Some Virulence-Associated Genes of Proteus Isolates Could Predict Antibiotic Susceptibility and Even Infection Source
Narges Jafari, Roya Ahmadrajabi, Omid Tadjrobehkar

TL;DR
This study shows that certain genes in Proteus bacteria can predict antibiotic resistance and whether the infection is from a hospital or community source.
Contribution
The study identifies specific virulence genes as potential biomarkers for predicting antibiotic susceptibility and infection origin in Proteus isolates.
Findings
mrpA and atfA genes predict sensitivity to certain antibiotics in Proteus isolates.
rsbA gene helps distinguish community-acquired from hospital-acquired isolates.
Community-acquired isolates showed higher virulence compared to hospital-acquired ones.
Abstract
In the present study, the probable association of virulence-associated genes (VAGs) with antibiotic resistance and also sample sources in Proteus isolates was investigated. Then, 91 Proteus mirabilis and nine Proteus vulgaris were used in this study. The disk diffusion method was used in order to perform an antibiotic susceptibility assessment. A combination double-disc synergy test was used for the evaluation of extended-spectrum β-lactamases. Eight VAGs were investigated by polymerase chain reaction (PCR) method. ERIC-PCR fingerprinting was also performed for P. mirabilis isolates. Maximum frequency of resistance was detected against trimethoprim–sulfamethoxazole combination in P. mirabilis isolates and against cefalexin in P. vulgaris isolates. Then, 6% of isolates were multidrug-resistant (MDR) and all were P. mirabilis. Community-acquired (CA) isolates were more virulent than…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
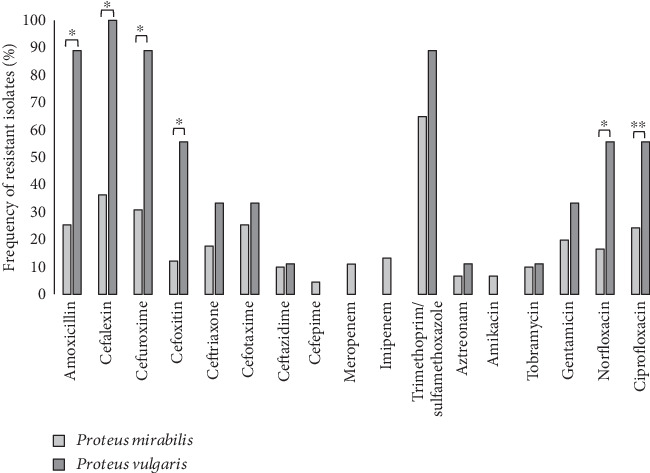 Figure 1
Figure 1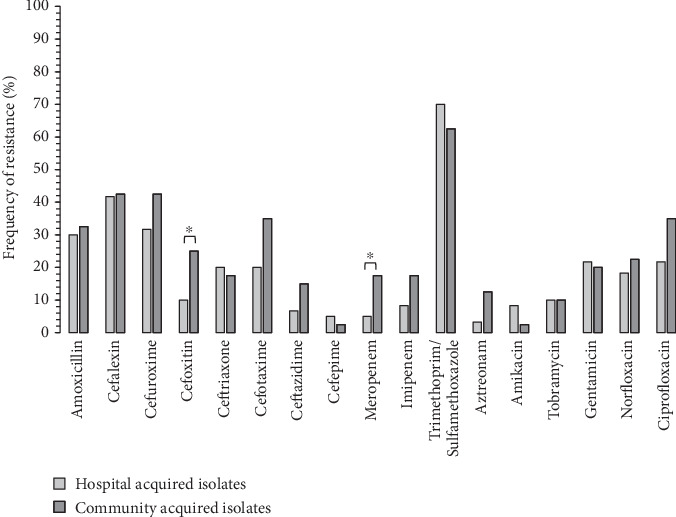 Figure 2
Figure 2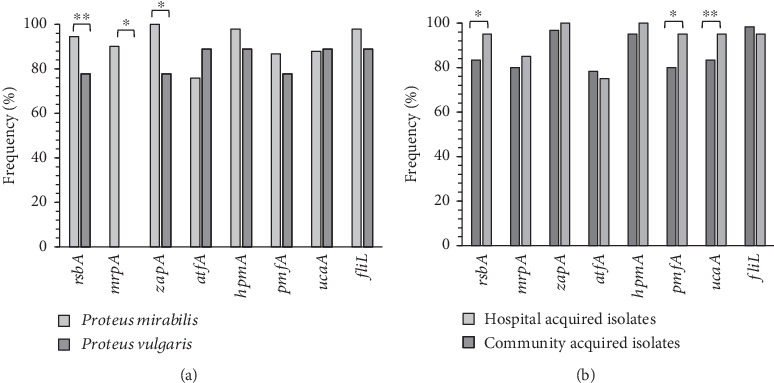 Figure 3
Figure 3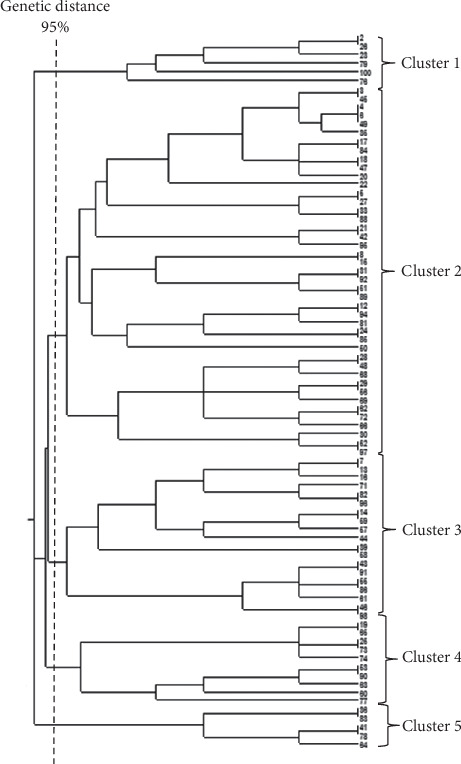 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Genetics and Biotechnology · Antibiotic Resistance in Bacteria · Antimicrobial Resistance in Staphylococcus
1. Introduction
Proteus mirabilis and Proteus vulgaris were reported as the most frequent opportunistic pathogenic species of Proteus that were isolated from clinical samples [1]. Proteus species are involved in different types of infections including urinary tract infection (UTI), bacteremia, indwelling catheter-associated infections, wound infections, and also localized lesions in elderly and diabetic persons [2–4]. Proteus is also a major cause of nosocomial infections, especially catheter-associated urinary tract infections (CAUTIs) [5, 6].
