Solanum stellaticalycinum, a new simple-leaved species of the Pteroidea clade from Peru (Potato Clade, Solanum, Solanaceae)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1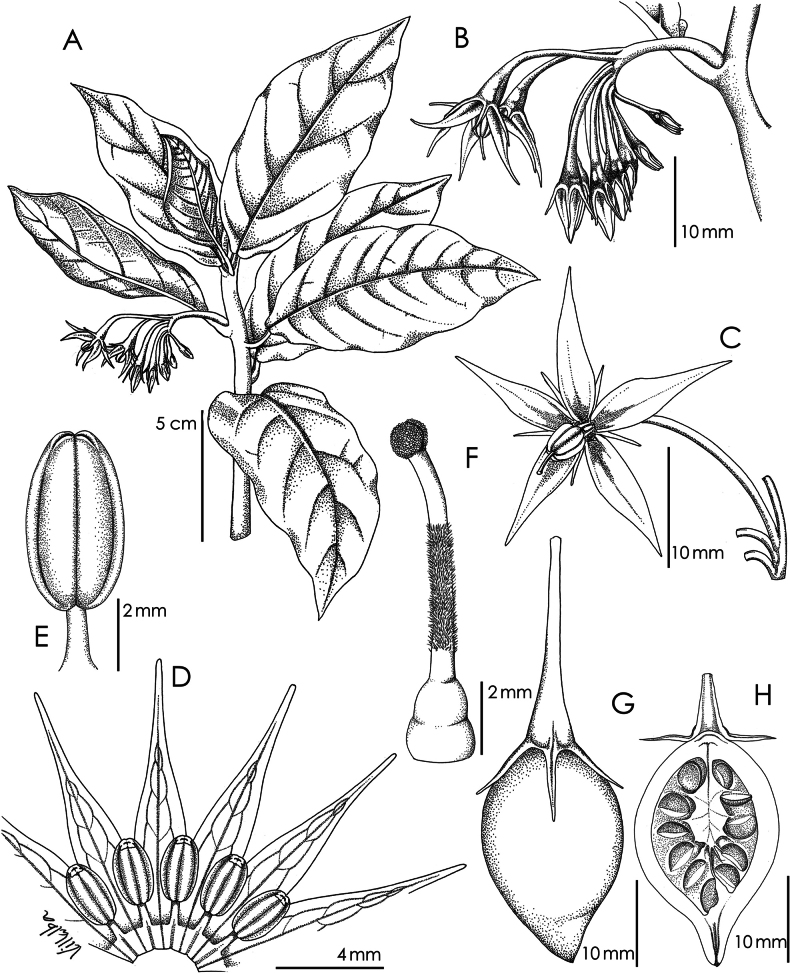 Figure 2
Figure 2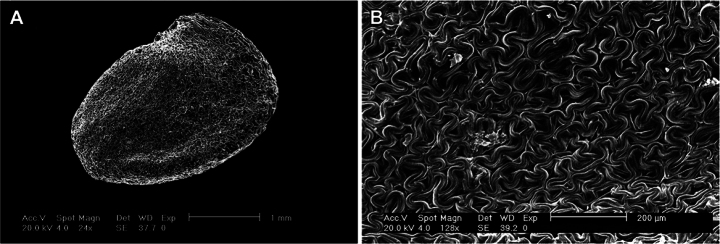 Figure 3
Figure 3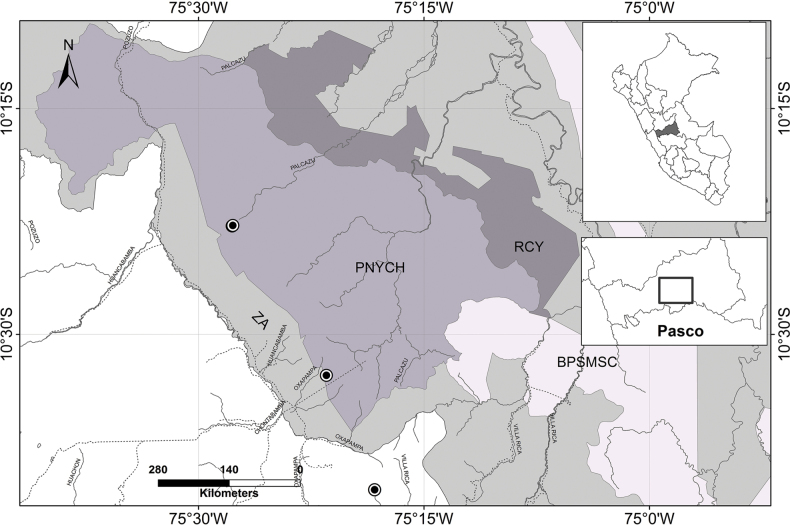 Figure 4
Figure 4| Attributes |
|
|
|
|
|---|---|---|---|---|
| Habit | Single‐stemmed shrub or herb | Single‐stemmed shrub or herb | Single‐stemmed shrub or herb | Climbing or single‐stemmed shrub or herb |
| Stem form | Terete | Terete | Angular or winged | Terete |
| Mature leaf width | 1.6–4.3 cm | 5–18.5 cm | 4–15 cm | 3.9–10.5 cm |
| Petiole form | Cylindrical canaliculate | Cylindrical canaliculate | Winged | Cylindrical canaliculate |
| Petiole length | 0.4–1.2 cm | 1–5 cm | 0–3 cm | 4–12.5 cm |
| Bud shape | Conical | Ovoide or globose | Globose | Ovoid, ellipsoid |
| Corolla in bud | Enclosed in calyx lobes | Exserted from calyx lobes | Exserted from calyx lobes | Exserted from calyx lobes |
| Corolla diameter | 17–20 mm | 5–7 mm | 4–5 mm | 12–25 mm |
| Corolla texture | Membranous | Membranous | Membranous | Fleshy |
| Corolla lobes at anthesis | Spreading | Strongly reflexed | Strongly reflexed | Spreading |
| Calyx lobe shape | Linear to narrowly triangular | Deltate, tips acute | Deltate, tips rounded | Deltate, tips acute to obtuse |
| Calyx lobe length | 3.5–4.5 mm | 0.5–1 mm | 0.5 mm | 1–2 mm |
| Fruit position | Deflexed to pendent | Erect | Erect | Pendent |
| Fruit shape | Ovoid-conical | Conical | Conical | Globose |
| Fruit surface | Smooth or slightly rugose | Rugose | Rugose | Smooth |
| Seed shape | Ovoid‐reniform | Ovoid‐reniform | Ovoid‐reniform | Flattened‐reniform |
| Seed number per berry | 18–27 | ca. 40 | ca. 20 | ca. 80–100 |
| Testal cell walls | Sinuate | Sinuate | Sinuate | Straight (rectangular) |
| 1 | Leaves simple (undivided) with entire margins |
|
| – | Leaves pinnatifid (apparently pinnate or ternate), with distinct leaflets |
|
| 2 | Corollas 17–25 mm in diameter at anthesis, the lobes spreading at anthesis, 8–12 mm long, fleshy or membranous; mature fruits pendulous; pedicels 17–22 mm long; berries conical ovoid or spherical, yellowish green or orange, smooth or slightly rugose |
|
| – | Corollas 3–10 mm in diameter at anthesis, the lobes reflexed at anthesis, 3–5 mm long, membranous; mature fruits erect; pedicels 7–10 mm long; berries conical or ovoid, green, the surface very rugose, rarely smooth |
|
| 3 | Inflorescences with peduncles and pedicels generally dark purple; calyx with linear or narrowly triangular lobes; corolla lobes membranous, narrowly lanceolate, the apex elongate-acuminate; mature berries ovoid-conical, yellowish green, smooth or rugose; seeds ovoid reniform, brown (Peru [Pasco]) |
