Ruminococcus torques Administration Modestly Alleviates Dietary Selenium Deficiency-Induced Glucose Intolerance in Mature Female Mice
Ying-Chen Huang, Wen-Hsing Cheng

TL;DR
Giving Ruminococcus torques to mice with low selenium diets slightly improves their glucose intolerance, but not insulin resistance.
Contribution
The study reveals a novel interaction between Ruminococcus torques and dietary selenium in modulating glucose metabolism in mice.
Findings
R. torques administration modestly alleviated glucose intolerance in selenium-deficient mice.
Selenium deficiency increased cecal E. coli abundance, which was further elevated by R. torques treatment.
R. torques gavage altered liver GPX1 and SELENOP protein levels in selenium-deficient mice.
Abstract
We previously found that dietary selenium (Se) deficiency and age increase the fecal abundance of Lachnospiraceae in aged telomere-humanized mice in a sexually dimorphic manner. Although Lachnospiraceae are key contributors of short-chain fatty acids to the host, different taxa within this family exert distinct effects on host physiology. Among them, Ruminococcus torques has been associated with type 2 diabetes. In the present study, we aimed to determine whether, and how, R. torques interacts with dietary Se to influence type 2 diabetes-like symptoms. Sixteen weaning female C57BL/6J mice were fed either a Se-adequate or Se-deficient diet for 26 weeks. From weeks 21 to 25, half of the mice in each dietary group received daily oral gavage of R. torques (2 × 108 CFU in 0.2 mL). All mice were euthanized at week 26. Dietary Se deficiency induced glucose intolerance (13%) and insulin…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