A massive storage of virulence-associated genes (VAGs) was detected in the Proteus species. Diverse gene clusters express different adhesins (17 fimbriae operons) in the Proteus species [7]. Among them, mannose-resistant/Proteus-like (MR/P) fimbriae, Proteus mirabilis fimbriae (PMF), and Proteus mirabilis P-like pili (PMP) are the most studied, and they play critical roles in the colonization of P. mirabilis in the uroepithelium of humans [8].
MR/P fimbria is composed of two structural subunits, with the main subunit encoded by the mrpA gene and mrpBEFG encoding other subunits. The MR/P fimbriae are involved in the colonization of the urinary tract and also biofilm formation, especially in UTI cases [4, 9]. PMF is another fimbria of P. mirabilis, which is encoded by the pmfACDEF operon and is involved in the colonization of the bacterium in the kidney and bladder epithelium [10]. Uroepithelial cell adhesin (UCA) is another fimbria of the Proteus genus that is optimally expressed at the ambient temperature (23°C), and it facilitates the attachment of the Proteus species to the glycolipid molecules of the human uroepithelium under in vitro conditions [9, 10].
Several factors are associated with the destruction of the host tissues by Proteus species, including toxins, lipopolysaccharide (LPS), vigorous motility, swarming, and proteolytic enzymes [11, 12]. HpmA and HlyA are the two most important hemolysins of the Proteus genus. The hpm locus contained two associated genes, hpmA, which is a Ca2+-dependent pore-forming toxin, and hpmB, which is a toxin activator [13]. HpmA has a critical role in UTI pathogenesis [14] and also in biofilm formation [15].
The fliL gene is a member of the fliLMNOPQR locus that encodes a basal body protein of the flagellum [16]. The fliL has a critical role in the control of swimming and swarming behavior of P. mirabilis, and it also stimulates the expression of the zapA and hpmA genes [17]. The RsbA is a protein sensor of P. mirabilis and is encoded by the rsbA gene. The regulatory activities of the rsbA gene on swarming motility and biofilm formation have been reported recently [13, 14]. The zapA gene encodes a zinc metalloprotease of P. mirabilis with multifunctional properties. ZapA protease cleaves IgA and complement components of the human serum and protects bacteria against the host immune responses [5, 13].