|
| – | Inflorescences with peduncles and pedicels generally whitish green; calyx with triangular or deltate lobes; corolla lobes fleshy, if membranous, always broadly triangular, the apex acute; mature berries globose, orange (ovoid and white when immature), always smooth; seeds flattened reniform, whitish cream or pale tan (Ecuador; Peru [Cusco, Huancavelica, Huánuco, Junín. Loreto, Pasco]) |
|
| 4 | Stems cylindrical; leaves distinctly petiolate (Bolivia; Brazil; Colombia; Ecuador; French Guiana; Guyana; Peru [Amazonas, Cajamarca, Cusco, Huánuco, Junín, Loreto, Madre de Dios, Pasco, San Martín, Ucayali]; Suriname) |
|
| – | Stems strongly angular or winged; leaves strongly decurrent onto the stems, sessile (Peru [San Martín]) |
|
| 5 | Climbing herbs or woody vines |
|
| – | Terrestrial herbs or wand-like subshrubs, occasionally in large colonies |
|
| 6 | Woody or semi-woody vines, the basal stems to several cm in diameter; flowers 1.6–2 cm in diameter, pinkish tinged or white; petals spreading, fleshy; fruit globose, the surface smooth (Bolivia; Colombia; Ecuador; Peru [Amazonas, Cajamarca, Cusco, Huánuco, Junín, Pasco, San Martín, Ucayali]); Venezuela) |
|
| – | Herbaceous vines, often adhering to tree trunks; flowers 0.6–1 cm in diameter, greenish white; petals strongly reflexed, membranous; fruit conical, the surface rugose (Brazil; Colombia; Ecuador; Peru [Cusco, Huánuco, Junín, Loreto, Madre de Dios, Pasco, San Martín, Ucayali]) |
|
| 7 | Leaves with 5 or fewer pairs of lateral leaflets; leaflets usually obovate and widest in the distal third, especially the terminal leaflet |
|
| – | Leaves with more than 5 pairs of lateral leaflets; lateral leaflets lanceolate to elliptic; terminal leaflet similar in shape to the laterals |
|
| 8 | Leaves pubescent on veins and lamina on both surfaces |
|
| – | Leaves glabrous on lamina, occasionally pubescent along the veins and rachis |
|
| 9 | Fruit conical, the surface slightly rugose; leaf pubescence denser adaxially; basal leaflet pair markedly smaller than the other lateral leaflets (Ecuador; Peru [Amazonas, Pasco, San Martín]) |
|
| – | Fruit globose or occasionally apically pointed, the surface smooth; leaves equally pubescent on both surfaces; lateral leaflets all more or less the same size (Bolivia; Brazil; Colombia; Ecuador; Peru [Cajamarca, Cusco, Huánuco, Junín, Loreto, Madre de Dios, Pasco, San Martín, Ucayali]) |
|
| 10 | Fruit conical, the surface rugose; leaves fleshy; plants often rooting at the nodes, very small (Peru [Amazonas, Pasco, San Martín]) |
|
| – | Fruit globose or occasionally apically pointed, the surface smooth; leaves membranous; plants often semi-woody and up to 1 m tall (Bolivia; Brazil; Colombia; Ecuador; Peru [Cajamarca, Cusco, Huánuco, Junín, Loreto, Madre de Dios, Pasco, San Martín, Ucayali]) |
