL_ACI_014.
Acicnemis ryukyuana Lewis, 2023 (OKENT0063788). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Okinawa Prefecture: • Takeda-rindo, Ishigaki Island, 19–22.II.2007, H. Makihara & E. Kagaya (1, KUM), JHL_AREV_625.
Diagnosis.
Body length 3.2–5.0 mm. Covered in dark gray, black, and bright yellow scales, with a black-bordered, yellow spot at the center of each elytron. Apex of elytra covered in yellow scales, and contrasting with overall dark dorsal surface of elytra. Pronotum lacking impunctate longitudinal midline (present in A. koguma), but with an M-shaped band of yellow scales. Scutellum small, red, shiny, and largely bare over dorsal surface. Elytra with long, erect scales only on odd intervals. Sclerolepidia distinct and protruding. Third tarsomeres weakly emarginate to truncate (Fig. 2F). Pedon with lateral edges approx. parallel in dorsal view, but converging evenly into a blunt, rounded point. Internal sac not visible inside aedeagus, but with protruding basal structure that bears a distinctive U-shaped formation anteriorly at its apex (Fig. 6N1–2).
Distribution.
This species is known only from Okinawa Prefecture, Japan (Yambaru National Park (Okinawa Island) and forested regions of Ishigaki Island.
Remarks.
This species is very distinct from other Japanese Acicnemis and its range-restricted, endemic presence in pristine, subtropical rain forest dominated parts of Okinawa Prefecture was hitherto a mystery. However, preliminary morphological and molecular evidence suggest a sister relationship between A. ryukyuana and an undescribed Southeast Asian (Thai) species that possesses emarginate third tarsomeres. Acicnemis ryukyuana likely represents a lineage that was isolated from the undescribed Thai species lineage when the Ryukyu Islands separated from mainland Asia (see Osozawa et al. 2011) and evolved truncate third tarsomeres as derived character.
Acicnemis
sauteri
Taxon classificationAnimaliaColeopteraCurculionidae
Hubenthal, 1919
5D5F21B1-AD66-5E6F-8067-88A6CA815813
Type material examined
(Japan: 21; Taiwan: 18; Total: 39). Lectotype (designated here). Taiwan • Hoozan, 1910, H. Sauter, bears a red label reading “Typus” and a new, red lectotype label reading “LECTOTYPE / Acicnemis sauteri / Hubenthal, 1919 / Det. Lewis & Kojima, 2025”, SNSD, JHL_ACITAI_004. Paralectotypes. Taiwan • Hoozan, “08–10”, Sauter (3, SDEI), bear a red label reading “Syntypus”, SDEI Muncheberg Col 18573, 18574, 18583; • Kosempo, Sauter (4, SDEI), bear a red label reading “Syntypus”, SDEI Muncheberg Col 18575, 18576, 18577, 18578; • Fuhosho, “VIII-09”, Sauter (3, SDEI), bear a red label reading “Syntypus”, SDEI Muncheberg Col 18580, 18581, 18582; • Taihorinsho, “X-09”, Sauter (1, SDEI), bears a red label reading “Syntypus”, SDEI Muncheberg Col 18579.
Acicnemis sauteri Hubenthal, 1919 (JHL_AREV_623). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Taiwan • Lushan Wenchuan, Nantou Hsien, 6–7.VI.1976, H. Makihara (2, KUM), JHL_AREV_609, JHL_AREV_611; • Sun Moon Lake, Nantou Hsien, 6.XII.2013, J. Aoki (10, KUM), JHL_SYN_148 (dissected male), JHL_SYN_149 (dissected male), JHL_AREV_613 – JHL_AREV_620; • Wukongshan, Liu-kuei, Kaohsiung Hsein, 2.V.1996, H. Takizawa (1, HUM), JHL_ACITAI_085; Japan: Kagoshima Prefecture: • Nakanoshima Island, 25–30.IV.1975, H. Irie (1, KUM), JHL_AREV_608; • Jinushi-daimyojin, Satomura, Nakano-jima Island, 7–9.VII.2019, S. Imada (1, KUM), JHL_AREV_610; • Sato, Nakano-jima Island, 7–9.VII.2019, N. Tsuji (3, KUM), JHL_AREV_691 – JHL_AREV_693; • Aden, Kikai-jima Island, 6.V.2022, R. Ito (3, KUM), JHL_AREV_623, JHL_AREV_908, JHL_AREV_909; • Kikai-jima Island, 4.III.2012, J. Aoki (1, KUM), JHL_AREV_695; • Torigamine, Amami-Oshima, 3.VIII.1976, N. Morishima (1, LBM), LBM 5123; • Hatsuno, Amami-Oshima, 24–30.VI.1973, M. Ito (1 dissected male, LBM), LBM 5132; • Sumiyo, Amami Oshima, 27.V.2004, K. Takahashi (1, KUM), JHL_AREV_694; • Akuseki-jima Island, Ue-shuraku, Sakamori-jinja, 2–6.VII.2019, S. Imada (1, KUM), JHL_DNA_121 (GenBank accession number: PV255642); • Nakano-jima Island, Satomura, Jinushi-daimyojin, 7–9.VII.2019, S. Imada (1, KUM), JHL_DNA_120 (GenBank accession number: PV255641).