Antibiotic resistance continuously has been regarded as a major global problem in health systems from the beginning of the antibiotic era (1950s) till now. It is suggested that antibiotic–resistance genes (ARGs) have evolved in the surrounding environment even before upcoming new antibiotics developed [18]. Enterobacteriaceae members such as Proteus are known as important antibiotic-resistant human pathogens. Proteus species are intrinsically resistant to polymyxin antibiotics, ampicillin, tetracycline, and nitrofurantoin, and also reduced susceptibility to other agents has been reported repeatedly [1, 19]. In the present study, we first tried to investigate the antibiotic susceptibility profile of Proteus species, and also, a detailed arsenal of VAGs has been detected. In the next step, the association between antibiotic resistance and VAGs has been analyzed. We also tried to find logical relationships between antibiotic resistance and VAGs.
2. Material and Methods
2.1. Specimen Collection
In the present study, some suspected Proteus isolates were cultivated by laboratory practitioners from hospitalized and outpatient individuals referred to different laboratories and medical centers affiliated with Kerman University of Medical Sciences. Proteus isolates were obtained from different samples (urine, surgical wounds, and burning wounds). They were transferred immediately to the bacteriology laboratory of Afzalipour School of Medicine for confirmatory identification and also species differentiation. Finally, 100 confirmed bacterial isolates (91 Proteus mirabilis and 9 Proteus vulgaris) were used in all experiments. The isolates that were isolated immediately after admission to the hospitals or within the first 48 h of hospitalization were referred to as community-acquired (CA), and the isolates that were obtained after 48 h of the hospitalization period were regarded as hospital-acquired (HA) [20, 21].
2.2. Culture and Identification
Proteus isolates were identified by standard biochemical and molecular tests [22, 23]. The isolated bacteria were stored in trypticase soy broth enriched with 20% glycerol at −70°C for subsequent use.
2.3. Antimicrobial Susceptibility Test
Disk diffusion method was used for antibiotic susceptibility assessments of Proteus isolates according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI) [24].
Then, 20 antibiotic discs from the following classes: β-lactams including amoxicillin (25 μg), cefalexin (30 μg), cefuroxime (30 μg), cefoxitin (30 μg), ceftazidime (30 μg), ceftriaxone (30 μg), cefotaxime (30 μg), cefepime (30 μg), aztreonam (30 μg), imipenem (10 μg), and meropenem (10 μg); fluoroquinolones including norfloxacin (10 μg) and ciprofloxacin (5 μg); aminoglycosides including tobramycin (10 μg), gentamicin (10 μg), amikacin (30 μg), and trimethoprim/sulfamethoxazole (1.23/25.75 μg) as folate synthesis inhibitors were used in order to assess antibiotic susceptibility. Antibiotic discs were provided by Padtan-Teb.Co.(Iran). E. coli ATCC 25922 was used for quality control of the disk diffusion method. All experiments were performed in triplicate.
2.4. Extended Spectrum β-Lactamase (ESBL) Screening and Confirmatory Test
Isolates showed primary resistance against aztreonam, ceftazidime, and cefotaxime in the screening disk diffusion method and were regarded as ESBL producers. ESBL-producing isolates were confirmed using two pairs of antibiotic disks: ceftazidime (30 μg), ceftazidime–clavulanic acid (30 μg/10 μg), cefotaxime (30 μg), and cefotaxime–clavulanic acid (30 μg/10 μg) through the combination double-disc synergy test (CDDST) according to CLSI guidelines [24, 25].
2.5. Bacterial Cell Lysate Preparation and DNA Extraction
In summary, a loopful of an overnight culture of bacterial isolates was inoculated into the microtubes including 500 μL of distilled water. The microtubes were heated in a water bath at 100°C for 10 min. In the next step, the sample was centrifuged (12,000 rpm for 5 min). After centrifugation, the supernatant was separated and kept at −70°C.
2.6. Polymerase Chain Reaction and Primer List
The eight VAGs (rsbA, mrpA, zapA, atfA, hpmA, pmfA, ucaA, and fliL) were investigated by PCR method using a Biometra thermocycler (Germany) and specific primers (purchased from Bioneer, South Korea). The primer sequences are shown in Table 1.