|
| 11 | Flowers 0.5–0.6 cm in diameter, the corolla lobes strongly reflexed at anthesis; fruit globose or occasionally apically pointed (Bolivia; Brazil; Colombia; Ecuador; Peru [Cajamarca, Cusco, Huánuco, Junín, Loreto, Madre de Dios, Pasco, San Martín, Ucayali]) |
|
| – | Flowers 0.9–1.3 cm in diameter, the corolla lobes usually spreading or only slightly reflexed at anthesis; fruit conical, rugose |
|
| 12 | Leaflets with petiolules 3–17 mm long; leaflets somewhat oblique and truncate at the base; leaves densely pubescent in a groove along the rachis adaxially; flowers usually more than 1 cm in diameter (Ecuador; Peru [Amazonas, Cajamarca, Cusco, Huánuco, Madre de Dios, Pasco, San Martín, Ucayali]) |
|
| – | Leaflets sessile or with short petiolules ca. 1 mm long; leaflets acute to attenuate at the base; leaves glabrous or if on sparsely pubescent then only abaxially; flowers less than 1 cm in diameter (Costa Rica, Guatemala, Mexico, Panama, Venezuela) |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Botany and Geology in Latin America and Caribbean · Fern and Epiphyte Biology
Introduction
Solanum L. (Solanaceae) is a cosmopolitan genus and the most species-rich within the family Solanaceae. It comprises approximately 1,250 species worldwide, and it is considered one of the most species-rich genera of flowering plants (Moonlight et al. 2024). Many species of Solanum are cultivated and of economic importance as sources of food (e.g., potatoes, tomatoes) and alkaloids for the pharmaceutical industry (Hawkes 1999). Species of Solanum are found in a variety of habitats, but the greatest species richness is concentrated in the tropics of South America (Knapp 2002; Hilgenhof et al. 2023). Solanum species are herbs, shrubs, and even large trees, with pentamerous flowers, fused sepals and petals, star-shaped to rotate corollas, stamens with short filaments and connivent anthers with poricidal dehiscence (Nee 1999; Bohs 2005; Hilgenhof et al. 2023). In light of its large number of species and varied morphology, Solanum has been classified under different systems at the infrageneric level (D’Arcy 1972; Child 1990, 1998; Nee 1999) within which several sections or groups of species have been identified based on morphological studies. Recently, studies using molecular markers have confirmed the monophyly of many of these, and a system of informal clade names has been established (Gagnon et al. 2022; Hilgenhof et al. 2023).