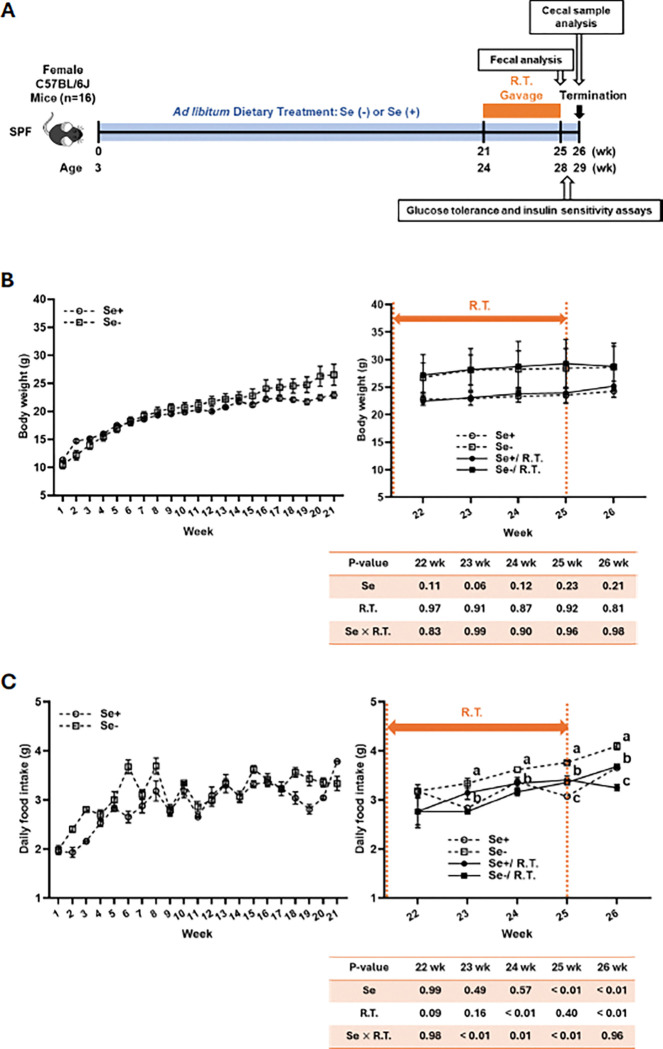 Figure 1
Figure 1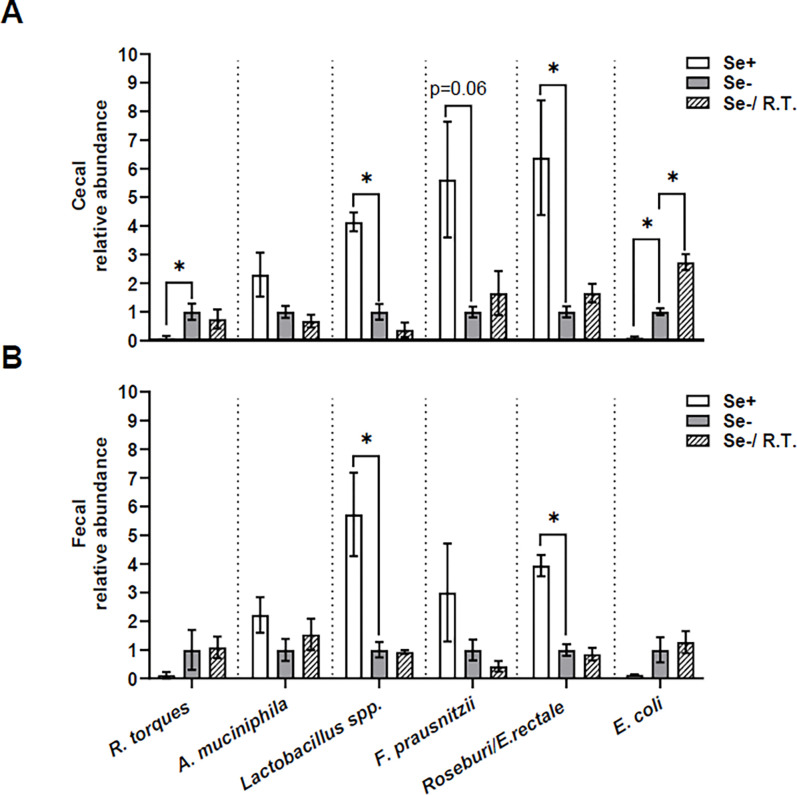 Figure 2
Figure 2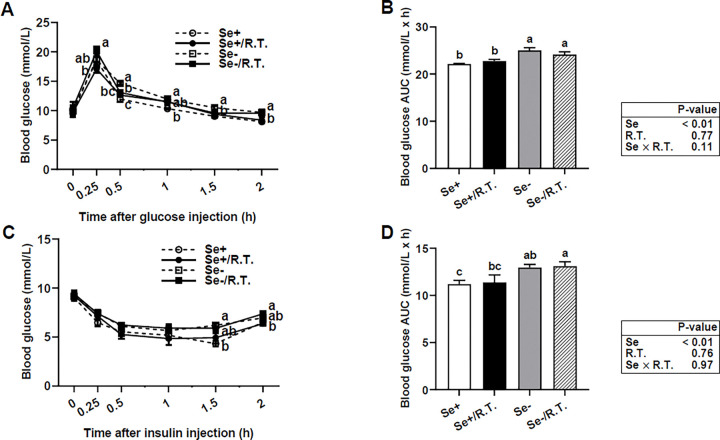 Figure 3
Figure 3 Figure 4
Figure 4- —NIH
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSelenium in Biological Systems · Rabbits: Nutrition, Reproduction, Health · Moringa oleifera research and applications
Introduction
The Lachnospiraceae are a family of anaerobic bacteria within the Clostridiales order of the Firmicutes phylum, and include species previously classified as part of Clostridium cluster XIVa [1, 2]. This family is abundant and dominant in the unperturbed adult gut microbiota [3, 4]. However, only 25% of the genomes are closely related (> 97% similarity) to known species, and 28% do not match any records of existing databases [3]. Interestingly, 19% of sequences are closely related to recently isolated butyrate-producing bacteria from Clostridium clusters XIVa and IV, while 18% are related to Ruminococcus obeum and Ruminococcus torques, which are members of XIVa [3]. Ruminococcus is currently considered a polyphyletic genus, with species distributed across two distinct families: the Ruminococcaceae and the Lachnospiraceae [5]. Based on phenotypic traits, 16S rRNA gene sequence similarity [6], phylogenetic analysis, G + C content of genomic sequence, and DNA-DNA hybridization studies, the species R. torques, R. obeum, R. lactaris, R. gnavus, and R. gauvreauii are classified within the Lachnospiraceae family [7].