PCR products were separated by gel electrophoresis using a 1.5% agarose gel alongside 100-bp DNA ladder.
2.7. Enterobacterial Repetitive Intergenic Consensus (ERIC) PCR Fingerprinting
The following pair of recommended primers 5′ ATGTAAGCTCCTGGGGATTCAC 3′ and 5′ AAGTAAGTGACTGGGGTGAGCG 3′ was used in fingerprinting of P. mirabilis isolates by ERIC-PCR technique [29]. After separation of PCR products by gel electrophoresis, band sizes were recorded in the Excel software datasheets, and finally, band size patterns were analyzed by NTSYSpc v2-10e software, and a dendrogram was created through unweighted pair group method with arithmetic mean (UPGMA) by using Dice coefficient, and 95% genetic distance was used for allocation of isolates in different clusters.
2.8. Statistical Analysis
Data analysis was performed using SPSS 26 statistical software. Chi-square and Fisher's exact test were used to analyze the association between variables. Binary logistic regression analysis was used to evaluate the predictive role of VAGs for resistance or susceptibility to different antibiotic agents. All experiments were performed in triplicate and means were used. The p value ≤ 0.05 was considered significant.
3. Results
3.1. Distribution of Bacterial Isolates in Different Samples
Totally, 60 out of 100 isolates were regarded as HA; 54 (90%) of them were P. mirabilis and 6 (10%) isolates were confirmed as P. vulgaris. The prevalence of P. mirabilis and P. vulgaris in CA isolates was 37 (92.5%) and 3 (7.5%), respectively. The majority of P. mirabilis isolates, 82 (90.1%), were cultivated from urine samples; three (3.3%) isolates were cultivated from surgical wounds and six (6.6%) isolates were obtained from burning wounds. Eight (88.9%) out of nine P. vulgaris isolates were cultivated from urine samples and only one isolate (11.1%) was obtained from a surgical wound.
3.2. Antibiotic Susceptibility
Data collected from antibiotic susceptibility assessment showed various frequencies of resistance against the following antibiotic agents: β-lactam antibiotics including amoxicillin (31%), cefalexin (42%), cefuroxime (36%), cefoxitin (16%), ceftriaxone (19%), cefotaxime (26%), ceftazidime (10%), cefepime (4%), meropenem (10%), imipenem (12%), and aztreonam (7%), trimethoprim–sulfamethoxazole (67%) as a folate synthesis inhibitor, aminoglycosides including amikacin (6%), tobramycin (10%), and gentamicin (21%), and fluoroquinolones including norfloxacin (20%) and ciprofloxacin (25%).
Maximum antibiotic resistance was detected against cefalexin (100%) in P. vulgaris isolates, and maximum resistance was detected against trimethoprim–sulfamethoxazole (64.8%) in P. mirabilis. Minimum antibiotic resistance was detected against cefepime (4.4%) and amikacin (6.6%) in P. mirabilis. Resistance against cefepime, amikacin, imipenem, and meropenem was not detected in P. vulgaris isolates (Figure 1).
Isolates that were not susceptible to three antibiotic agents from different classes were regarded as multidrug-resistant [19]. Totally, six isolates (6%) were MDR, of which 5 were P. mirabilis and only 1 P. vulgaris isolate was MDR. No extremely drug-resistant (XDR) isolate was detected in our study. There were not significant differences between the frequency of MDR isolates among HA isolates and CA isolates.
Variable frequency of resistance was detected against different antibiotic agents among two studied species (Figure 1). Higher frequency of resistance against amoxicillin, cefalexin, cefuroxime, cefoxitin, norfloxacin, and ciprofloxacin was detected in P. vulgaris isolates in comparison to P. mirabilis isolates (Figure 1).
Data obtained from the double disk synergy test revealed that 12 (13.2%) of P. mirabilis isolates and one (11.1%) of P. vulgaris isolates were ESBL producers (totally 13% of Proteus isolates).
Pearson Chi-square analysis showed that resistance against cefoxitin (p = 0.021) and meropenem (p = 0.041) was significantly higher in CA isolates in comparison to the HA isolates. There were not observed significant differences regarding remaining antibiotic agents (Figure 2).
Fisher's exact test analysis showed that ESBL activity was significantly (p ≤ 0.05) more frequent in CA isolates (22.5% and 17.5%, respectively) in comparison to the HA isolates (6.7% and 5%, respectively).