The Pteroidea clade (Solanum section Pteroidea Dunal) is a monophyletic group (Tepe et al. 2016; Gagnon et al. 2022) comprising ten erect herbaceous or climbing species, with simple or compound (deeply pinnatifid without interstitial leaflets) leaves and with axillary scorpioid inflorescences, many of the species have rugose conical fruits (Knapp and Helgason 1997). The axillary inflorescences found in all species of the group are unique in Solanum and are the distinguishing apomorphy of the clade (Hilgenhof et al. 2023). Species of the group are found from Mexico to Bolivia, with a centre of diversity in the eastern Amazonian premontane or montane forests in Peru and Ecuador (Knapp and Helgason 1997). Recent DNA studies support the monophyly of group and its relationship with Solanum section Herpystichum Bitter, both considered within the larger clade informally known as the “Potato clade”, which also includes the wild relatives of the potato, tomato and pepino (Bohs 2005; Tepe and Bohs 2010; Tepe et al. 2016).
In the most recent taxonomic review of the group (Knapp and Helgason 1997), the ten known species were divided into two informal groups: 1) the “Solanum ternatum group”, with S. incurvum Ruiz & Pav. and S. ternatum Ruiz & Pav., characterized by larger fleshy corollas up to 2 cm in diameter at anthesis with triangular lobes and globose and smooth fruits with numerous flat and reniform seeds; and 2) the “Solanum mite group”, with S. anceps Ruiz & Pav., S. angustialatum Bitter, S. chamaepolybotryon Bitter, S. conicum Ruiz & Pav., S. mite Ruiz & Pav., S. savanillense Bitter, S. trizygum Bitter, and S. uleanum Bitter, all characterized by having small membranous corollas less than 1 cm in diameter at anthesis with strongly very reflexed lobes, rugose conical fruits (except for S. mite that possesses globose fruits), and with few ovoid reniform seeds. Of these species only S. anceps, S. angustialatum, and S. incurvum have simple leaves, the rest all have imparipinnate pinnatifid leaves with three to more leaflets (Knapp and Helgason 1997). Solanum anceps is one of the species with the widest distribution inhabiting the Amazonian region and montane forests of Peru and Bolivia up to 2,000 m in elevation. Solanum angustialatum is only known from premontane forests in the Department of San Martín in Peru. Solanum incurvum grows in cloud forests close to 3,000 m in elevation and is poorly collected; it is known just from a few localities in southern Ecuador and central Peru (Knapp and Helgason 1997).
While undertaking taxonomic revision of the Solanaceae in the Yanachaga-Chemillén National Park, we collected several specimens of an unusual species of Solanum that we recognized as a simple-leaved member of the Pteroidea clade distinctly different from any currently known species in the group. Here we describe this new species and discuss its distribution, relationships, and conservation status.