Representing a core component of the gut microbiota, Lachnospiraceae colonize the intestinal lumen from birth, with their species richness and relative abundance increasing with age. Lachnospiraceae have been implicated in obesity and diabetes in both humans and mouse models [8–14]; however, their specific role in type 2 diabetes remains unclear. We have previously shown that long-term dietary selenium (Se) deficiency induces type 2 diabetes-like symptoms in both male and female telomere-humanized mice [15], and increases the fecal abundance of Lachnospiraceae at 18 months of age in both sexes, but only in females at 24 months, based on 16 rRNA gene sequence analyses [16]. In contrast, Akkermansia muciniphila, a mucin-degrading bacterium that comprises 3–5% of the microbial community in healthy individuals [17], shows the most pronounced age-associated enrichment in response to dietary Se deficiency, but only in male telomere-humanized mice. Within the Lachnospiraceae family, R. torques is another mucin-degrading bacterium whose abundance is positively associated with irritable bowel syndrome in humans [18, 19]. Because oral gavage of A. muciniphila alleviates type 2 diabetes-like symptoms in Se-deficient male mice, and these findings collectively suggest that mucin-degrading bacteria may play a critical role in modulating inflammatory responses at the gut mucosal surface [16]. Therefore, we aimed to investigate the effects of oral administration of R. torques on type 2 diabetes-like symptoms induced by dietary Se deficiency, as well as on symbiotic changes involving other mucin-degrading or short-chain fatty acid-producing bacteria in mature female mice. These results may also provide insights into sexually dimorphic responses in Se deficiency-induced type 2 diabetes and gut microbiota alterations.
Materials and Methods
Culture of Ruminococcus torques
R. torques (ATCC BAA-2281) were cultured under strict anaerobic conditions using the Anoxomat III Jar system (Advanced Instruments, Norwood, MA) in modified reinforced clostridial medium following ATCC protocol. Cultures were washed and concentrated anaerobically in sterile, anaerobic PBS containing 25% (vol/vol) glycerol to a final concentration of 1 × 10^10^ CFU/mL, then immediately frozen and stored at −80°C. Before oral administration, stocks were thawed and diluted anaerobically in sterile PBS with 2.5% glycerol to a final concentration of 1 × 10^9^ CFU/mL.
Mice, diets, and treatment
As shown in Fig. 1A, sixteen weaning female C57BL/6J mice were housed under conventional specific pathogen-free (SPF) conditions and fed either a Se-deficient or Se-adequate torula yeast-based purified diet for 26 weeks, as previously described previously [16]. At week 21, mice (n = 4 per group) received a daily oral gavage of either 2 × 10^8^ CFU live R. torques or a control vehicle (2.5% glycerol in 200 μL) for 4 weeks. Mice were handled aseptically in a controlled environment with 12-hour light/dark cycle (lights on from 6 p.m. to 6 a.m.), with ad libitum access to food and water. Body weight and food intake were monitored weekly. Fresh fecal samples were collected before and after R. torques treatment, snap-frozen in liquid nitrogen, and stored at −80°C. Two days after the insulin sensitivity assay, mice were fasted for 6 hours, anesthetized with carbon dioxide, and euthanized by cardiac exsanguination. Liver, skeletal muscle, and cecal contents were collected, rapidly frozen in liquid nitrogen, and stored at −80°C for further analyses. All procedures were approved by the Institutional Animal Care and Use Committee of Mississippi State University.
Glucose tolerance and insulin sensitivity
At weeks 21 and 26 of dietary intervention, mice fasted for 8 hours were intraperitoneally injected with glucose (1 g/kg body weight) or insulin (0.25 U/kg body weight) (Sigma Aldrich, St. Louis, MO). Blood glucose concentrations were measured using a glucose meter (Bayer Contour Next EZ, Ascensia Diabetes Care US, Inc., Parsippany, NJ) from a drop of tail vein samples collected at 0 (baseline), 0.25, 0.5, 1, 1.5, 2 hours post-injection. Insulin sensitivity was assessed 2 days after glucose tolerance testing. The area under the curve (AUC) was calculated to quantify glucose and insulin responses.
Bacterial genomic DNA extraction and Quantitative PCR (qPCR) analysis
Bacterial DNA was extracted from cecal contents and feces using the QIAamp PowerFecal Pro DNA Kit (#51804, QIAGEN, Germantown, MD) following the manufacturer’s instructions. Universal primers targeting the V4 region of the bacterial 16S rRNA gene were used for amplification (primer sequences listed in Supplemental Table 1). qPCR was performed in 10 μL reactions using PowerUp^™^ SYBR^™^ Green Master Mix (#A25741, Applied Biosystems, Waltham, MA) on a QuantStudio 5 Real-Time PCR System (#A34322, Applied Biosystems) with the following thermal profile: 95°C for 2 min, followed by 40 cycles of 95°C for 5 sec and 60°C for 30 sec. Total bacterial load in fecal samples was quantified using the ΔCT method. Due to normalization of ΔCT values by subtraction against the control group, two-way ANOVA was not appropriate for analyzing ΔΔCT data, as a designated control could not be applied across two factors. Instead, unpaired t-tests were used for statistical analysis.