3.3. Frequency of VAGs
Prevalence of studied VAGs among Proteus isolates was as follows: rsbA (93%), mrpA (82%), zapA (98%), atfA (77%), hpmA (97%), pmfA (86%), ucaA (88%), and fliL (97%).
Statistical analysis showed that the distribution of the zapA and mrpA genes was significantly different (Fisher's exact test analysis, p ≤ 0.05) between two investigated Proteus species. The mrpA gene was detected only in P. mirabilis isolates (Figure 3).
Chi-square analysis revealed that the rsbA gene was more prevalent in CA isolates in comparison to the HA isolates (100% against 88.3%, p = 0.024). Similar results were detected for the pmfA gene (95% against 80%, p = 0.034). Data have been presented in Figure 3.
Collection of all of the studied VAGs was detected in 47 (47%) out of 100 isolates. Pearson Chi-square analysis confirmed that these highly virulent isolates were harvested significantly (p = 0.027) more from the community (60%) than from the hospital (37.3%). Antibiotic resistance against different antibiotic agents was not significantly (p ≤ 0.05) different in these isolates than in the others.
3.4. Association of VAGs With Antibiotic Resistance
Pearson Chi-square analysis showed that the atfA gene is significantly associated with sensitivity to tobramycin (p = 0.032) and amikacin (p = 0.044). Data have been presented in Table 2. The binary regression analysis revealed that the atfA gene is a predictor of sensitivity to amikacin (p = 0.043, odds ratio: 1.188) and tobramycin (p = 0.019, odds ratio: 2.209).
Data analysis (Pearson Chi-square) also revealed that the mrpA gene was significantly associated with susceptibility against amoxicillin (p = 0.013), cefoxitin (p = 0.027), and norfloxacin (p = 0.027). Data have been presented in Table 2. The binary regression analysis also revealed that the mrpA gene is a predictor of sensitivity to cefoxitin (p = 0.005, odds ratio: 3.262), norfloxacin (p = 0.015, odds ratio: 1.817), and amoxicillin (p = 0.021, odds ratio: 2.317).
3.5. Genotyping Trough ERIC-PCR Fingerprinting Method
Only P. mirabilis isolates were typable by the ERIC-PCR method. Only 83 out of 91 P. mirabilis isolates were categorized in five clusters, and eight (8.8%) isolates were totally different from other isolates. Six isolates (6.6%) were placed in Cluster 1; Clusters 2 to 5 included 46 (50.5%), 16 (17.5%), 10 (11%), and 5 (5.5%) isolates, respectively (Figure 4). Statistical analysis did not show any significant association between different ERIC clusters and the sample or source type of isolates.
3.6. Frequency of Phenotypic Antimicrobial Resistance Among Different Clusters
Cluster 5 contains five CA urine isolates. These isolates were significantly different from other clusters regarding resistance against amoxicillin, cefalexin, cefoxitin, cefotaxime, ceftazidime, cefuroxime, and meropenem. Tobramycin and cefepime resistance were more frequent in Cluster 1 and Cluster 3, respectively (Table 3). Statistical analysis did not find any significant differences between other clusters regarding antibiotic resistance or virulence gene profile.
3.7. Frequency of VAGs Among Different Clusters
Frequency of rsbA gene was significantly different (Chi-square analysis, p = 0.001) among five ERIC clusters, and minimum frequency (60%) was detected among Cluster 5 isolates. Meanwhile, cumulative frequency of rsbA gene among other clusters was 96.2%. The rsbA gene was detected in all isolates in Clusters 1, 3, and 4, and 93.5% of isolates in Cluster 2 also had it.
4. Discussion
Uncontrolled use of antibiotic agents in various industries, including agriculture, animal husbandry, food-related industries, and also medicine during the last decades, has resulted in widespread evolution of antibiotic resistance in different bacterial pathogens [30]. In this context, reduced susceptibility of Proteus species to different antibiotic agents was also reported globally, and the spread of MDR and XDR strains has limited treatment options [1, 31].
In the present study, maximum resistance was detected against the trimethoprim-sulfamethoxazole combination in P. mirabilis isolates (64.8%). Mirzaei et al. also reported similar findings from a different geographic region of Iran recently [32]. The maximum resistance of P. mirabilis isolates against the trimethoprim–sulfamethoxazole combination was also reported from other countries [31, 33, 34].