Materials and methods
This study was conducted primarily using field-collected material. We analyzed living plants of all species of the Pteroidea clade with simple leaves and herbarium specimens available at the following herbaria: AMAZ, BM, CUZ, HAO, HOXA, HUT, HUSA, K, MA, MOL and USM. We also examined the types of S. anceps and S. incurvum at MA, and S. angustialatum at K. Fine-scale morphological details were observed with stereoscopic and compound microscopy. Seeds were extracted from mature fruits, air-dried and mounted on aluminum stubs previously furnished with carbon adhesive tape on both sides. Mounted seeds were then covered with gold-palladium in a Denton Vacuum Dex II metal-supplier for approximately 60 seconds and then observed with a Phillips XL 20 Scanning Electron Microscope (SEM) at the Center for Electron Microscopy of Universidad Nacional de San Agustín, Arequipa, Peru. Distribution of S. stellaticalycinum is based on geographical information from herbarium specimens; where coordinates were not recorded on labels they were calculated using Google Earth. A preliminary conservation status was assessed using the IUCN (2022) criteria. All specimens were collected in accordance with Peruvian regulations under permits 004-2013-SERNANP-DGANP-JEF and 026C/C-2008-INRENA-IANP issued by the national environmental authority (SERNANP).
Taxonomic treatment
Solanum
stellaticalycinum
Taxon classificationPlantaeSolanalesSolanaceae
M.A.Cueva, Treviño & D.Sotomayor sp. nov.
6BC1FD64-C1CC-59CC-BD84-6160ECC54ED5
urn:lsid:ipni.org:names:77371677-1
Diagnosis.
Solanum stellaticalycinum differs from all other species of the Pteroidea clade in its calyx with narrow linear lobes and corolla with narrow lanceolate lobes with elongate, acuminate apices that are spreading at anthesis. It differs from the sympatric S. anceps Ruiz & Pav. in its larger flowers and fruits on pendent rather than erect pedicels, and from S. mite Ruiz & Pav. and S. conicum Ruiz & Pav. in its simple, rather than deeply pinnatifid, leaves.
Type.
Peru • Pasco: Prov. Oxapampa, Distrito de Huancabamba, Parque Nacional Yanachaga‐Chemillén, Sector Abra Yanachaga, camino hacia parcela, 10°22'48"S, 75°27'45.25"W, 2,880 m, 2 Oct 2009, M. Cueva, A. Monteagudo, A. Peña, J. Mateo, R. Rivera & V. Peña 645 (holotype: HUSA [acc. # 10917]; isotypes: USM [acc. # 253156], HOXA, MO [MO-4348867, acc. # 7105417], HUT).
Description.
Erect, single-stemmed herbs or subshrubs, 0.4–2 m tall. Stems 0.4–1.3 cm in diameter, terete; young growth minutely papillose glandular; internodes 0.7–1.2 cm long, brownish green, pubescent with whitish gray 7-8-celled simple uniseriate trichomes 0.5–0.7 mm long; older internodes 1.3–2.7 cm long, glabrous or glabrescent, dark brownish green. Sympodial units unifoliate. Leaves simple, entire; blades 7–10 cm long, 1.6–4.3 cm wide, elliptic to ovate, membranous, discolorous, both surfaces glabrous or with a few simple uniseriate trichomes ca. 0.5 mm long along the veins and minutely papillate with tiny glandular hairs; adaxial surface dark green and shiny (dark brown when dry); abaxial surface pale green or with purple veins (yellowish brown when dry); principal veins 9–10 pairs, slightly raised on the abaxial surface; base cuneate to attenuate; margins entire; apex acute to acuminate; petioles 0.4–1.2 cm long, green or purple, glabrous, with tiny papillae like those of the stems and leaves. Inflorescences axillary scorpioid cymes, (7–) 10–25 mm long, with 4–13 flowers generally inserted in the distal 1/3 of the axis,1–4 flowers open at a time; peduncle 6–17 mm long, dark purple, shiny, glabrous or glabrescent, with