Tonoplast sucrose transporter SUT4‐dependent sugar partitioning modulates phenological transitions and reproductive success in poplar
Trevor T. Tuma, Holly A. McInnes, HongDuyen Pham, William P. Bewg, Michihito Deguchi, Ran Zhou, Samantha M. Surber, Brent Lieb, Anna Lipzen, Kerrie W. Barry, Daniela L. Weber Wyneken, Anne E. Harman‐Ware, Joseph Dahlen, Scott A. Harding, Chung‐Jui Tsai

TL;DR
A study shows that a specific sugar transporter in poplar trees, SUT4, is crucial for seasonal growth and reproduction, and its absence disrupts timing and fertility.
Contribution
The study reveals SUT4's role in sugar partitioning and seasonal development under field conditions, not just greenhouse settings.
Findings
Loss of SUT4 causes earlier leaf senescence, delayed spring growth, and reduced stem growth in poplar.
SUT4 mutants produce sterile ovules despite normal-looking flowers and show disrupted sugar and raffinose dynamics.
Mutants exhibit elevated stress and downregulated circadian clock and proanthocyanidin biosynthesis genes.
Abstract
Climate uncertainty is intensifying the need for greater plasticity in carbohydrate reserve utilization to support winter survival and spring growth in woody perennials. In poplar, the single‐copy SUT4, which encodes a tonoplast‐localized sucrose transporter, and the SUT5/SUT6 genome duplicates, which encode plasma membrane‐localized transporters, are expressed year‐round, with SUT4 showing the highest expression during cool seasons. Given its role in vacuolar sucrose efflux and winter‐predominant expression, SUT4 may play a key role in modulating seasonal carbohydrate dynamics. While SUT4‐knockdown and knockout effects have been studied under greenhouse conditions, their