Immunoblotting
Tissues were homogenized in RIPA lysis buffer with protease inhibitors (# sc-24948, Santa Cruz Biotech, Dallas, TX) and centrifuged at 12,000 × g for 10 min at 4°C. Supernatants (30 μg protein per lane) were separated by 14% SDS-PAGE, then transferred to polyvinylidene difluoride membranes. Membranes were incubated overnight at 4°C with primary antibodies (listed in Supplemental Table 2), followed by HRP-conjugated secondary antibodies for 2 hours at room temperature. Signals were developed using Clarity Western ECL substrate, and images were captured and quantified using a Chemidoc-XS system with the volume tool in Image Lab Software (Bio-Rad Lab, Hercules, CA). Protein levels were normalized to albumin, β-tubulin, or total AKT.
Statistical analysis
Data are presented as means ± SEM. Most datasets were analyzed by two-way ANOVA followed by Tukey’s post hoc test, except for qPCR-based assays, which were evaluated using unpaired t-tests. Statistical analyses were performed using SAS (version 9.4) and GraphPad Prism (version 8.0). A significance level of α = 0.05 was used for all tests.
Results
Body weight and food intake
Weaning female C57BL/6J mice exhibited steady increases in body weight (103–153%; P < 0.05; Fig. 1B) and food intake (56–70%; P < 0.05; Fig. 1C) over the 21-week dietary intervention. In mice receiving control oral gavage, dietary Se deficiency increased food intake (8–18%; P < 0.05) but had no effect on body weight. Daily oral gavage with R. torques resulted in a deduction in food intake in Se-deficient mice at weeks 23–26 (10–17%; P < 0.05), without affecting body weight. In R. torques-gavaged mice, dietary Se deficiency increased food intake at week 26 (13%; P < 0.05) but did not alter body weight.
Changes in the abundance of specific genera by dietary Se deficiency and R. torques oral gavage in mice
In Se-adequate mice, oral gavage with R. torques was associated with apparent increases (3- to 5-fold) in the abundances of R. torques, alongside decreases (52–82%; P < 0.05) in Lactobacillus spp. and Roseburia/E. rectale in the cecal and fecal contents (Supplemental Fig. 1). In mice receiving control oral gavage, dietary Se deficiency increased (P ≤ 0.06) the abundances of R. torques (10.9-fold) and E. coli (9.5-fold), while decreasing that of Lactobacillus spp. (3.1-fold), F. prausnitzii (4.6-fold), and Roseburia/E. rectale (5.3-fold) in cecal content (Fig. 2A). A similar trend was observed in fecal content (Fig. 2B), but only Lactobacillus spp. (4.7-fold) and Roseburia/E. rectale (2.9-fold) reached statistical significance (P < 0.05). Oral gavage with R. torques had no effect on these Se deficiency-induced alterations, except for a 1.7-fold increase (P < 0.05) in cecal E. coli abundance. In mice receiving oral gavage with R. torques, dietary Se deficiency increased the relative abundance of E. coli in both cecal and fecal contents (4- to10-fold; P < 0.05) and decreased that of fecal Roseburia/E. rectale by 54% (P < 0.05), but not in other taxa (supplemental Fig. 2).
Modest alleviation by R. torques of dietary Se deficiency-induced glucose intolerance, but not insulin insensitivity, in conventional mice
Se-deficient female mice at ~ 7 months of age exhibited glucose intolerance (Fig. 3A and B) and insulin resistance (Fig. 3C and D) over a 2-hour period following injections. However, fasting blood glucose concentrations were not affected by dietary Se deficiency or by R. torques oral gavage (Fig. 3A and C). While oral gavage with R. torques modestly alleviated (11% improvement; P < 0.05) Se deficiency-induced glucose intolerance at 0.5 h post-injection (Fig. 3A), it had no significant effect on overall glucose tolerance or insulin sensitivity across the entire time course in either the Se-deficient or Se-adequate mice.