Diagnosis.
Body length 4.9–6.2 mm. Covered in pale and dark brown scales, with two obliquely shaped white, black-bordered spots across middle of elytra. Pronotum lacking impunctate longitudinal midline (present in A. koguma). Scutellum small, bordered by pale yellow scales, and weakly bare antero-dorsally (compare with A. kiotoensis which possesses a large, broad, bare scutellum). Odd and 2^nd^ elytral intervals with erect scales (4^th^ elytral interval also occasionally with 1–2 scales near white spot). Sclerolepidia indistinct, forming low, rounded, non-protruding tubercles. Third tarsomeres emarginate (as opposed to truncate, as in A. koguma). Pedon with lateral edges approx. parallel in dorsal view, but converging in distal 1/2 evenly into a blunt point. Pedon elongate (pedon length / total aedeagus length including temo = 0.26). Internal sac protruding and unmodified in the posterior 1/2, but with roughened musculature in anterior 1/2 (Fig. 6O1–2).
Distribution.
This species occurs in southern Japan (Kagoshima Prefecture) and Taiwan, and has also been recorded from South Korea (Alonso-Zarazaga et al. 2023). The apparent lack of this species in the southern Ryukyu Islands (Okinawa Prefecture) is odd; however, similar patterns are known in other weevils (e.g., Aphanerostethus magnus Lewis & Kojima, 2024)
Remarks.
This species is closely allied with A. maculaalba (see Remarks for that species). We designate one of Hubenthal’s syntypes (SNSD, JHL_ACITAI_004) as a lectotype to fix the identity of this species.
Acicnemis
shibatai
Taxon classificationAnimaliaColeopteraCurculionidae
Voss, 1971
F6287459-457D-5125-A8A1-2D3A16A2CFE4
Type material examined
(Japan: 17; South Korea: 1; Taiwan: 2; Total: 20). Holotype. Japan: Kagoshima Prefecture: • Ikari, Amami-Oshima, 28.V.1960, T. Shibata, KUM, bears red label that reads “Holotype”. Paratypes. Japan: Kagoshima Prefecture: • Ikari, Amami-Oshima, 6.V.–4.VI.1960, T. Shibata, JHL_AREV_444 – JHL_AREV_451, JHL_ACITAI_064 (9, KUM), bear red label that reads “Paratype”.
Acicnemis shibatai Voss, 1971 (JHL_AREV_776). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Chiba Prefecture: • Mt. Kiyosumi, 9.VI.1963, K. Morimoto (1, KUM), JHL_AREV_459; • Mt. Kiyosumi, 14.VI.1992, T. Ito (1, CMIC), CBM ZI 63020; Kagoshima Prefecture: • Yuwan, Amami-Oshima, 31.VII.1963, Y. Hirashima (1, KUM), JHL_AREV_457; • Mt. Yuwan, Amami-Oshima, 30.VII.1963, L. Gressitt (1 dissected male, KUM), JHL_ACITAI_070; • Ishihara-rindo, 9–11.VI.1978, R. Noda (1, KUM), JHL_AREV_458; • Mikyo, Tokunoshima, 27.VII.1963, Y. Hirashima (1, KUM), JHL_AREV_453; • Nishinakama, Amami-Oshima, 2.VI.1970, H. Makihara, (1, KUM), JHL_AREV_776; Okinawa Prefecture: • Shirahama, Iriomote Island, 31.VIII–5.IX.1969, H. Makihara (1, KUM), JHL_AREV_452; South Korea • Kwangnung, Pochon-gun, Kyonggi-do, 16–19.VII.1992, K. Morimoto (1, KUM), JHL_AREV_419; Taiwan • Wulai, 27.V.1965, K. Morimoto (2, KUM), JHL_AREV_455, JHL_AREV_456.