impact under field conditions remains unexplored. Here, we report a field‐based study comparing CRISPR knockout mutants of winter‐expressed SUT4 and SUT5/SUT6 in Populus tremula × alba. We show that sut4, but not…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
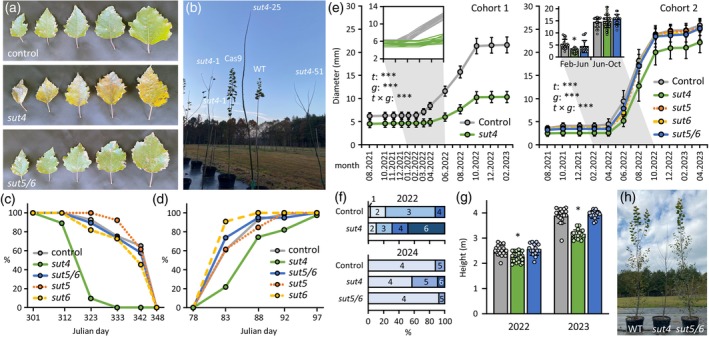 Figure 1
Figure 1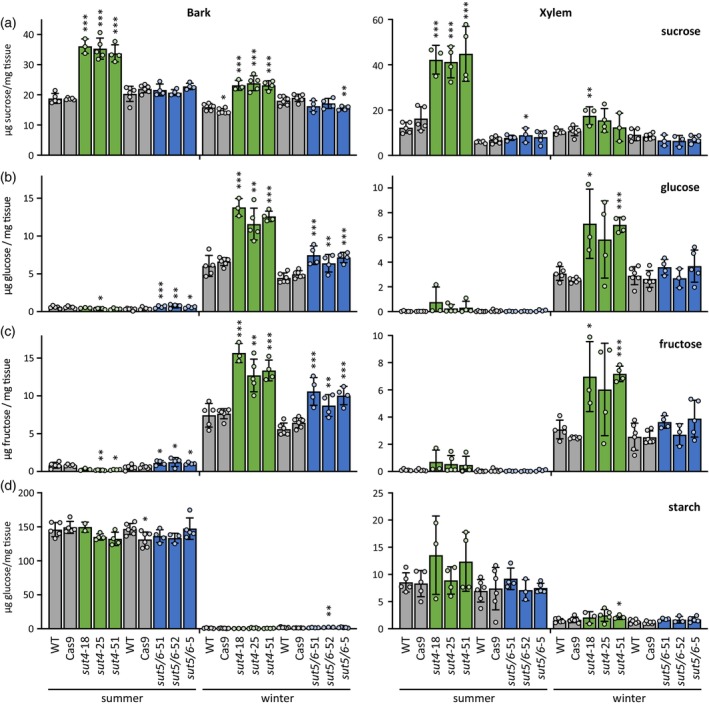 Figure 2
Figure 2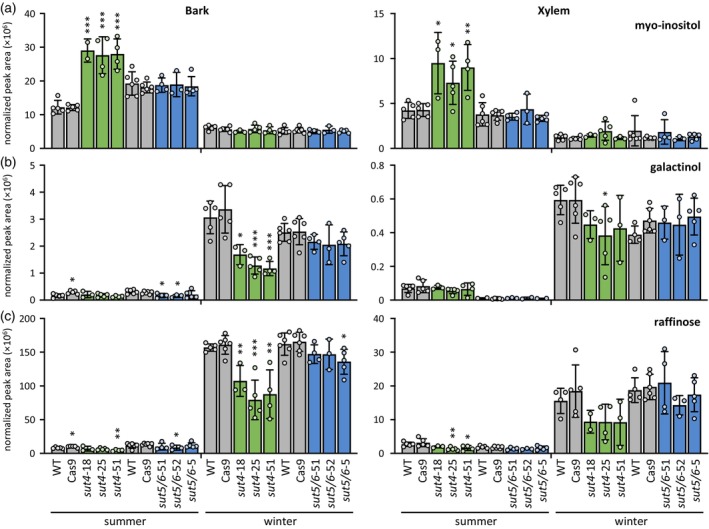 Figure 3
Figure 3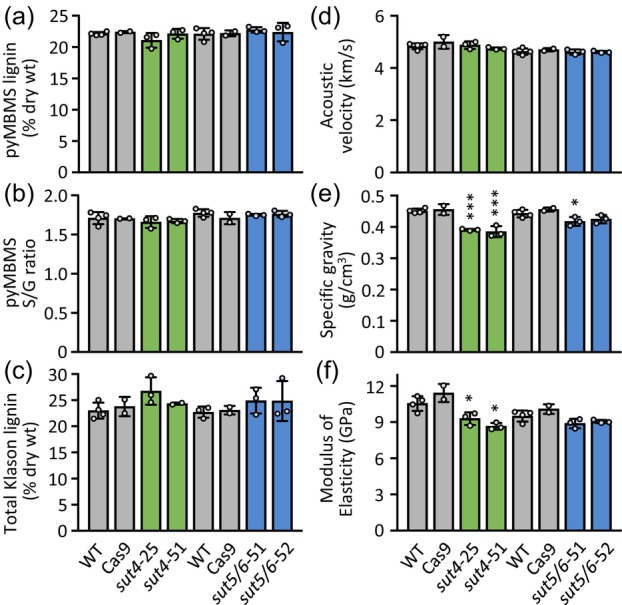 Figure 4
Figure 4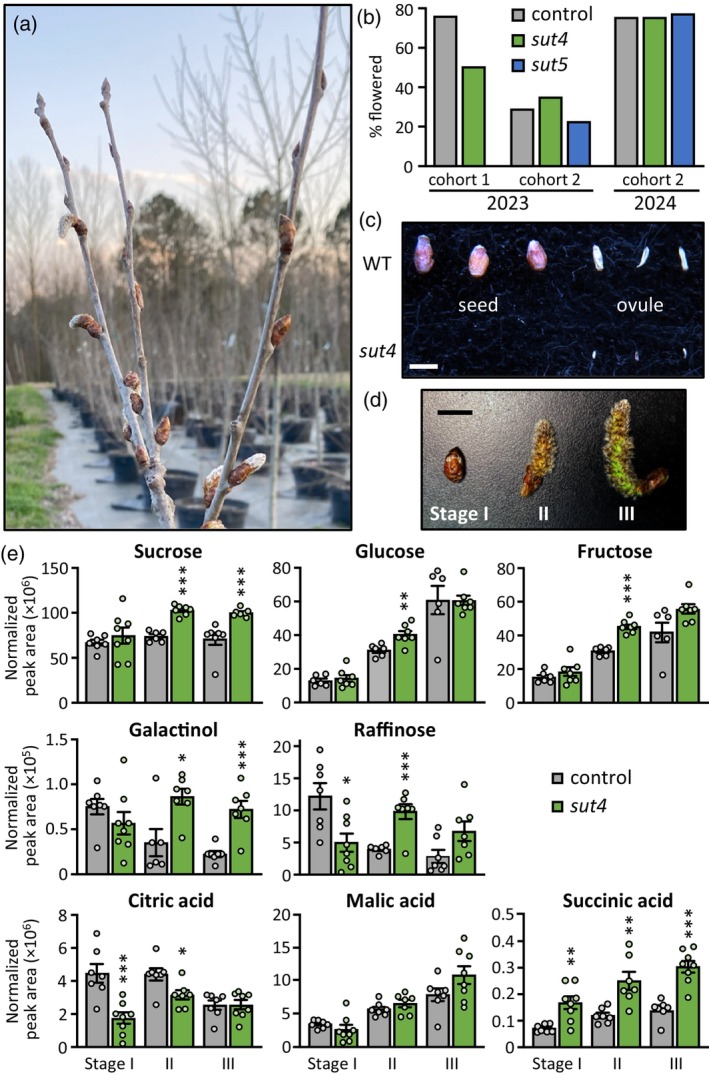 Figure 5
Figure 5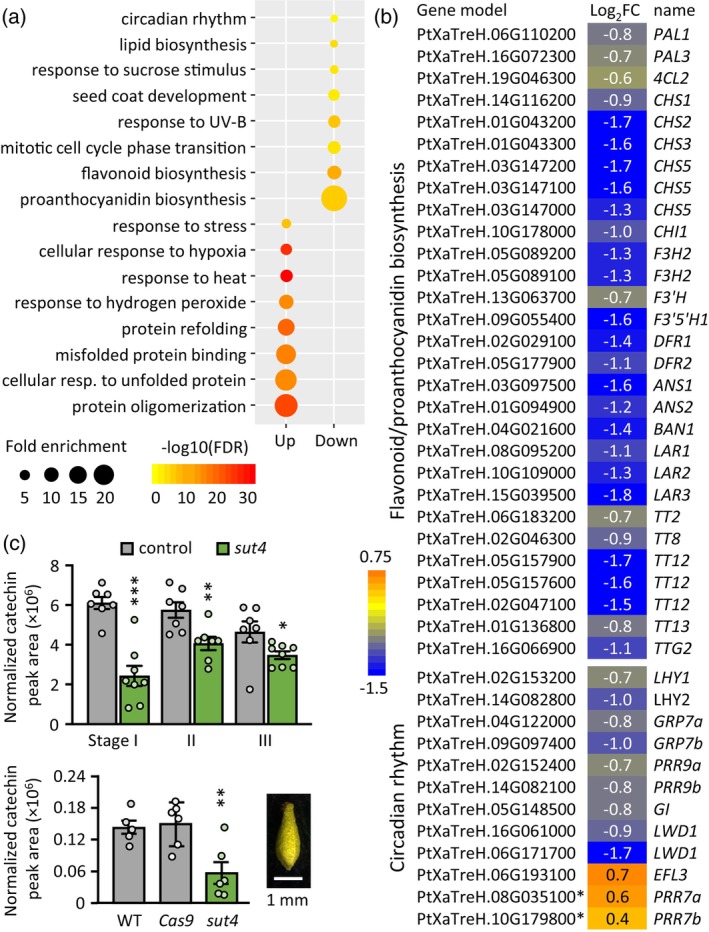 Figure 6
Figure 6- —National Science Foundation10.13039/100000001
- —U.S. Department of Energy10.13039/100000015
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant nutrient uptake and metabolism · Plant Molecular Biology Research · Plant Gene Expression Analysis
INTRODUCTION
Woody perennials in temperate regions undergo an annual cycle of growth, dormancy, and resumption in response to seasonal cues such as temperature and day length. Following autumnal leaf drop, these species rely on stored carbohydrate reserves to support critical transitions through dormancy, floral development, flowering, and leaf emergence in the spring (Cook et al., 2012; Rohde & Bhalerao, 2007; Savage & Chuine, 2021). Rising temperatures and increasing climate variability are exacerbating uncertainty in the timing, trajectory, and phenological consequences of seasonal weather transitions (Calinger & Curtis, 2023; Rohde et al., 2011; Tanino et al., 2010). For perennial plants, this uncertainty underscores the growing importance of plasticity in carbohydrate resorption, storage, and remobilization for long‐term resilience.