R. torques oral gavage and dietary Se deficiency differentially affect selenoprotein expression in muscle and liver
We next assessed body Se status in the liver and skeletal muscle by evaluating protein expression levels of selected selenoproteins. Western blot analysis revealed that dietary Se deficiency reduced (P < 0.05) the protein levels of GPX1 by 41% and SELENOW by 89% in muscle; whereas SELENOH and SELENOP expression remained unaffected (Fig. 5A). In the liver (Fig. 5B), Se deficiency decreased (P < 0.05) protein levels of SELENOP by 50%, GPX1 by 86%, SELENOH by 56%, and SELENOW by 88%. Notably, the reduction in hepatic GPX1 was partially reversed by R. torques oral gavage (1.6-fold increase; P < 0.05). In Se-adequate mice, oral gavage of R. torques reduced (P < 0.05) hepatic levels of both SELENOP and GPX1 by 27%, but had no effect on SELENOH or SELENOW in the liver, nor on any selenoprotein expression in muscle.
Discussion
We have previously shown that long-term dietary Se deficiency induces type 2 diabetes-like symptoms in telomere-humanized mice aged 12 and 18 months in both sexes [15], as well as middle-aged and mature wild-type male mice [16, 20]. Here, compared to wild-type male mice aged 4–7 months [16], dietary Se deficiency in age-matched females induces glucose intolerance and insulin resistance to a lesser extent. Consistent with our findings, there is evidence that females are generally more insulin-sensitive than males [21, 22], and women have decreased susceptibility to fatty acid–induced peripheral insulin resistance [23]. In rodent models, males typically exhibit more pronounced diabetes symptoms than females [24–27]. These sex-related differences in type 2 diabetes-like symptoms may be partially attributed to the actions of estrogen and testosterone. For example, decreases in estrogen and increases in testosterone levels during menopause are associated with a loss of subcutaneous fat, a gain of visceral fat, and increased insulin resistance [28]. Indeed, we have previously shown that Se status under Se deficiency is tissue-specific and dependent on sex and age [29].
In contrast to A. muciniphila [16], oral gavage with R. torques only modestly alleviates Se deficiency-induced type 2 diabetes symptoms. An intriguing question arises: why do these two mucin-degrading bacteria have opposing impacts on Se deficiency-induced metabolic symptoms? Although some mucin-degrading bacteria, such as A. muciniphila, are associated with health benefits, their close proximity to the intestinal epithelium may pose risks to host cells by compromising the gut barrier. It has been suggested that a reduction in cecal R. torques abundance in response to Lactobacillus acidophilus treatment may improve reproductive performance of broiler chickens by reducing the incidence of pasty vent, a stress-induced condition characterized by dry, cake-like droppings around the vent of baby chicks [30]. Supporting this observation, mucin degradation has been proposed to be pathogenic, as the loss of the protective mucus layer may increase the exposure of gastrointestinal epithelial cells to pathogens [31]. Moreover, the abundance of total mucosa-associated bacterial 16S rRNA genes is elevated in inflammatory bowel disorder, suggesting an increased availability of digestible endogenous mucus that may, in turn, sustain the growth of non-mucolytic mucosa-associated bacteria [32]. These findings highlight the need for further investigation into the role of mucin in shaping microbial community dynamics and host–microbe interactions.
R. torques has been reported to be correlated with markers of insulin resistance in humans [33]. In our study, R. torques abundance was increased in mice fed a Se-deficient diet compared to those on a Se-adequate diet. However, our results indicate that oral administration of R. torques modestly improves glucose intolerance but has no effect on insulin resistance in Se-deficiency female mice. We speculate that 1) female mice may exhibit greater baseline insulin sensitivity, potentially masking any mild effects of R. torques; 2) interactions between R. torques and host selenoproteins may counterbalance each other’s influence on type-2 diabetes pathogenesis; 3) the effects of R. torques on Se deficiency-induced type 2 diabetes may be age-dependent, becoming apparent only in middle-aged or older mice, but not in younger mice.