Diagnosis.
Body length 3.0–3.8 mm. Covered in dark and pale brown, white, and black scales, with a white-scaled, V-shaped band in the center of the elytron. Pronotum lacking distinct longitudinal, impunctate midline (present in A. koguma). Scutellum covered in pale scales. Elytra with erect scales only on odd intervals. Sclerolepidia distinct and protruding. Third tarsomeres emarginate. Pedon tube-shaped with lateral edges approx. parallel in dorsal view, but sinuating evenly into a blunt, rounded point. Internal sac with a small sclerotized structure posteriorly, and two prong-like structures anteriorly at its apex (Fig. 6P1–2).
Distribution.
This species occurs in Japan (mainland, Ryukyu Islands south to Amami-Oshima), South Korea, and Taiwan.
Acicnemis
shigematsui
Taxon classificationAnimaliaColeopteraCurculionidae
Morimoto & Miyakawa, 1995
B87BD522-0EA3-561B-A8C4-3A55BE18EDB3
Type material examined
(Japan: 9). Holotype. Japan: Okinawa Prefecture: • Ishigaki Island, Omoto-dake, 1.VIII.1989, K. Shigematsu, bears red label reading “(HOLOTYPE) Acicnemis shigematsui Morimoto et. Miyakawa, 1994”, JHL_ACITAI_052, ELKU, ELKU 2960. Paratypes. Okinawa Prefecture: • Iriomote Island, Shirahama, 31.VIII–5.IX.1969, H. Makihara (2, KUM), JHL_ACITAI_005 (dissected male), JHL_ACITAI_047; • Iriomote Island, Mt. Ushiku, 4.XI.1963, G. A. Samuelson (1, KUM), JHL_ACITAI_053; • Iriomote Island, Upper Nakara River, 12.III.1964, Y. Miyatake (1, KUM), JHL_ACITAI_045.
Acicnemis shigematsui Morimoto & Miyakawa, 1995 (JHL_ACITAI_052). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Non-type material.
Japan: Okinawa Prefecture: • Ishigaki Island, Yarabu-dake, 3.V.1998, K. Takahashi (1, KUM), JHL_ACITAI_054; • Omoto-dake, 1–4.VII.2009, R. Ito (1, KUM), JHL_AREV_481; • Omoto-dake, 29.III.2022, T. Nozaki (1, KUM), JHL_AREV_482; • Okinawa Island, Naha, Shuri, “Sumiyoshi Park” (probably referring to Sueyoshi Park), 23.XI.1990, K. Morimoto (1, KUM), JHL_ACITAI_044.
Diagnosis.
Body length 5.1–7.0 mm. Covered in pale brown, dark gray, and black scales, with a tan-colored, V-shaped, transverse band across middle of elytra. Pronotum lacking distinct longitudinal, impunctate midline (present in A. koguma). Scutellum covered in pale scales (Fig. 3F). Elytra with erect scales only on odd intervals. Sclerolepidia distinct and protruding. Third tarsomeres distinctly emarginate. Pedon with lateral edges approx. parallel in dorsal view, but weakly sinuating into a blunt, subtriangular point. Internal sac not visible inside aedeagus, but with a protruding basal structure extending to the end of the temo and tuberculate (roughened) musculature anteriorly at the apex (Fig. 6Q1–2).
Distribution.
This species is endemic to Okinawa Prefecture, Japan, and is currently only known from Ishigaki, Iriomote, and apparently also Okinawa Island.
Remarks.
This species is quite similar in size and color to A. palliata; however, it was differentiated by Morimoto and Miyakawa (1995) on the basis of elytral color differences alone. To verify the validity of this species the type series of A. shigematsui and A. palliata were compared, and several significant differences in external and genital morphology were discovered which are all newly reported in the dichotomous key below. Furthermore, although A. shigematsui was previously only known from Ishigaki and Iriomote Islands, a specimen deposited in the KUM was collected on Okinawa Island (400 km away from the type localities). No specimens of A. shigematsui have since been collected on the island. Further field studies are required to ascertain whether this rarely collected species is truly established on the island.