An important player in this context may be the tonoplast sucrose transporter SUT4, which mediates sucrose efflux from the vacuole into the cytosol (Payyavula et al., 2011; Reinders et al., 2008; Schneider et al., 2012). SUT4 expression is sensitive to drought and temperature stress, and its regulation by hormone and light quality signals positions it as an integrative component in carbohydrate transport plasticity (Chincinska et al., 2013; Gong et al., 2015; Xu et al., 2017; Xue et al., 2016). In Populus species, SUT4 is expressed at much higher levels in overwintering than in summer stems and buds (Ko et al., 2011; Sreedasyam et al., 2023; Tuma et al., 2024), suggesting a potential seasonal role. Based on evidence of its importance for hydrostatic control and abiotic stress tolerance in various organs during the active growth of Populus tremula × alba INRA 717‐1B4 (hereafter, 717) (Frost et al., 2012; Harding et al., 2020; Harding et al., 2022; Xue et al., 2016), SUT4 may also serve nutritive and/or protective roles in overwintering trees.
A coppicing study using SUT4‐knockout (KO) 717 mutants provided evidence for a role of SUT4 in the emergence and early growth of epicormic buds under suboptimal winter glasshouse conditions (Tuma et al., 2024). Although starch and sucrose depletion magnitudes were similar between sut4 and wild‐type (WT) plants, epicormic bud release and sprout growth were reduced in the mutants, consistent with impaired vacuolar sucrose efflux (Tuma et al., 2024). Notably, this coincided with higher raffinose abundance in the winter stem of sut4 and a steeper decline following coppicing relative to WT. Raffinose, a trisaccharide synthesized directly from sucrose in the cytosol (Schneider & Keller, 2009), is known to contribute to membrane stabilization, reactive oxygen species (ROS) scavenging, and protection against winter freezing (Ameglio et al., 2004; Regier et al., 2010). Raffinose accumulation is modulated by multiple abiotic stress signaling pathways (Khan et al., 2021; Noronha et al., 2022; Pluskota et al., 2015), which may explain why its abundance does not consistently align with SUT4 expression in SUT‐KO or knockdown (KD) poplars (Frost et al., 2012; Harding et al., 2022; Tuma et al., 2024). Furthermore, storage water capacitance and modulus of elasticity differ between WT and SUT4‐KD lines (Harding et al., 2020). During drought stress, trajectories of leaf relative water content reductions also differ between WT and SUT4‐KO or KD lines (Harding et al., 2022). These findings suggest that the metabolic outcomes of SUT4 plasticity are shaped by multiple stress signaling pathways—some that directly modulate SUT4 expression (Gong et al., 2015), and others that are themselves modulated by SUT4 activity (Xue et al., 2016).
Under non‐stress conditions, plant growth, cell wall composition, and vessel development were altered in transgenic poplar with elevated raffinose levels via overexpression of a galactinol synthase gene (Unda et al., 2017). Although not necessarily raffinose‐dependent, spring bud break and growth resumption are partially influenced by stem vessel architecture, the timing of new xylem formation, and post‐winter hydraulic capacity (Lechowicz, 1984; Perrin et al., 2017). More broadly, raffinose may modulate salicylic acid signaling (La Mantia et al., 2017). In Populus, salicylic acid has been shown to affect carbon allocation toward flavonoid‐derived proanthocyanidins (i.e., condensed tannins), which have documented protective functions in leaves (Gourlay & Constabel, 2019; Ullah et al., 2019). Proanthocyanidins also play protective roles during reproductive development (Debeaujon et al., 2000; Dixon & Sarnala, 2020).
In poplar, vascular‐specific Type I SUT transcripts (SUT1 and SUT3 genome duplicates) are detected only in the summer, whereas the broadly expressed Type II (SUT5 and SUT6 genome duplicates) and Type III (single‐copy SUT4) transcripts are present year‐round, with SUT4 predominating during the cool seasons (Ko et al., 2011; Sreedasyam et al., 2023; Tuma et al., 2024). The present study investigated the effects of SUT4‐KO under field conditions. For comparison, we also generated single‐ and double‐KO mutants of the other winter‐expressed SUT5/SUT6 paralogs encoding plasma membrane transporters (Peng et al., 2014). SUT5/SUT6‐KO (hereafter sut5/6) mutants showed no observable effects on seasonal growth or carbohydrate metabolism relative to controls. In contrast, SUT4‐KO (sut4) mutants exhibited accelerated leaf senescence in the fall and delayed bud flush in the spring, culminating in reduced biomass growth. The field study further revealed SUT4‐dependent effects on the metabolic trajectories of developing female catkins. Together, these findings strengthen the rationale of SUT4 as a key modulator of sugar allocation and stress response mechanisms during seasonal growth transitions, with implications for perennial plant adaptation to changing climates.
RESULTS
Altered phenology and reduced stem growth in sut4 but not sut5/6 mutants
We established a field trial in summer 2021 to investigate the effects of winter‐expressed SUT gene knock‐outs on seasonal growth and phenology. Over 200 trees were planted in 20‐gal pots; Cohort 1 (control and sut4) in early June and Cohort 2 (control, sut4, sut5, sut6, and sut5/6) in mid‐August. In late October, following their first (partial) growth season, sut4 mutants showed signs of leaf yellowing while leaves of the other genotypes remained green regardless of Cohort (Figure 1a). By mid‐November, nearly 90% of sut4 trees were in a denuded state, whereas the majority of the controls and sut5, sut6, and sut5/6 mutants still retained most of their foliage (Figure 1b,c). Post‐dormancy bud flush the following spring (March 2022) was delayed by an average of 7 days in sut4 trees compared with the control, sut5, sut6, and sut5/6 plants (Figure 1d). Similar trends were observed in two subsequent years, suggesting a reduced leaf lifespan in sut4 trees.