Low frequency of resistance against different β-lactam antibiotics, aminoglycosides, and fluoroquinolones was observed in P. mirabilis isolates in the present study. Similar findings were also reported in a meta-analysis from Iran recently that support our findings [35]. However, increasing resistance against these antibiotic agents is also reported from other countries [31]. Low frequency of resistance against β-lactam antibiotics among P. mirabilis isolates in our study could also be explained by the low prevalence of ESBL-producing isolates (13%) among the investigated bacterial population. Similar findings were also reported by other researchers [36, 37]. Different frequencies of resistance against β-lactam antibiotics, fluoroquinolones, and aminoglycosides have been reported from different countries. It could be the result of different patterns of antibiotic prescription in various geographical regions worldwide, and horizontal transfer of ARGs within integrons, transposons, and plasmids between different Enterobacteriaceae members could be regarded as the major reasonable cause [31].
Our analysis showed a low resistance rate against amikacin, tobramycin, meropenem, and aztreonam in Proteus isolates. In this extent, high susceptibility to aminoglycoside antibiotics, except gentamicin, carbapenems, and aztreonam, was reported in other studies recently [29, 38–41]. Therefore, these antibiotics could be indicated as the most effective antibiotic agents against pathogenic Proteus isolates.
All the investigated VAGs except atfA were detected in more than 80% of the Proteus isolates. The atfA gene was detected only in 77% of isolates, and zapA (98%) had maximum frequency. Elhoshi et al. have also reported zapA (100%) and atfA (84%) as the VAGs with maximum and minimum frequency, respectively [42].
Data analysis showed that all of the studied VAGs were collectively detected in 47% of isolates. Thus, they could be assigned as highly virulent isolates, and the majority of them (60%) were CA. Therefore, the higher virulence potential of CA isolates in comparison to the HA isolates could be estimated. Similar findings were reported recently regarding other bacteria such as Escherichia coli, Staphylococcus aureus, and Pseudomonas aeruginosa [43–45]. It could logically explain that community isolated bacteria need a richer store of virulence factors for surviving among a very versatile community of bacterial and nonbacterial competitors in the surrounding environment in comparison to the HA isolates that live in an antimicrobial-rich environment of hospitals with a comparable limited number of competitors. Instead, HA isolates acquired more complex antibiotic-resistance mechanisms. Similar estimation has also been reported before by Radera et al. [43].
Association of some of the VAGs with susceptibility against different antibiotic agents was reported regarding Proteus isolates recently [14, 28]. In the present study, logistic regression analysis of data showed that the atfA gene was a predictor of sensitivity to amikacin and tobramycin. The mrpA gene was detected only in P. mirabilis isolates, and the predictive role of the mrpA gene for susceptibility to cefoxitin, norfloxacin, and amoxicillin was also detected by logistic regression analysis. Finally, it could be concluded that atfA and mrpA could be assigned as predictor VAGs for estimating antibiotic susceptibility.
Using qualitative morphologic methods such as the disk diffusion test in order to assess antibiotic susceptibility is popularized because of the simplicity and economic efficiency. But the results are not always trustable. On the other hand, molecular evaluation of antibiotic resistance through screening of the responsible genes, considering the versatile store of ARGs, is usually more expensive and time-consuming in comparison to the phenotypic methods. Therefore, we thought in some cases special VAGs could be introduced as suitable markers for the prognosis of antibiotic susceptibility. According to this, the atfA gene could be introduced as a genetic marker for simple and quick diagnosis of susceptibility against aminoglycoside antibiotics such as amikacin and tobramycin. The mrpA gene could be used similarly for the estimation of susceptibility to cefoxitin, norfloxacin, and amoxicillin in Proteus isolates. However, it has to be evaluated with more detail through future studies.
Based on the study finding, the rsbA gene was detected in all of the CA-isolates. Therefore, it may be introduced for differentiation of HA Proteus isolates from CA isolates. The rsbA gene involved regulation of chemotactic motility and swarming motility, and it also controls expression of other VAGs [46, 47]. Thus, the critical role of the rsbA gene in the natural survival of Proteus species, especially in the surrounding natural environment, and also the commensal behavior of Proteus species in the intestinal tract of animals and humans could be concluded. Therefore, the higher frequency of this gene in CA isolates in comparison to the HA isolates could be logically acceptable.