papillate trichomes like those of stems and leaves; pedicels 10–14 (–16) mm long at anthesis, ca. 0.8 mm in diameter at the base widening to ca. 2 mm in diameter at the apex, dark purple, glabrous or with some dispersed simple uniseriate trichomes like those of the stems and leaves, articulated at the base, deflexed at maturity so the flowers face downwards; pedicel scars slightly raised, closely spaced. Buds tapering-ellipsoid, enclosed within the linear calyx lobes. Flowers 5-merous, cosexual. Calyx with the tube obconical, ca. 2 mm long, 3–3.4 mm in diameter, dark purple, shiny, glabrous or sparsely white-pubescent with 6-7-celled simple uniseriate trichomes up to 0.4 mm long and some tiny papillae, the lobes (2–) 3.5–4.5 mm long and 0.5 mm wide, linear to narrowly triangular, dark purple with white or green margins, glabrescent with whitish 6-7-celled simple uniseriate hairs abaxially, brown-glandular papillae on the proximal half and extending towards the distal portion of the calyx tube adaxially. Corolla 17–20 mm in diameter at anthesis, deeply stellate, divided almost to the base, the lobes 8–11.5 mm long, ca. 2 mm wide, lanceolate, spreading at anthesis, membranous, white with dark purple spots, the margins puberulent with 1-3-celled simple uniseriate hairs, less than 0.2 mm long, both surfaces glabrous, the apex elongate-acuminate. Stamens equal; filament tube minute; free portion of the filaments ca. 1.5 mm long, glabrous, white; anthers 3–4 mm long, 1.5–2 mm wide, oblong to oblong-ellipsoid, yellow, poricidal at the tips, the pores introrse, lengthening to slits with age. Ovary conical ca. 5 mm long and 1 mm in diameter, glabrous; style 6–6.5 mm long, ca. 0.5 mm in diameter, exserted 2.5–3.5 mm beyond the anthers, white, straight, densely pubescent in the basal half with 1-3-celled simple uniseriate hairs up to 0.2 mm long; stigma ca. 0.5 mm long, capitate and slightly bilobed, white, the surface minutely papillose. Fruit an ovoid-conical berry, 1.3–2.1 (–2.7) cm long, 0.9–1.3 (–1.7) cm in diameter, yellowish green, the pericarp shiny, smooth or slightly rugose; fruiting pedicels 1.7–1.8 cm long, deflexed to pendent, the calyx persistent, the lobes 3–4 mm long, linear to narrowly triangular. Seeds 18–27 per berry, ovoid-reniform, ca. 3 mm long, 2.5 mm wide, brownish green, the testal cells deeply sinuate, with anticlinal walls ca. 10 µm in thickness; stone cells absent. Chromosome number: not known.
Distribution
**(Fig. 4).**Solanum stellaticalycinum is endemic to the Department of Pasco (Prov. Oxapampa) in Peru. All specimens have been collected in the Yanachaga-Chemillén National Park and surrounding areas (Fig. 4), in the sectors of Abra Yanachaga, San Alberto and Chacos.
Ecology and habitat.
Solanum stellaticalycinum grows as a single stemmed herb (Figs 1A, 2A) in the understory of primary montane forests, often close to the somewhat open borders of rural paths, between 2,399 and 3,000 m elevation. Plants have been collected in flower between August and November (Fig. 1A), and fruiting specimens in April and October. It is likely that these plants bloom irregularly all year round, as do many other solanums from similar habitats.
Solanum stellaticalycinum M.A.Cueva, Treviño & D.Sotomayor. A, B. Habit; C. Inflorescence; D. Flower at full anthesis; E. Floral buds; F. Fruit (M. Cueva et al. 645). Photos by M.A. Cueva.
Solanum stellaticalycinum. A. Habit; B. Inflorescence; C. Flower; D. Floral dissection; E. Anther detail; F. Gynoecium detail. G, H. Fruit longitudinally dissected to reveal seeds (based on M. Cueva et al. 645). Illustration by Maria I. Villalba.