*Biomass yield and growth characteristics of sut and control poplars.(a) Representative adaxial views of leaf plastochron index (LPI)‐4, LPI‐6, LPI‐8, LPI‐10, and LPI‐12 prior to leaf abscission.(b) Representative photos of field‐grown trees taken in November 2021.(c) Progression of leaf abscission from the main stem in Fall 2021. Data represent the percentage of trees per genotype that had abscised or desiccated leaves using pooled transgenic events and biological replicates (n = 69 for control, n = 78 for sut4, n = 13 for sut5, n = 11 for sut6, and n = 19 for sut5/6).(d) Bud flush was evaluated in the spring of 2022 based on the epicormic buds on the main stem. Data represent the percentage of trees per genotype that had flushed buds using pooled transgenic events and biological replicates (n = 69 for control, n = 78 for sut4, n = 13 for sut5, n = 11 for sut6, and n = 19 for sut5/6).(e) Stem diameter of trees from Cohort 1 and Cohort 2 measured 1 m above root collar monthly or bimonthly. Data are mean ± standard deviation of pooled transgenic events and biological replicates (n = 40 for control or 29 for sut4 in Cohort 1, or n = 23 for control, 40 for sut4, 13 for sut5, 11 for sut6, and 18 for sut5/6 in Cohort 2). The effects of time (t), genotype (g), and their interaction (t × g) were assessed with a repeated measures two‐way ANOVA. ***P ≤ 0.001. Inset in Cohort 1 was from size‐matched control or sut4 trees at the start of the monitoring (n = 13). Inset in Cohort 2 represent diameter increments during spring and summer of 2022.(f) Cambial growth reactivation in the spring of 2022 (Cohort 1) and 2024 (Cohort 2). Data represent the percentage of trees per genotype that had the first measurable diameter increases, with months indicated by numbers (n = 29–40 trees for Cohort 1 or n = 13–24 trees for Cohort 2).(g) Height of dominant stem measured at the beginning of the growing season in 2022 and 2023. Data are mean ± standard deviation of n = 15–22 trees per genotype using pooled transgenic events and biological replicates. Significance was conducted using two‐sample t‐test. P < 0.05.(h) Representative image of control, sut4, and sut5/6 trees 14 months after field outplanting.
Main stem diameter growth was monitored monthly for Cohort 1 and bimonthly for Cohort 2. Since no growth differences were observed among independent events across all traits, data were pooled by mutant for ease of comparison. Diameter growth in sut5, sut6, and sut5/6 trees was indistinguishable from controls, whereas sut4 trees exhibited significantly reduced growth during 2022 (Figure 1e). This reduction was more pronounced in Cohort 1 than in Cohort 2, consistent with previous reports of poor growth performance following summer outplanting (Grossnickle, 2005; Landhäusser et al., 2012). In Cohort 1, all control trees had resumed cambial growth and showed increased diameters by April 2022, whereas only 51% of sut4 trees exhibited measurable growth (Figure 1f).
To determine whether the delayed spring growth in sut4 trees was due to their smaller initial size, we analyzed 13 size‐matched control and sut4 trees. We found that sut4 diameter growth still lagged by 2–3 months compared with size‐matched controls (Figure 1e Cohort 1 inset). Similarly, in Cohort 2, diameter growth of sut4 trees between February and June 2022 was significantly reduced, reaching only half the growth observed in control and sut5/6 trees (Figure 1e Cohort 2 inset). However, growth rates were comparable among genotypes from June to October 2022 (Figure 1e Cohort 2 inset). The 2022–2023 winter was mild (Figure S1), and diameter differences between sut4 and the other genotypes remained constant through the end of the field trial in April 2023 (Figure 1e). A subset of Cohort 2 trees was permitted to continue, and by April 2024, diameter growth had resumed in 88% of control and 92% of sut5/6 trees, but in only 58% of sut4 trees (Figure 1f).
Growth was unaltered in both the single‐ and double‐KO of SUT5/SUT6; therefore, only sut5/6 trees were included in subsequent analyses. Tree height was slightly reduced, by approximately 10%, in sut4 compared with the controls and sut5/6 trees in April of 2022 (Figure 1g,h). By April 2023, sut4 trees were ~20% shorter than control and sut5/6 trees (Figure 1g). Taken together, reduced growth, shorter leaf lifespans, and delayed spring cambial growth were consistently observed in sut4 trees over multiple years.
Altered non‐structural carbohydrate abundance in sut4 stems
We compared levels of monosaccharides and oligosaccharides in the bark and xylem tissues sampled during summer and winter seasons. Bark and xylem sucrose levels were higher in sut4 mutants than in the other lines during the summer (Figure 2a), consistent with previous findings from greenhouse‐grown plants (Harding et al., 2022; Payyavula et al., 2011; Tuma et al., 2024). Sucrose levels decreased during the winter most drastically in sut4, neutralizing most but not all of the summer genotypic increases (Figure 2a). Glucose and fructose, the major hexoses, exhibited low abundances in all lines during the summer and increased in the winter by 1–2 orders of magnitude in both tissues, reaching highest abundances in sut4 in both bark and xylem (Figure 2b,c). Bark and xylem starch abundance differed little between genotypes and exhibited similarly large declines in all genotypes during winter (Figure 2d).
*Non‐structural carbohydrate abundance in stem organs of different genotypes.(a) Sucrose, (b) glucose, (c) fructose, and (d) starch abundance in bark and xylem during summer and winter. Data are mean ± standard deviation of n = 3–6 biological replicates. Significance was determined by Student's t‐test against WT. *P ≤ 0.05; **P ≤ 0.01; **P ≤ 0.001.
Raffinose and its precursor galactinol are known to accumulate under desiccation stress such as drought and cold (Yan et al., 2022). Galactinol and raffinose were only marginally detected in summer whereas their precursor myo‐inositol was readily detected and significantly more abundant in both bark and xylem of sut4 trees compared with other genotypes (Figure 3). During winter, galactinol and raffinose levels increased sharply in all genotypes, presumably at the expense of myo‐inositol (Figure 3). However, these winter increases were significantly attenuated in sut4 bark and trended so in xylem (Figure 3b,c). Overall, the data raise the possibility that altered subcellular partitioning of sugars, rather than total carbohydrate availability per se, could contribute to the developmental delays in spring bud flush and stem diameter growth in sut4 trees. Furthermore, altered raffinose dynamics under outdoor winter conditions may also be relevant to these growth phenotypes.