ERIC-PCR fingerprinting revealed high diversity among studied P. mirabilis isolates. In such a way that, 53 different patterns were detected for 83 isolates. Finally, five patterns were repeated among the studied isolates at 95% genetic distance. Hence, studied isolates were allocated in 5 different clusters (95% confidence interval for genetic distance). ERIC-PCR is known as a relatively simple and not-expensive method that has some limitations such as low reproducibility; however, it has acceptable outcomes, especially in epidemiological studies. In the present study, Proteus isolates were collected from different sources and also diverse samples. Therefore, such high diversity was expected and is acceptable.
5. Conclusion
The present study is one of the few reports that proposed using VAGs for differentiation of Proteus isolates from each other and also antibiotic susceptibility estimation. We suggest that atfA and mrpA genes are marker genes for sensitivity of Proteus species to antibiotic agents. Moreover, the rsbA gene could be introduced as a genetic marker in order to distinguish CA isolates from HA Proteus isolates. Meanwhile, these have to be investigated in a bigger, multicenter bacterial population later. In the present study, only gene presence/absence was screened through PCR without functional assays. Hence, some confirmatory tests such as VAG and ARG expressions in association with antibiotic resistance are proposed through future studies that would confirm our findings. We believe that the conduction of the same studies on more VAGs in association with ARGs could be beneficial in finding a probable direct association between VAGs and antibiotic resistance. It may provide new insights for designing future diagnostic protocols and also treatment strategies. We had some limitations that have to be explained. We do not have access to patient clinical characteristics, which might be helpful in data analysis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Girlich D. Bonnin R. A. Dortet L. Naas T. Genetics of Acquired Antibiotic Resistance Genes in Proteus spp. Frontiers in Microbiology 20201110.3389/fmicb.2020.0025632153540 PMC 7046756 · doi ↗ · pubmed ↗
- 2Rus M. Licker M. Musuroi C. Association of Proteus mirabilis and Providencia stuartii Infections With Diabetes Medicina 2022582 p. 27110.3390/medicina 5802027135208593 PMC 8880118 · doi ↗ · pubmed ↗
- 3Yang J. H. Sheng W. H. Hsueh P. R. Program S. Antimicrobial Susceptibility and Distribution of Extended-Spectrum β-Lactamases, Amp C β-Lactamases and Carbapenemases Among Proteus, Providencia and Morganella Isolated From Global Hospitalised Patients With Intra-Abdominal and Urinary Tract Infections: Results of the Study for Monitoring Antimicrobial Resistance Trends (SMART), 2008-2011 Journal of Global Antimicrobial Resistance 20202239840710.1016/j.jgar.2020.04.01132311502 · doi ↗ · pubmed ↗
- 4Wasfi R. Hamed S. M. Amer M. A. Fahmy L. I. Proteus mirabilis Biofilm: Development and Therapeutic Strategies Frontiers in Cellular and Infection Microbiology 20201010.3389/fcimb.2020.0041432923408 PMC 7456845 · doi ↗ · pubmed ↗
- 5Jansen A. M. Lockatell C. V. Johnson D. E. Mobley H. L. Visualization of Proteus mirabilis Morphotypes in the Urinary Tract: The Elongated Swarmer Cell Is Rarely Observed in Ascending Urinary Tract Infection Infection and Immunity 20037163607361310.1128/IAI.71.6.3607-3613.20032-s 2.0-003844303912761147 PMC 155743 · doi ↗ · pubmed ↗
- 6Mirzaei A. Habibi M. Bouzari S. Asadi Karam M. R. Characterization of Antibiotic-Susceptibility Patterns, Virulence Factor Profiles and Clonal Relatedness in Proteus mirabilis Isolates From Patients With Urinary Tract Infection in Iran Infection and Drug Resistance 2019123967397910.2147/IDR.S 23030331920349 PMC 6938180 · doi ↗ · pubmed ↗
- 7Pearson M. M. Yep A. Smith S. N. Mobley H. L. Transcriptome of Proteus mirabilis in the Murine Urinary Tract: Virulence and Nitrogen Assimilation Gene Expression Infection and Immunity 20117972619263110.1128/IAI.05152-112-s 2.0-7995947304221505083 PMC 3191972 · doi ↗ · pubmed ↗
- 8Nielubowicz G. R. Mobley H. L. Host-Pathogen Interactions in Urinary Tract Infection Nature Reviews Urology 20107843044110.1038/nrurol.2010.1012-s 2.0-7795565459120647992 · doi ↗ · pubmed ↗