Images of the seed coat using SEM from Solanum stellaticalycinum. A. Whole seed; B. Testal cell shape detail. Scale bar in image.
Distribution of Solanum stellaticalycinum. Abbreviations: Yanachaga‐Chemillén National Park (PNYCH), Buffer Zone (ZA), Reserva Comunal Yanesha (RCY), Bosque de Protección San Matías San Carlos (BPSMSC).
Etymology.
The species epithet refers to the shape of the calyx, which is like a star (see Figs 1, 2). This character is not present in any other species of the Pteroidea clade.
Preliminary conservation status
**(IUCN 2022).**Solanum stellaticalycinum is here assigned a preliminary threat status of Endangered (EN), using the criteria B1ab (i, iii). It is only known from three localities, and the known range (EOO) is approximately 111.9 km^2^. It is likely that this species is distributed along the entire extension of the Yanachaga range, which occurs within the Yanachaga-Chemillén National Park, an area protected by the Peruvian government. It possible however, that in the future, the area of occupancy of this species will shrink in response to habitat modification resulting from human activities in buffer zone areas of the park.
Additional specimens examined
(paratypes). Peru • Pasco: Prov. Oxapampa: Dist. Oxapampa, Yanachaga-Chemillén National Park, San Alberto sector, surroundings of Refugio el Cedro, 2483 m, 10°32'43.3"S, 75°21'29.5"W, 26–27 Apr 2009, M. Cueva & R. Rivera 614 (USM); New highway between Oxapampa-Bosque Shollet-Villa Rica, 2399 m, 25 Oct 2013, M. Cueva & S. Smith 1700 (HUSA); • Dist. Huancabamba, Abra Yanachaga, Yanachaga-Chemillén National Park, 2900–3000 m, 10°22'46"S, 75°27'43"W, 23 Nov 2004, A. Monteagudo, A. Peña, J. Perea, R. Francis, J. L. Mateo & E. Becerra 7840 (BM [BM015735503], HOXA [acc. # 00012525], USM, HUT, AMAZ, MOL, MO [MO-2265895, acc. # 6217980]); • Dist. Huancabamba, Abra Yanachaga, Yanachaga-Chemillén National Park, 2930 m, 10°22'46.3"S, 75°27'42.9"W, 03 Aug 2005, E. Ortiz & R. Francis735 (HOXA [acc.# 00018108], USM, MO [MO-1662368, acc. # 4824338]).
Discussion.
Solanum stellaticalycinum is not easily confused with any other species of Pteroidea. The flowers with linear calyx lobes alternating with the lanceolate corolla lobes (Fig. 1D) are unique in the clade, and unusual in Solanum overall. It shares membranous corollas, conical ovoid fruits with smooth or rugose surfaces, and ovoid reniform seeds with irregularly shaped testal cells with sinuate anticlinal walls (Fig. 3) with other members of the S. mite species group (sensu Knapp and Helgason 1997), where only S. anceps and S. angustialatum have simple leaves. The other six known taxa in the S. mite group (S. conicum, S. chamaepolybotryon, S. mite, S. savanillense, S. trizygum and S. uleanum) have compound imparipinnatifid leaves with three or more leaflets (Knapp and Helgason 1997). Solanum anceps and S. angustialatum are morphologically similar and both differ from S. stellaticalycinum in their whitish green pedicels 4–7 mm long (versus purple pedicels 10–14 mm long, Figs 1E, 2B, C), very small flowers with delate calyx lobes up to 1 mm long (versus linear calyx lobes 3.5–4.5 mm long, Figs 1C, F, 2), corollas at anthesis 3–10 mm in diameter with very reflexed membranous triangular lobes 3–5 mm long (versus corolla at anthesis 17–20 mm in diameter, with extended, membranous, narrowly lanceolate lobes 8–11.5 mm long, Figs 1C–F, 2B, C ,G and anthers ca. 2 mm long (versus 3–4 mm long in S. stellaticalycinum, Fig. 2E).
