Duplicated genes find their space: Spatial transcriptomics illuminates evolutionary fates
Min-Yao Jhu, Fabian van Beveren, Bruno Guillotin

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
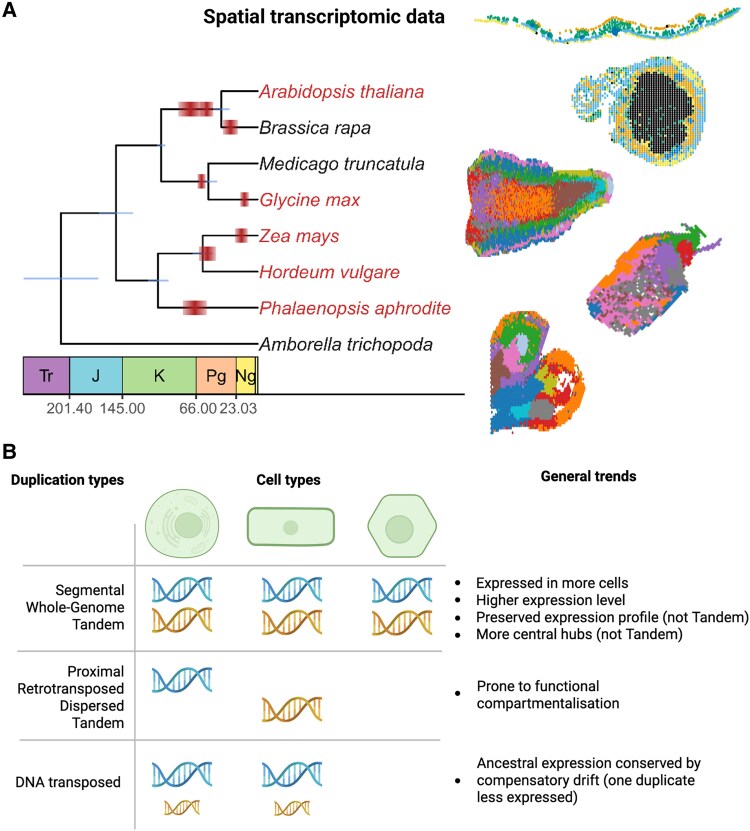 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSingle-cell and spatial transcriptomics · Genomics and Phylogenetic Studies · Chromosomal and Genetic Variations
Gene and genome duplications are fundamental forces driving plant evolution, leading to relaxed selection and expanding genetic toolkits (Van de Peer et al. 2017; Almeida-Silva and Van de Peer 2023). However, the subsequent fate of these duplicates—whether they retain redundant functions, divide tasks (subfunctionalization), or acquire new roles (neofunctionalization)—largely depends on how they originated (Birchler and Yang 2022). While previous evolutionary studies relied primarily on bulk RNA sequencing, which aggregates expression profiles across entire tissues, recent research published by Almeida-Silva and Van de Peer uses spatial transcriptomics across 5 diverse plant species (Arabidopsis thaliana, soybean, orchid, maize, and barley) to reveal the evolution of gene expression divergence at the cell-type level (Figure A). This provides a spatially resolved view of how the mode of duplication impacts the expression of duplicated genes across angiosperms.
The authors confirm a major paradigm: duplication mechanisms that preserve the cis-regulatory landscape generally yield paralogs with more preserved expression profiles (Figure B). Genes originating from segmental or whole-genome duplication (SD/WGD) and tandem duplication (TD) events often exhibit greater expression levels and are expressed in more cell types compared with genes derived from dispersed duplications. This preservation is hypothesized to be linked to the retention of ancestral transcription factor binding sites in promoters and enhancers. These large-scale duplicates are also found to be overrepresented in spatially variable gene sets, indicating variable expression across tissue compartments. Furthermore, SD/WGD-derived genes frequently hold central roles as hubs within coexpression networks, a finding consistent with the preferential retention of essential, dosage-sensitive genes following SD/WGD events (Jeong et al. 2001).
When classifying the eventual expression outcome of these duplicates, a clear separation emerges based on the scale of the duplication. Paralogs from large-scale duplications (SD/WGD) and small-scale DNA-transposed duplications (dTRD) typically display redundant or overlapping expression profiles across cell types more frequently than expected, suggesting functional redundancy or subfunctionalization in certain species. Conversely, other small-scale duplicates, including TD, proximal duplications, retrotransposed duplications, and dispersed duplications, often diverge asymmetrically. One gene maintains broad expression while the other specializes in a few cell types, a hallmark of functional compartmentalization (Figure B).