Acicnemis
Taxon classificationAnimaliaColeopteraCurculionidae
sp. 1
F14219CE-481D-553D-900D-14BA7FAE6024
Material examined
(Myanmar: 2; Taiwan: 7; Thailand: 3; Vietnam: 2; Total: 14). Myanmar • Tamu Village, Kachin State, 7.VII.2003, A. Abe (2, KUM), JHL_SYN_169 (dissected male), JHL_AREV_717; Taiwan • “Formosa”, T. Kano (1, NMNS), JHL_AREV_681; • His-Leng, 5.V.1981, S. Tsuyuki (5, TUA), JHL_SYN_175 (dissected male), JHL_SYN_176 (dissected male), JHL_AREV_714 – JHL_AREV_716; • Lushan Wenchuan, Nantou Hsien, 6.VI.1976, H. Makihara (1, KUM), JHL_AREV_719; Thailand • Pupin-Meo, 11.X.1993, T. Senoh (1 dissected male, KUM), JHL_SYN_170; • Chiang Rai, Wiang Pa Pao, 12.XI.2012, K. Takahashi (2, KUM), JHL_AREV_718 (EGP0160-H9), JHL_AREV_722; • Doi Suthep, 23.VI.1993, T. Senoh, (1, KUM), JHL_AREV_766; Vietnam • Phia Oac National Park near Phia Den, Cao Bang Province, 22°32'20"N, 105°51'57"E, 19–22.III.2012, S.D. Gaimari, M. Hauser, H.T. Pham (2, CMNC), JHL_AREV_703, JHL_AREV_704; • “IIea” Binh, Tonkin, bears red label reading “simpliciata Typus H.”, bears brown label reading “A. simpliciata m ♂ Heller det. 1930” (1, SNSD), JHL_ACITAI_093.
Acicnemis sp. 1 (JHL_AREV_766). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Diagnosis.
Body length 4.1–5.0 mm. Covered in pale and dark brown scales, with bright X-shaped band of white scales across apical 1/2 of elytra, which extends up to the elytra humeri. Distinctive bright white scale bands also present on hind femora. Pronotum lacking distinct longitudinal, impunctate midline (present in A. koguma). Scutellum small, bordered by pale, yellow scales, and weakly bare dorsally (or covered in scales); compare with A. kiotoensis which possesses a large, broad, bare scutellum. Only odd elytral intervals with broad, erect scales. Sclerolepidia numerous, dense, and nub-like, protruding laterally outward (unlike A. maculaalba, A. kiotoensis, and A. koguma which have indistinct, low, rounded sclerolepidia). Third tarsomeres emarginate (as opposed to truncate, as in A. koguma). Pedon with lateral edges approx. parallel in dorsal view, but sinuating in distal 1/2 into a distinct point. Internal sac protruding and clear across posterior 2/3; with two bean-shaped and U-shaped sclerotized structure anteriorly at apex (Fig. 6R1–2).
Distribution.
This widely distributed species occurs in Vietnam, Thailand, Myanmar, and Taiwan.
Remarks.
One of K.M. Heller’s specimen of this species in the SNSD collection has a holotype label with the name “A. simpliciata” written (JHL_ACITAI_093); however, we were unable to find a formal description of this species in any of Heller’s works. Heller often attached type labels to specimens without formally publishing the descriptions (Olaf Jäger – SNSD Collection Manager to JHL, pers. comm. 2025; Katsura Morimoto – KUM Curator to HK pers. comm., 2025). Thus, this species is likely undescribed; however, there are a number of other similar Southeast Asian species for which type material should be compared with Acicnemis sp. 1 before formal description. This falls outside of the scope of the current work (study area); however, we still acknowledge the presence of this species in Taiwan and treat it here as an unnamed species.
Acicnemis
squamata
Taxon classificationAnimaliaColeopteraCurculionidae
Lewis & Kojima sp. nov.