*Oligosaccharide abundance dynamics in stem organs of different genotypes.(a) Myo‐inositol, (b) galactinol, and (c) raffinose abundance in bark and xylem during summer and winter. Data are mean ± standard deviation of n = 3–6 biological replicates. Significance against WT was determined by Student's t‐test. *P ≤ 0.05; **P ≤ 0.01; **P ≤ 0.001.
Wood physicochemical properties were altered in sut4 trees
In light of the reduced stem diameter growth observed in sut4 lines, wood samples were analyzed for changes in chemical and physical properties. Neither lignin content nor syringyl‐to‐guaiacyl (S/G) monolignol ratio was altered in sut4 or sut5/6 trees compared with controls (Figure 4a–c). Acoustic velocity also did not differ between genotypes (Figure 4d). However, specific gravity and the modulus of elasticity were reduced in sut4, but not sut5/6, relative to the controls (Figure 4e,f). These findings suggest that vacuolar sequestration of sucrose may exert complex effects on sut4 trees, influencing not only diameter growth but also structural development and wood properties.
*Wood characteristics and chemical properties.(a) Total lignin content and (b) syringyl‐to‐guaiacyl lignin (S/G) ratio determined by pyMBMS, (c) total Klason lignin content, (d) acoustic velocity, (e) specific gravity, and (f) modulus of elasticity. Data are mean ± standard deviation of n = 2–4 biological replicates. Significance was determined against WT trees by Student's t‐test; **P ≤ 0.01; **P ≤ 0.001.
Reproductive growth and development were altered in sut4 trees
Field‐grown female 717 trees typically flower after reaching 5–8 years of age (Mohamed et al., 2010). However, during the spring of year 2 and prior to leaf emergence, catkins were unexpectedly observed on many nursery trees (Figure 5a). Catkin emergence frequency did not differ between genotypes in either spring 2023 or 2024. In 2023, flowering frequency appeared to correlate with tree age: an average of 69% of Cohort 1 trees produced catkins compared with 36% in Cohort 2 which had been released 2 months later (Figure 5b). In the extended trial with a subset of Cohort 2 trees, 75% produced catkins in 2024, regardless of genotype (Figure 5b).
*Flowering phenotypes in control and sut trees.(a) Axillary inflorescence bud flushing in early spring 2023.(b) Percentage of trees per genotype from Cohort 1 and Cohort 2 that developed catkins during spring of 2023 and 2024 (n = 41 control or 29 sut4 trees for 2023 Cohort 1; n = 27 control, 49 sut4, or 18 sut5/6 trees for 2023 Cohort 2; n = 24 control, 36 sut4, or 13 sut5/6 trees for 2024).(c) Representative images of seeds and ovules from WT and sut4 catkins. Scale bar = 1 mm.(d) Representative catkin developmental stages used for metabolite profiling (I, unopened floral buds; II, newly emerged catkins; and III, elongating catkins). Bar = 1 cm.(e) Abundance of sucrose, glucose, fructose, galactinol, raffinose, citric acid, malic acid, and succinic acid during catkin development. Data are mean ± standard error of n = 6–8 biological replicates. Significance was determined by Student's t‐test. *P ≤ 0.05; **P ≤ 0.001.
To test whether SUT4‐KO affected fertility, we performed controlled pollination using wild P. tremuloides pollen. Of the 13 WT and 14 sut4 catkins pollinated, nine from each genotype developed capsules with abundant seed trichomes. While WT flowers produced seeds as expected, no seeds were recovered from sut4 capsules (Figure 5c). Although unfertilized ovules in aspens can undergo limited development (Fechner, 1972), the ovules in sut4 capsules were notably smaller than those in WT (Figure 5c). It is unclear whether the observed sterility in sut4 trees was due to ovule failure, other developmental anomalies, or environmental sensitivity associated with SUT4 disruption.
Metabolic profiling revealed contrasting developmental trajectories between WT and sut4 catkins
SUT4 is the predominant SUT gene expressed in poplar flowers (JGI Plant Gene Atlas, Sreedasyam et al., 2023), suggesting that cytosolic access to vacuolar sucrose reserves is likely important during floral development. Given that the timing of ovule development relative to catkin growth is known (Wang et al., 2012), we reasoned that metabolic profiling of developing catkins could reveal how loss of SUT4 function impacted fertility. Metabolic profiling was conducted on unopened floral buds, newly emerged catkins, and elongating catkins, designated as stages I, II, and III, respectively (Figure 5d). The temporal dynamics of sucrose abundance differed markedly between genotypes. In sut4 catkins, sucrose levels increased steadily during development, whereas in WT, they remained stable (Figure 5e).
To assess whether the changes in sut4 reflected increased vacuolar sequestration, we next examined sucrose‐derived metabolites that originate in the cytosol. Glucose and fructose levels increased in both genotypes during catkin expansion (Figure 5e). Yet, this is not a reliable indicator of cytosolic sucrose, as hexoses can be derived from starch or sucrose pools in the vacuole, plastid, or cytosol. In contrast, the biosynthesis of raffinose occurs exclusively in the cytosol and directly utilizes sucrose (Schneider & Keller, 2009). In WT, both galactinol and raffinose peaked in unopened floral buds and declined sharply during catkin development despite stable sucrose levels (Figure 5e). The pattern is consistent with high raffinose abundance in dormant poplar buds (Watanabe et al., 2018). However, this trend was absent in sut4 trees. Raffinose levels were significantly lower in stage I sut4 buds compared with WT, even though sucrose levels were similar. As catkins elongated and sucrose accumulated in sut4, raffinose, and to a lesser extent galactinol, also increased (Figure 5e).
We also compared several Krebs cycle intermediates across all three stages of catkin development. Malic acid, the most abundant intermediate, increased throughout catkin development and did not differ between genotypes (Figure 5e). In contrast, citric acid levels in sut4 buds were less than half those of WT at stage I, partially recovered at stage II, and became comparable by stage III (Figure 5e). This shift is notable given citric acid's central role in fatty acid and acyl metabolism, and its connection to gluconeogenesis via the glyoxylate shunt under carbohydrate‐limited conditions (Eastmond & Graham, 2001; Smith, 2002). Succinic acid, a product of citric acid metabolism during glyoxylate shunt activity, was significantly elevated in sut4 catkins throughout development (Figure 5e). These findings suggest that SUT4‐KO affected metabolic pathways beyond carbohydrate metabolism.