Crucially, the high-resolution data unveiled a fascinating mechanistic exception within the small-scale group: dTRD. Although dTRD genes do not preserve cis-regulatory elements to the same extent as SD/WGD or TD genes, they are nonetheless often overrepresented in pairs displaying expression-level redundancy (Figure B). The researchers determined that dTRD redundancy is not due to regulatory preservation but rather is better explained by “compensatory drift”: one copy evolves toward lower expression while the paralog evolves toward higher expression, thereby maintaining the overall total ancestral expression level (Iohannes and Jackson 2023). This leads to dTRD pairs displaying significantly greater absolute differences in expression levels between paralogs across cell types compared with other modes of duplication.
The study also underscores that the impact of the duplication mode is not static: time is a crucial factor. While young duplicates (less than 20 million years ago) clearly show lower divergence when resulting from mechanisms that preserve regulatory landscapes (SD, TD, proximal duplications), these differences based on duplication mode disappear or fade over time in more ancient duplicates.
Finally, gene function imposes strong constraints on expression divergence. Dosage-sensitive genes involved in basic cellular processes (like chromatin assembly or transcription regulation) display highly preserved expression profiles, likely due to selection pressures maintaining stoichiometric balance (Birchler and Veitia 2012). Meanwhile, genes involved in specialized processes, such as flowering and phytohormone biosynthesis, diverge much more rapidly.
Collectively, by leveraging spatial transcriptomics, this research not only confirms that established theories of gene expression evolution following duplication hold true within tissue compartments but also provides mechanistic insights, like compensatory drift, which explain divergence outcomes when traditional cis-regulatory preservation is lacking. By mapping expression divergence within tissues and across cell types, they reveal how the combination of duplication mode, gene function, and evolutionary time impacts the transcriptional fate of gene duplicates and by extension shapes plant genome evolution and cell identity. While these overarching evolutionary outcomes and patterns are broadly conserved, the degree of divergence and functional specialization can vary among species, as also shown in this work, highlighting a potential future direction to uncover lineage-specific evolutionary trajectories.
Recent related articles in The Plant Cell:
Birchler and Yang (2022) discussed the multiple evolutionary fates of duplicated genes—including deletion, subfunctionalization, neofunctionalization, dosage balance, and neutral variation—highlighting how duplication mechanisms and functional constraints shape plant genome evolution. Van de Peer et al. (2021) reviewed how whole-genome duplications and polyploidy contribute to plant survival and diversification, emphasizing their role in adaptation to abiotic and biotic stress. Li et al. (2016) showed that core gene families display consistent duplicability across angiosperms, with single-copy genes linked to genome stability and multicopy genes to signaling and metabolism.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Almeida-Silva F, Van de Peer Y. Whole-genome duplications and the long-term evolution of gene regulatory networks in angiosperms. Mol Biol Evol. 2023:40(7):msad 141. 10.1093/molbev/msad 14137405949 PMC 10321489 · doi ↗ · pubmed ↗
- 2Almeida-Silva F, Van de Peer Y. Gene expression divergence following gene and genome duplications in spatially resolved plant transcriptomes. Plant Cell. 2025:37(10):koaf 243. 10.1093/plcell/koaf 24341092097 · doi ↗ · pubmed ↗
- 3Birchler JA, Veitia RA. Gene balance hypothesis: connecting issues of dosage sensitivity across biological disciplines. Proc Natl Acad Sci U S A. 2012:109(37):14746–14753. 10.1073/pnas.120772610922908297 PMC 3443177 · doi ↗ · pubmed ↗
- 4Birchler JA, Yang H. The multiple fates of gene duplications: deletion, hypofunctionalization, subfunctionalization, neofunctionalization, dosage balance constraints, and neutral variation. Plant Cell. 2022:34(7):2466–2474. 10.1093/plcell/koac 07635253876 PMC 9252495 · doi ↗ · pubmed ↗
- 5Chen H, Almeida-Silva F, Logghe G, Maere S, Bonte D, Van de Peer Y. The Rise of Polyploids During Environmental Upheaval. bio Rxiv 2025. 10.1101/2024.11.22.624806. Preprint. · doi ↗
- 6Iohannes SD, Jackson D. Tackling redundancy: genetic mechanisms underlying paralog compensation in plants. New Phytol. 2023:240(4):1381–1389. 10.1111/nph.1926737724752 · doi ↗ · pubmed ↗
- 7Jeong H, Mason SP, Barabási A-L, Oltvai ZN. Lethality and centrality in protein networks. Nature. 2001:411(6833):41–42. 10.1038/3507513811333967 · doi ↗ · pubmed ↗
- 8Li Z, Defoort J, Tasdighian S, Maere S, Van de Peer Y, De Smet R. Gene duplicability of core genes is highly consistent across all angiosperms. Plant Cell. 2016:28(2):326–344. 10.1105/tpc.15.0087726744215 PMC 4790876 · doi ↗ · pubmed ↗