DE46B977-D138-5A19-A1AF-1AE7EC20886C
https://zoobank.org/E5C566CF-28A3-4D1B-918D-4FE0D42B0B50
Type material
(Japan: 46). Holotype. Japan: Tokushima Prefecture: • Mt. Tsurugi, Awa, 2.VI.1957, S. Ueno, JHL_ACITAI_068, dissected male, deposited in KUM. Paratypes. Japan: Aomori Prefecture: • Oyama-nagane, Nakadomari-machi, Kitatsugaru-gun, 7.VI.2010, K. Morimoto (1, KUM), JHL_AREV_702; Fukui Prefecture: • Kinome-toge, 25.VI.1982, K. Morimoto (1, KUM), JHL_ACITAI_065; Fukushima Prefecture: • Yunohana, south Aizu, 8.VIII.1949, K. Nagayama (1, HUM), JHL_AREV_474; • Yunohana, south Aizu, 7.VI.1981, Y. Kurosawa (1, NMNS), JHL_AREV_682; • Yunohana, south Aizu, 20.VII.1974, Y. Kurosawa (2, NMNS), JHL_AREV_683; • Masuzawa Valley, Tateiwa-mura, 6.VI.2005, K. Morimoto (1, KUM), JHL_AREV_698; • Odayama Mt., Monden V., Kita-Aizu, 22.VI.1948, Y. Kurosawa (1 dissected male, NMNS), JHL_AREV_107; Gifu Prefecture: • unknown locality, 24.V.1949 (1, NHMB), JHL_ACITAI_077; • Hinaga, 22.VII.1946 (1, NHMB), JHL_ACITAI_078; • Hida, 1.VI.1957, T. Nakane (1, HUM), JHL_AREV_472; • Kawai-Idani, Hida, 1–2.VI.1957, T. Nakane (3, HUM), JHL_AREV_476 – JHL_AREV_478; Gunma Prefecture: • Osawa, Katashina, 5.VI.1988, T. Shinbori (1, KPMNH), JHL_AREV_684; • Numata, 30.V.1953, T. Takei (1, ZMH), ZMH 837203; • Hokkaido: Sapporo, “Takano” & H. Kono (2, HUM), JHL_AREV_462, JHL_AREV_465; • Rausu Shiretoko, 8.VII.1958, T. Nakane (2, HUM), JHL_AREV_475; • Chitose, 1.VIII.2011, J. Aoki (1, KUM), JHL_AREV_699; Ibaraki Prefecture: • Mt. Tsukuba, 11.VIII.1989, M.J. Sharkey (2, CMNC), JHL_AREV_3000, JHL_AREV_3001; Ishikawa Prefecture: • Yamanaka, Kaga, 20.VII.1910, T. Otsuka (1, HUM), JHL_AREV_463; Iwate Prefecture: • Shizukuishi, 7.VI.2010, Aoki Junichi (1, KUM), JHL_ACITAI_072; Kanagawa Prefecture: • Kuno, Odawara, 8.VI.2014, K. Watanabe (3, OIST), JHL_AREV_687 – JHL_AREV_689; • Mt. Ono, Yamakita, 10.V.1988, N.Niwa, (1, KUM), JHL_AREV_771; • Kochi Prefecture: • Engyoji, 22.VII.1952, Y. Wada (1, KUM), JHL_ACITAI_066; • Kumamoto Prefecture: • Mt. Hakucho, 14.V.1983, M. Chujo (1, KUM), JHL_ACITAI_063; • Kyoto Prefecture: • Miyama, Kyoto University Research Forest, 31.V.2010, Junichi Aoki (1, KUM), JHL_ACITAI_069 (GenBank accession number: PV255643); Nagano Prefecture: • Kaida Highlands, Kiso, 5.IX.2009, J. Aoki (1, KUM), JHL_AREV_696; • “Noziri Hotel”, 29.V.1941, T. Nakane (1, HUM), JHL_AREV_471; Nara Prefecture: • Mt. Omine, 23.V.1974, T. Hattori (1, KUM), JHL_AREV_467; Niigata Prefecture: • Mt. Yakemine, 3.VII.1949, H. Koike (1, KUM), JHL_ACITAI_061; • Kurokawa, 28.V.1955, K. Baba (1, ZMH), ZMH 837202; Okayama Prefecture: • Tsuki, Soja, 18.IV.1976, T. Aono (1, HUM), JHL_AREV_469; Osaka Prefecture: • Minoh, 30.V.1940, K. Taniguchi (1, HUM), JHL_AREV_470; Shizuoka Prefecture: • Kakegawa, Ijiri, 3.V.2010, J. Aoki (1, KUM), JHL_ACITAI_073; • Kawanehon-machi, 28.VIII.2009, J. Aoki (1, KUM), JHL_AREV_697; Tokushima Prefecture: • Mt. Tsurugi, 27.VII.1961, M.T. Chujo (1, KUM), JHL_ACITAI_067; Tochigi Prefecture: • Nikko-shi, Chugushi, Ryuzuno-taki Fall, 19.VI.2014, S. Yamamoto (1, KUM), JHL_ACITAI_076 (GenBank accession number: PV255644); • Ashio, Funeishi-toge, 2.VII.2006, K. Takahashi (1, KUM), JHL_AREV_701; • Tokyo Metropolis: Niijima Island, Mt. Mukoyama, 15.V.1979, H. Fujita (1, KUM), JHL_ACITAI_071; • Kogezawa, “1954.5.10” (1 dissected male, KUM), JHL_ACITAI_009; • Tokura, 6.VIII.1950, S. Ueno (2, HUM), JHL_AREV_468; • Mt. Takao, 14.IV.1985, A. Tanaka (1 dissected male, HUM), JHL_AREV_106; Wakayama Prefecture: • Mt. Kohya, VII.1949, M. Hayashi (1, HUM), JHL_AREV_473; Yamagata Prefecture: • Yonezawa, 22.VI.1948, Y. Kurosawa (1, HUM), JHL_AREV_464; • Naderayama, 23.V.1943, Y. Kurosawa (1, NMNS), NSMT-I-C 07303; • Seki, near Yonezawa, 2.VII.1944, Y. Kurosawa (1, NMNS), NSMT-I-C 07304; Yamanashi Prefecture: • Masutomi, 13–14.VI.1970, H. Takizawa (1, HUM), JHL_AREV_466; • Masutomi, “10.VI.2603” (1, NMNS), NSMT-I-C 07305; • Akeno, 6.VI.2010, T. Shinbori (2, KPMNH), JHL_AREV_685, JHL_AREV_686; • Kami-yoshida, Fuji-yoshida city, 15.VII.2024, J.H. Lewis (1, OIST), JHL_AREV_986.