Transcriptional profiling revealed elevated stress responses in sut4 catkins
To investigate whether sut4 infertility was due to insufficient protection during ovule development or to an inherent constitutive stress response, we performed RNA‐seq analysis on stage III elongating catkins. We identified 821 significantly upregulated and 660 downregulated genes in sut4 catkins compared with controls (Dataset S2). Gene ontology (GO) categories, such as protein oligomerization, protein refolding, responses to heat, hypoxia, and hydrogen peroxide were significantly over‐represented among the upregulated genes (Figure 6a). Interestingly, none of these GO categories were enriched among differentially expressed genes in vegetative tissues (e.g., leaves, xylem, bark, root tips) of SUT4‐KD plants under benign greenhouse conditions (Xue et al., 2016). Specifically, over 22% of the approximately 360 genes annotated to encode various heat shock proteins (HSPs) (Yer et al., 2018) were significantly upregulated in sut4 catkins. Also upregulated were genes encoding several heat shock factors (HSFs) and late embryogenesis abundant (LEA) proteins. All three galactinol synthase genes detected in elongating catkins were sharply induced in sut4 catkins, including the Populus alba × grandidentata ortholog previously shown to exhibit cool season‐biased expression (Unda et al., 2012), but the expression of raffinose synthase genes remained largely unchanged.
*Gene expression responses in Stage III elongating catkins.(a) GO enrichment of differentially upregulated or downregulated genes in the elongating catkins of the sut4 mutants relative to control. Representative GO terms with fold‐enrichment >2 and gene number >5 are shown.(b) Heatmaps showing expression responses of genes associated with flavonoid and proanthocyanidin biosynthesis and regulation and circadian rhythm. Log2FC, log2‐transformed fold change (sut4/control) of n = 7 (pooled from 4 WT and 3 Cas9) and 5 (sut4) biological replicates. ANR, anthocyanidin reductase; ANS, anthocyanidin synthase; CHS, chalcone synthases; CHI, chalcone isomerase; DFR, dihydroflavonol reductase; EFL, early flowering; F3H, flavanone 3‐hydroxylase; F3′H, flavonoid 3′‐hydroxylase; F3′5′H, flavonoid 3′,5′‐hydroxylase; GI, gigantea; GRP, glycine‐rich RNA‐binding protein; LAR, leucoanthocyanidin reductase; LHY, late‐elongated hypocotyl; LWD, light‐regulated WD repeat‐containing protein; TT, transparent testa; TTG, transparent testa glabra; PRR, pseudo‐response regulator.(c) Catechin abundance during catkin development (top) or in pollinated carpellate flowers, with a representative image shown on the right (bottom). Data are mean ± standard error of n = 5–6 biological replicates. Significance was determined by Student's t‐test. *P ≤ 0.01; **P ≤ 0.01; **P ≤ 0.001.
The smaller group of downregulated genes was enriched for GO terms associated with flavonoid—particularly proanthocyanidin—biosynthesis, lipid biosynthesis, responses to UV‐B and sucrose stimulus, mitotic cell cycle phase transition, seed coat development, and circadian rhythm (Figure 6a). Nearly, all flavonoid pathway genes (Tsai et al., 2006) and several upstream regulators detected in catkins were significantly downregulated in sut4 catkins (Figure 6b). Also downregulated were genes encoding orthologs of Arabidopsis TRANSPARENT TESTA12 (TT12), a multidrug and toxin extrusion (MATE)‐type proton antiporter, and TT13 (also known as AHA10), a P‐type H^+^‐ATPase proton pump that energizes TT12‐mediated proanthocyanidin transport into the vacuole (Debeaujon et al., 2001; Marinova et al., 2007). In Arabidopsis, both TT12 and TT13 are specifically expressed in ovules and developing seeds (Appelhagen et al., 2015; Debeaujon et al., 2001). tt12 and tt13 mutants show defects in vacuole development and proanthocyanidin deposition in seed coat endothelial cells, resulting in reduced seed coat pigmentation and decreased dormancy (Appelhagen et al., 2015, Debeaujon et al., 2001). To examine whether sut4 catkins exhibit reductions in proanthocyanidin abundance, we compared levels of the proanthocyanidin precursor catechin between genotypes. Consistent with the RNA‐seq data, catechin levels were significantly reduced during catkin development, as well as in individual carpellate flowers after pollination in sut4 compared with controls (Figure 6c).
Several genes encoding core components of the circadian clock system were significantly downregulated in sut4 catkins. These include LHY1, LHY2 (late‐elongated hypocotyl), GI (gigantea), GRP7a, GRP7b (glycine‐rich RNA‐binding protein 7, also called CCR2 for cold circadian rhythm RNA‐binding 2), PRR9a, and PRR9b (pseudo‐response regulator 9) (Edwards et al., 2018). On the other hand, PRR7a and PRR7b, which act antagonistically with LHY (Nakamichi et al., 2007), were upregulated (Figure 6b). Many circadian clock genes are sensitive not only to light and temperature cues, but also to sucrose and abiotic stress signaling, influencing developmental, physiological, and metabolic functions (Haydon et al., 2017; Liang et al., 2024; Oravec & Greenham, 2022). Collectively, the transcriptomic data from catkins implicated elevated basal stress and circadian dysregulation as consequences of altered sucrose compartmentalization due to SUT4‐KO.