Solanum stellaticalycinum is also somewhat similar to the sympatric S. incurvum from the S. ternatum group (sensu Knapp and Helgason 1997). Solanum incurvum differs from S. stellaticalycinum in its calyx with deltate lobes ca. 1 mm long (versus linear lobes 3–4 mm long), a larger fleshy corolla generally 23–25 mm in diameter with triangular lobes (versus membranous corollas 10–14 mm in diameter), and in its globose smooth fruits with flattened reniform seeds (versus conical fruits with ovoid reniform seeds in S. stellaticalycinum). Some of the examined individuals of S. anceps and S. incurvum exhibit variation regarding fruit texture and corolla consistency. Some specimens of S. anceps develop smooth ovoid fruits with only slight whitish rugosities, similar to those seen in some specimens of S. stellaticalycinum, but the pedicels are still short and erect rather than pendent, and corolla diameter does not exceed 10 mm in diameter. In some individuals of S. incurvum, the corollas can develop a membranous consistency, and fruits can be ovoid (and somewhat flattened) in the first stages of their development, but the deltate or triangular shape of both calyx and corolla lobes, as well as the flattened reniform seeds are distinctive. Morphological differences amongst the simple-leaved species of Pteroidea are summarized in Table 1.
Table 1.: Morphological differences between the simple-leaved members of Pteroidea: Solanum stellaticalycinum, S. anceps, S. angustialatum, and S. incurvum.
Artificial key to all species of the Pteroidea clade
[Note: Total country distribution noted for each species noted; distribution to Department level included for Peru. An online multiaccess key for all species of Solanum, including Pteroidea, can also be found at https://solanum-species.identificationkey.org/mkey.html.]
**: **
Supplementary Material
XML Treatment for Solanum stellaticalycinum
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bohs L (2005) Major clades in Solanum based on ndh F sequences. In: Keating RC Hollowell VC Croat TB (Eds) A festschrift for William G.D’Arcy: the legacy of a taxonomist, Monographs in Systematic Botany from the Missouri Botanical Garden, Vol. 104. Missouri Botanical Garden Press, St. Louis, 27–49.
- 2Child A (1990) A synopsis of Solanum subgenus Potatoe (G. Don) D′Arcy (Tuberarium (Dun.) Bitter (s.l.)).Feddes Repertorium 101: 209–235. 10.1002/fedr.19901010502 · doi ↗
- 3Child A (1998) Studies in Solanum and related genera (6). New infrageneric taxa for the genus Solanum L. (Solanaceae).Feddes Repertorium 109: 407–427. 10.1002/fedr.19981090512 · doi ↗
- 4D’Arcy WG (1972) Solanaceae studies II: typification of subdivisions of Solanum.Annals of the Missouri Botanical Garden 59: 262–278. 10.2307/2394758 · doi ↗
- 5Gagnon E Hilgenhof R Orejuela A Mc Donnell A Sablok G Aubriot X Giacomin L Gouvêa Y Bragonis T Stehmann JR Bohs L Dodsworth S Martine C Poczai P Knapp SSärkinen T (2022) Phylogenomic discordance suggests polytomies along the backbone of the large genus Solanum. American Journal of Botany 109: 580–601. 10.1002/ajb 2.1827 PMC 932196435170754 · doi ↗ · pubmed ↗
- 6Hawkes JG (1999) The economic importance of the family Solanaceae. In: Nee M Symon DE Lester RN Jessop JP (Eds) Solanaceae IV Advances in Biology and Utilization.Royal Botanic Gardens Kew, Richmond, 1–8.
- 7Hilgenhof R Gagnon E Knapp S Aubriot X Tepe EJ Bohs L Giacomin LL Gouvêa YF Martine CT Orejuela A Orozco CI Peralta IESärkinen T (2023) Morphological trait evolution in Solanum (Solanaceae): Evolutionary lability of key taxonomic characters.Taxon 72: 811–847. 10.1002/tax.12990 · doi ↗
- 8IUCN (2022) Guidelines for Using the IUCN Red List Categories and Criteria. Version 13. Prepared by the Standards and Petitions Subcommittee of the IUCN Species Survival Commission. http://www.iucnredlist.org/documents/Red List Guidelines.pdf