Acicnemis squamata Lewis & Kojima, sp. nov. (JHL_AREV_771). A. Lateral; B. Dorsal. Scale bar: 1 mm.
Diagnosis.
Body length 3.0–4.4 mm. Covered in pale yellow, white, gray, and black scales. Pronotum with central longitudinal band of dark gray to black scales, bordered laterally by pale brown scales. Elytra with first intervals alternating black (at base of scutellum) and yellow; second to sixth intervals gray in posterior 1/2 of elytra, humeri with yellow to pale scales; apical 1/2 of elytra with transverse white V-shaped band extending from first to fifth intervals; apex of elytra largely with pale to yellow scales. Pronotum lacking impunctate longitudinal midline (present in A. koguma). Scutellum covered in bright yellow to dull gray scales. Only odd elytral intervals with erect scales. Sclerolepidia numerous, dense, and nub-like, protruding laterally outward. Third tarsomeres emarginate. Apex of pedon with distinct sinuation and pointed (Fig. 6S1–2). Internal sac of aedeagus distinctly scaly in appearance, protruding, and not extending to midpoint of temo; anterior apex of internal sac ending in two scaly feather-like tips (Fig. 6S1–2).
Distribution.
This species is only known from mainland Japan, from Hokkaido south to Kyushu (Kumamoto Prefecture).
Etymology.
The specific name squamata is a reference to the diagnostic internal sac of the aedeagus which bears scaly patterning similar to snakeskin.
Remarks.
See A. palliata Remarks section for taxonomic history. Acicnemis squamata differs from A. palliata (= A. dorsonigrita) in size (A. squamata is 3.0–4.4 mm in length whereas A. palliata is 6.1–7.0 mm in size), genitalia (A. squamata with internal sac of aedeagus scaly in appearance, protruding, and ending in two scaly featherlike tips; A. palliata without scaly internal sac, and with two protruding hook-like structures curved inwards at 90°) and scutellum scaling (A. squamata with scutellum covered in scales; A. palliata with scutellum prominently bare along the dorsal surface). See also the dichotomous key below.
Acicnemis
suturalis
Taxon classificationAnimaliaColeopteraCurculionidae
Roelofs, 1875
CDF7E382-0987-565C-A937-4527DCFB4285
Type material examined
(Japan: 24). Holotype. Japan: Hyogo Prefecture: • Kobe, “1910–320”, G. Lewis, NHMUK015026634, bears a circular, white and red label reading “Type H.T.”.
Non-type material.