Although a recent study using the type strain R. torques ATCC 27756 (BSL-1) demonstrated a pronounced alleviation of glucose intolerance in obese mice via oral gavage [2], we employed a non-type strain, R. torques ATCC BAA-2281 (BSL-2), and observed only a modest improvement in Se deficiency-induced glucose intolerance. In addition to the fact that ATCC BAA-2281 has been minimally characterized, we selected this strain to test its effect on type 2 diabetes-like symptoms because it is deposited in ATCC as a Lachnospiraceae bacterium. We have previously shown that dietary Se deficiency enriches the relative abundance of Lachnospiraceae in aged telomere-humanized mice in a sexually dimorphic manner [16]. To better understand the differing efficacy of these two R. torques strains in modulating glucose tolerance, future studies should consider factors such as host obesity status, strain-level genetic variation, and the underlying etiology of type 2 diabetes.
Other gut bacteria may respond to R. torques oral gavage and influence outcomes related to glucose intolerance and insulin resistance in Se-deficient mice. Indeed, Se deficiency in females decreases the abundance of Lactobacillus spp., F. prausnitzii, and Roseburia spp./E. rectale, while increasing the abundance of R. torques and E. coli in the cecal content. However, R. torques oral gavage results in increased E. coli abundance in Se-deficient mice and decreased Lactobacillus spp. and Roseburia spp./E. rectal abundance in Se-adequate mice. These findings are in contrast to those observed with A. muciniphila oral gavage, suggesting that these two mucin-degrading bacteria impact gut homeostasis and host health through distinct mechanisms.
Altogether, our results show that the administration of the R. torques ATCC BAA-2281 strain only modestly alleviates glucose intolerance in Se-deficient female mice, and that changes in five other bacterial taxa display patterns distinct from those observed with A. muciniphila oral gavage in male mice [16]. Further studies are needed to deepen our understanding and provide mechanistic insight into how R. torques influences Se status and type 2 diabetes in the host, as well as how Se may prevent the early onset of type 2 diabetes in a sexually dimorphic manner.
Supplementary Material
Supplementary Files
This is a list of supplementary files associated with this preprint. Click to download.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Vacca M, Celano G, Calabrese FM, Portincasa P, Gobbetti M, De Angelis M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms. 2020;8(4). doi: 10.3390/microorganisms 8040573. · doi ↗
- 2Fan Y, Lyu L, Vazquez-Uribe R, Zhang W, Bongers M, Koulouktsis A, Polypeptides synthesized by common bacteria in the human gut improve rodent metabolism. Nature Microbiology. 2025;10(8):1918–39. doi: 10.1038/s 41564-025-02064-x. · doi ↗
- 3Hold GL, Pryde SE, Russell VJ, Furrie E, Flint HJ. Assessment of microbial diversity in human colonic samples by 16S r DNA sequence analysis. FEMS microbiology ecology. 2002;39(1):33–9. doi: 10.1016/S 0168-6496(01)00190-8.19709182 · doi ↗ · pubmed ↗
- 4Lopetuso LR, Scaldaferri F, Petito V, Gasbarrini A. Commensal Clostridia: leading players in the maintenance of gut homeostasis. Gut Pathogens. 2013;5(1):23. doi: 10.1186/1757-4749-5-23.23941657 PMC 3751348 · doi ↗ · pubmed ↗
- 5Rainey FA, Janssen PH. Phylogenetic analysis by 16S ribosomal DNA sequence comparison reveals two unrelated groups of species within the genus Ruminococcus. FEMS microbiology letters. 1995;129(1):69–73. doi: 10.1016/0378-1097(95)00138-U.7781992 · doi ↗ · pubmed ↗
- 6Kim M, Oh HS, Park SC, Chun J. Towards a taxonomic coherence between average nucleotide identity and 16S r RNA gene sequence similarity for species demarcation of prokaryotes. International journal of systematic and evolutionary microbiology. 2014;64(Pt 2):346–51. doi: 10.1099/ijs.0.059774-0.24505072 · doi ↗ · pubmed ↗
- 7Togo AH, Diop A, Bittar F, Maraninchi M, Valero R, Armstrong N, Description of Mediterraneibacter massiliensis, gen. nov., sp. nov., a new genus isolated from the gut microbiota of an obese patient and reclassification of Ruminococcus faecis, Ruminococcus lactaris, Ruminococcus torques, Ruminococcus gnavus and Clostridium glycyrrhizinilyticum as Mediterraneibacter faecis comb. nov., Mediterraneibacter lactaris comb. nov., Mediterraneibacter torques comb. nov., Mediterraneibacter gnavus comb. nov. and · doi ↗ · pubmed ↗
- 8Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(22):9066–71. doi: 10.1073/pnas.1219451110.23671105 PMC 3670398 · doi ↗ · pubmed ↗