DISCUSSION
Knockout of
SUT4 but not of SUT5 / SUT6 accelerated autumn leaf senescence and delayed spring bud break and stem growth resumption
Seasonal alternation between carbohydrate storage during photoautotrophic growth and remobilization during heterotrophic subsistence is a hallmark of the perennial lifestyle (Regier et al., 2010; Tixier et al., 2019). Using CRISPR‐induced frameshift mutants, we investigated the role of winter‐expressed SUT plasticity during seasonal transitions. We found that CRISPR‐KO of SUT4, which encodes a tonoplast‐localized transporter for intracellular trafficking, but not SUT5/SUT6, which encode plasma membrane‐localized transporters for export from the cell (Peng et al., 2014), had detrimental effects on leaf phenology and stem diameter growth in the field. The absence of phenotypes in sut5/6 trees may reflect compensatory activity by SWEET15b, a plasma membrane‐localized sucrose transporter with strong winter expression in poplar xylem (Ko et al., 2011; Zhang et al., 2021). In contrast, compensation for the loss of SUT4 appears less likely, as none of the SWEETs known to be expressed in poplar xylem encode tonoplast‐localized sucrose transporters (Eom et al., 2015; Zhang et al., 2021).
sut4 trees consistently exhibited premature leaf senescence, delayed spring budbreak, and delayed diameter growth relative to control and sut5/6 trees (Figure 1), effectively shortening their growing season. Leaf senescence typically spans 3–5 weeks (Figure 1c), beginning with a sharp decline in chlorophyll and photosynthesis, followed by rapid cellular breakdown and nutrient resorption (Keskitalo et al., 2005). While SUT5/SUT6 is weakly expressed in source leaves, SUT4 is well expressed (Payyavula et al., 2011), which may explain the lack of senescence phenotypes in sut5/6 trees. A premature decline in photosynthetic capacity in sut4 trees would be expected to lead to intracellular sucrose and turgor imbalances, further exacerbating the already elevated vacuolar sucrose sequestration. Although SUT4‐KD or KO has been shown to alter leaf hydrostatics and pressure–volume characteristics (Harding et al., 2020), it does not affect leaf morphology or function under favorable growth conditions (Frost et al., 2012). Whether these physiological changes influence the timing or progression of autumn senescence remains unclear.
Low raffinose may reflect metabolic or osmotic perturbations due to loss of SUT4 plasticity during winter
The early onset of senescence and premature leaf drop in sut4 trees may have impaired the buildup of bud reserves and contributed to delayed spring bud break. Although vegetative buds were not analyzed, flowering bud data suggest that total soluble sugar levels were not limiting in spring (Figure 5e). Similarly, while spring diameter growth was delayed in sut4 trees, summer growth rates were comparable to controls across multiple years (Figure 1e, Ccohort 2). Notably, winter stem sugar and starch reserves were similar or even higher in sut4 mutants (Figure 2). Previous work has shown comparable sucrose and starch consumption rates in sut4 and WT plants under carbon‐limited conditions imposed by coppicing (Tuma et al., 2024). These findings suggest that spring growth delays may stem from factors other than reserve size or depletion rate.
Winter stem raffinose levels were much lower in sut4 trees (Figure 3). Given the antioxidant and osmoprotective roles of raffinose (Kornberg & Krebs, 1957; Regier et al., 2010), its attenuated accumulation may have contributed to delayed spring bud break and growth resumption. Among the possible causes are limited cytosolic sucrose (Schneider & Keller, 2009) and altered stress signaling upstream of raffinose biosynthesis (Khan et al., 2021; Noronha et al., 2022; Pluskota et al., 2015). Vacuolar sucrose sequestration in sut4 trees could constrain raffinose synthesis as seen in drought‐stressed SUT4‐KD leaves (Frost et al., 2012). SUT4 has been implicated in stress modulation, with SUT4‐KD and KO plants exhibiting changes in leaf pressure–volume curves, relative water content, and storage water capacitance during drought (Harding et al., 2020; Harding et al., 2022). A signaling role has also been suggested for galactinol, a precursor of raffinose, where its hyperaccumulation led to stunted growth and altered xylem properties in poplar (Unda et al., 2017). Together, these findings support a role for SUT4 in modulating sugar allocation and stress signaling, with consequences for seasonal growth.
Perturbed sucrose and raffinose trajectories were associated with elevated stress during sut4 catkin development
Galactinol and raffinose are known to accumulate in dormant buds and developing seeds to confer desiccation and cold tolerance (Cox & Stushnoff, 2001; Falavigna et al., 2018; Peterbauer & Richter, 2001). Meiosis during gamete development is particularly stress‐sensitive (Thakur et al., 2010) and raffinose has been shown to enhance floral bud hardiness (Cox & Stushnoff, 2001; Falavigna et al., 2018). During sut4 catkin development, raffinose levels were low at stage I—when female meiosis is expected to have begun (Wang et al., 2012)—while WT buds peaked at this stage with multifold higher raffinose accumulation (Figure 5e). Since raffinose synthesis is modulated by cold and desiccation stress signaling (Peterbauer & Richter, 2001; Yan et al., 2022), the steady decline of galactinol and raffinose levels after stage I in WT is consistent with reduced stress. In contrast, sut4 catkins showed a sustained increase, possibly reflecting delayed or abnormal metabolic activity under cytosolic sucrose limitation. Supporting this idea, sut4 catkins had elevated succinic acid and reduced citric acid abundance during emergence and early elongation (Figure 5e). One interpretation of this metabolic shift is increased gluconeogenesis to fuel dormancy release and reproductive development under sucrose‐limited conditions (Eastmond et al., 2015; Fadón et al., 2020; Walker et al., 2021; Watanabe et al., 2018). Sustained raffinose levels may also indicate persistent stress in sut4 catkins due to altered sucrose partitioning or stress signaling as discussed above. GO term enrichment for responses to heat, hypoxia, and hydrogen peroxide, along with protein oligomerization and refolding among genes upregulated in sut4 catkins is consistent with heightened cellular stress (Figure 6a).
High carbohydrate levels usually promote flavonoid biosynthesis, especially under low‐temperature conditions (Araguirang & Richter, 2022). The widespread suppression of flavonoid gene expression in sut4 catkins is therefore counterintuitive given the elevated sucrose levels, but may be consistent with cytosolic sucrose limitation. This may also suggest broader disruptions in cellular homeostasis. Impaired SUT4 symport activity for vacuolar sucrose efflux may have altered tonoplast membrane potential and affected other proton‐dependent tonoplast transporters (Martinoia, 2017). Supporting this idea, the single‐copy AHA10/TT13 gene required for vacuolar proanthocyanidin transport (Appelhagen et al., 2015) was downregulated in sut4 catkins. The coordinated downregulation of TT12, TT13, and their upstream regulator TTG2, which encodes a WRKY transcription factor (Gonzalez et al., 2016), points to cascading effects of SUT4‐KO on catechin accumulation, vacuolar biogenesis and seed development. Concurrent upregulation of HSPs associated with protein oligomerization and refolding responses further supports a signaling role for SUT4 during catkin development.