Japan: Aomori Prefecture: • Nei, Misawa-shi, 8.VI.2010, K. Morimoto (1, KUM), JHL_AREV_507; Chiba Prefecture: • Nagara, Chosei-gun, 12.VI.1988, M. Satoshi (2, CMIC), CBM ZI 159815, 159816; Gunma Prefecture: • Minakami, 6–7.VI.2008, K. Morimoto (6, KUM), JHL_AREV_504, JHL_AREV_505; Ehime Prefecture: • Mt. Takanawa, Matsuyama-shi, 11–12.VIII.2015, K. Kuroda (1, KUM), JHL_AREV_506; Fukuoka Prefecture: • Mt. Wakasugi (Chikuzen), 23.V.1954, K. Morimoto, JHL_AREV_422, JHL_AREV_423, JHL_ACITAI_092 (dissected male) (3, KUM); • Mt. Hiko, 15.VII.1955, H. Kamiya, JHL_AREV_424; Hyogo Prefecture: • Kobe, Mt. Maya, 3.IX.1963, K. Morimoto, JHL_AREV_421 (1, KUM); Kanagawa Prefecture: • Mimasutoge, 6.V.1990, H. Takizawa (1, HUM), JHL_AREV_632; • Hatano, 7.VI.1983, H. Takizawa (1, HUM), JHL_AREV_633; Kyoto Prefecture: • Katsura-gawa, 12.VI.1918, M. Suzuki, JHL_AREV_425 (1, KUM); • Kyoto University Ashu Forest, Miyama-cho, Tanann City, 23.V.2009, K. Morimoto (1, KUM), JHL_AREV_503; Tochigi Prefecture: • Nanma, Kanuma, 4.V.1993, H. Takizawa (1, HUM), JHL_AREV_631; Niigata Prefecture: • Kamikawa-cho, 3.VI.2000, K. Takahashi (2, KUM), JHL_AREV_508.
Acicnemis suturalis Roelofs, 1875 (JHL_AREV_421). A. Lateral; B. Dorsal. Scale bar: 1 mm.
X-ray μCT generated 3D models of two Acicnemis species. The colored parts in the segmented models below represent hindwing (in blue), metanotum + dorso-longitudinal flight muscles (in green), and dorso-ventral flight muscles (in orange). A. A. ryukyuana Lewis, 2023 (OKENT0053498) dorsal view with scales removed, showing a relatively smooth pronotum and distantly spaced punctures; B. A. albofasciata (Ter-Minasian, 1953) (JHL_AREV_782) dorsal view with scales removed, showing a moderately rugose to roughened pronotum with interspaces between punctures much smaller; C. A. ryukyuana segmented model with the right elytron removed, showing a comparively larger metanotum and dorso-longitudinal flight muscles relative to the elytra (~ 30%); D. A. albofasciata showing a comparatively smaller metanotum and dorso-longitudinal flight muscles relative to the elytra (~ 23%).
Diagnosis.
Body length 4.0–4.7 mm. Covered in pale brown, dark and pale gray, pale yellow, and black scales, with two small white spots on the elytra (one on each side) in the apical 1/2 of the elytra. Pronotum lacking impunctate longitudinal carina (present in A. koguma). Scutellum covered in pale scales. Elytra with erect scales only on odd intervals. Sclerolepidia distinct and protruding. Third tarsomeres distinctly emarginate. Pedon with lateral edges converging in dorsal view, sinuating into an elongate, tapered tip. Internal sac protruding from aedeagus, and with two cylinder-shaped sclerotized structures anteriorly at the apex (Fig. 6T1–2).
Distribution.
This distinctive species occurs in Japan (mainland), Taiwan, and has also been reported from South Korea and the Kuril Islands (Russia) (Park et al. 2009; Alonso-Zarazaga et al. 2023).
Key to the Acicnemis of East Asia
**: **
Discussion
The island country of Japan has experienced multiple, intermittent periods of land bridge connection with mainland Asia since the Miocene (25-15 MYA) (Tojo et al. 2017; Matsuzaki et al. 2022), and this complex geologic history is partially what accounts for the diverse faunal composition of the region. Another major factor accounting for the faunal diversity of Japan is the latitudinal breadth of the country, which extends northward from Hokkaido (45.52°N) south to the Ryukyu Islands (Hateruma-jima Island, 24.04°N) (Tojo et al. 2017). Although most of Japan falls within the Palearctic Region, the southern Ryukyu Islands lie on the Palearctic-Oriental Regions border (Komaki 2021), and Japan has seen influxes and mixing of fauna from both regions (Morimoto et al. 2006). The faunal composition of weevils (and presumably other phytophagous beetles) in Japan is particularly complicated by the recent discovery that weevil eggs can survive consumption by birds, pass through the gut, be deposited on neighboring islands, and develop without complication into mature adults (Lin et al. 2021). Furthermore, inter-island dispersal of some species has likely been facilitated by transport in fruits and seeds that are blown to sea (Yeh et al. 2018).
Herein, we reviewed the East Asian Acicnemis and uncovered three new species in mainland Japan. Although Acicnemis is most thoroughly studied in Japan, our study revealed that overlooked diversity stills exists even in this region. Two of our newly described species, A. cryptica (mainland Japan, excluding Hokkaido) and A. koguma (central mainland Japan), are cryptic and sister to more widespread, previously described species (A. albofasciata: Russia and mainland Japan (including Hokkaido); and A. kiotoensis: mainland Japan, Ryukyu Islands, Taiwan). The existence of overlooked Acicnemis diversity in a relatively well-studied region highlights the importance of extensive genital dissection across the entire geographic range of what otherwise may be assumed to be single species. In addition to dissection, we used DNA barcoding and X-ray μCT to successfully delineate species pairs. As dissection is not always an option when working with rare, fragile, or type material, these alternative non-destructive techniques can both potentially be incorporated in the delineation process as a complement to traditional morphological methods. Although DNA barcoding successfully delineated all species pairs treated in this work, X-ray μCT for sub-scale cuticular comparison was only informative when working with sufficiently distantly related species (e.g., A. albofasciata and A. ryukyuana). Although X-ray μCT has been successfully used to delineate weevils within the same genus by pronotal and elytral punctation (see Lewis et al. 2024), clearly the utility of X-ray μCT in species delineation and cryptic morphology discovery is highly dependent on the target group, a factor that should be considered when evaluating the suitability of the method in future works.
Supplementary Material
XML Treatment for Acicnemis
XML Treatment for Acicnemis albofasciata
XML Treatment for Acicnemis azumai
XML Treatment for Acicnemis biarcuata
XML Treatment for Acicnemis bickhardti
XML Treatment for Acicnemis cordata
XML Treatment for Acicnemis costulifera
XML Treatment for Acicnemis cryptica
XML Treatment for Acicnemis dividicincta dividicincta
XML Treatment for Acicnemis dividicincta okinawana
XML Treatment for Acicnemis exilis
XML Treatment for Acicnemis kiotoensis
XML Treatment for Acicnemis koguma
XML Treatment for Acicnemis laeta
XML Treatment for Acicnemis luteomaculata
XML Treatment for Acicnemis maculaalba
XML Treatment for Acicnemis nobilis
XML Treatment for Acicnemis nohirai
XML Treatment for Acicnemis palliata
XML Treatment for Acicnemis postica
XML Treatment for Acicnemis ryukyuana
XML Treatment for Acicnemis sauteri
XML Treatment for Acicnemis shibatai
XML Treatment for Acicnemis shigematsui
XML Treatment for Acicnemis
XML Treatment for Acicnemis squamata
XML Treatment for Acicnemis suturalis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alonso-Zarazaga MA Lyal CHC (1999) A world catalogue of families and genera of Curculionoidea (Insecta: Coleoptera) (excepting Scolytidae and Platypodidae). Entomopraxis, S.C.P., Barcelona, Spain, 315 pp.
- 2Alonso-Zarazaga MA Barrios H Borovec R Bouchard P Caldara R Colonnelli EGúltekin L HlaváčP Korotyaev B Lyal CHC Machado A Meregalli M Pierotti H Ren LSánchez-Ruiz M Sforzi A Silfverberg H Skuhrovec J Trýzna M Velázquez de Castro AJ Yunakov NN (2023) Cooperative Catalougue of Palearctic Coleoptera Curculionoidea. Monografías Electrónicas SEA 8. Sociedad Entomológica Aragonesa S.E.A, Zaragoza, 780 pp.
- 3Edgar RC (2004) MUSCLE: Multiple sequence alignment with high accuracy and high throughput.Nucleic Acids Research 32(5): 1792–1797. 10.1093/nar/gkh 34015034147 PMC 390337 · doi ↗ · pubmed ↗
- 4Fairmaire L (1849) Essai sur les Coléoptères de la Polynésie (Suite). Revue et Magasin de Zoologie Pure et Appliquée (2) 1 (10): 504–516.
- 5Hubenthal W (1919 a) Beiträge zur Kenntnis der Curculioniden-Gattung Acicnemis Lacordaire (Col.).Archiv für Naturgeschichte 83(8): 92–160.
- 6Hubenthal W (1919 b) Beiträge zur Kenntnis der Curculioniden-Gattung Acicnemis Lacordaire (Col.).Archiv für Naturgeschichte 83(9): 53–155.
- 7Kalyaanamoorthy S Minh BQ Wong TK Fvon Haeseler A Jermiin LS (2017) Model Finder: Fast model selection for accurate phylogenetic estimates.Nature Methods 14(6): 587–589. 10.1038/nmeth.428528481363 PMC 5453245 · doi ↗ · pubmed ↗
- 8Kleine R (1924) Neue Arten der Gattung Acicnemis Lacordaire nebst faunistischen Mitteilungen.Archiv für Naturgeschichte 90(4): 148–164.