Loss of SUT4 function disrupted circadian clock gene expression in catkins
Flavonoid biosynthesis is positively gated by the circadian morning complex anchored by LHY (Harmer et al., 2000), and the circadian clock itself is modulated by sucrose, flavonoids, and various hormonal signals (Dalchau et al., 2011; Haydon et al., 2013; Haydon et al., 2017; Hildreth et al., 2022). In sut4 catkins, morning‐ or daytime‐expressed clock genes (PtaLHYs, PtaGI, PtaPRR9s) were downregulated, while evening‐ or nighttime‐expressed genes (PtaEFL3 and PtaPRR7s) were upregulated (Figure 6b) (Ding et al., 2018; Takata et al., 2009; Toda et al., 2019). This pattern mirrors sucrose‐modulated inverse regulation of AtLHY and AtPRR7 in Arabidopsis (Haydon et al., 2013), though AtPRR9 was unresponsive unlike PtaPRR9s. Differential responses among orthologous PRRs are not surprising, given species‐specific expression divergence that may reflect adaptive tuning to distinct—particularly perennial—environmental conditions (Toda et al., 2019). It is worth noting that clock gene expression was unchanged in SUT4‐KD plants under well‐watered summer greenhouse conditions (Xue et al., 2016). This suggests that SUT4 plays a specific role in cool‐season sugar sensing, especially in reproductive tissues where it is highly expressed. Consistent with this idea, transgenic poplar with misregulated clock genes LHY1, LHY2, and GI exhibited altered phenology, including premature leaf senescence, delayed bud break, reduced growth, and compromised freeze tolerance (Ding et al., 2018; Fataftah et al., 2021; Ibáñez et al., 2010), similar to those observed in sut4 trees.
SUT4 integrates sugar allocation, stress response, and circadian regulation
SUT4 expression is responsive to abiotic stresses, including drought and heat, across multiple species (Xu et al., 2017), and its induction under cool‐season conditions has been recently reported in poplar (Tuma et al., 2024). Mutation or downregulation of SUT4 has been shown to affect biomass partitioning, flowering, and seed yield in rice (Oryza sativa), potato (Solanum tuberosum), and tomato (Solanum lycopersicum), with circadian gene expression changes also reported in potato (Chincinska et al., 2008; Chincinska et al., 2013; Eom et al., 2011; Liang et al., 2023). The present work provides evidence supporting a role for SUT4 in regulating cytosolic sucrose availability during carbohydrate reserve depletion in winter stem and spring catkins of poplar. Loss of this capacity appears to have repercussions for both vegetative and reproductive growth, protective metabolism, and circadian clock function. Taken together, our results highlight the potential of SUT4 to coordinate sugar signaling, stress responses, circadian regulation, and seasonal development in poplar.
MATERIALS AND METHODS
Production of
SUT ‐KO lines
The generation of sut4 mutants and Cas9‐only vector controls in the Populus tremula × alba INRA 717‐1B4 background was previously reported (Harding et al., 2022; Zhou et al., 2015). Two gRNAs were designed to target PtaSUT5 and PtaSUT6 individually to generate single mutants, and a third gRNA was designed to target the highly similar genome duplicates at a consensus sequence to generate double mutants. Target sites were cross‐checked against the Populus VariantDB (Zhou et al., 2025) to ensure the absence of sequence polymorphisms. Oligo pairs (Table S1) were assembled into the p201N‐Cas9 vector behind the Medicago truncatula U6.6 promoter, following the approach of Bewg et al. (2022). *Agrobacterium‐*mediated transformation and plant regeneration were performed as described (Bewg et al., 2022).
Amplicon sequencing to determine mutation patterns
Leaves from independent events of tissue‐cultured plants were collected for genomic DNA extraction and amplicon sequencing library preparation (Bewg et al. 2022) using consensus primers (Table S1). Samples were then barcoded with Illumina amplicon indexing primers, pooled, and sequenced on an Illumina MiSeq at the Georgia Genomics and Bioinformatics Core at the University of Georgia to obtain PE150 reads. Sequencing reads were processed and indel patterns identified using the Analysis of Genome Editing by Sequencing (AGEseq) program (Xue & Tsai, 2015) with a mismatch allowance set at 1%, followed by manual curation. Transgenic events with biallelic (single KO) or tetra‐allelic (double KO) edits were selected for further experiments (Dataset S1).
Plant propagation and field establishment
Multiple independent lines for sut4 (n = 8), sut5 (n = 3), sut6 (n = 2), and sut5/6 (n = 3) mutants along with WT and Cas9 controls were propagated from single‐internode stem cuttings (Frost et al., 2012). Cuttings were transferred to 1‐gallon pots containing a commercial soil mixture (Fafard 3B) supplemented with Osmocote (15‐9‐12 NPK, 3‐month slow release) and placed in a walk‐in growth chamber for approximately 3 weeks (Tuma et al., 2024). Plants were moved into an adjacent greenhouse for 2 weeks before transfer to a lath house within a fenced nursery at the University of Georgia's Whitehall Forest for an additional 2 weeks. For logistical reasons having to do with the timing of cutting establishment, sut4 plants were slightly shorter than control plants at the time of outplanting. Plants were transferred to 20‐gallon pots containing a mix of Jolly Gardener premium soil (75% aged pine bark, 25% peat moss) and Fafard 3B with additional Osmocote (15‐9‐12 NPK, 3‐month slow release) and planted at the nursery in summer 2021. Genotypes were randomized throughout the site and spaced 1.5 meters apart from one another. The first planting occurred in early June 2021 for sut4 mutants and controls (WT and Cas9), and the second planting occurred in mid‐August 2021 for sut4, sut5, sut6, sut5/6 and controls. Plants were watered using an overhead irrigation system for 2–4 h daily from early April to late October. The field trial was conducted with approval from the Animal and Plant Health Inspection Service (APHIS) of the U.S. Department of Agriculture.
Field growth and phenology monitoring
The diameter of the main stem approximately 1 m above the soil surface was measured monthly or bimonthly from August 2021 to April 2023, and after termination of most trees, monthly through August 2024 for the remaining trees. Plant height was measured from the base of the stem at soil level to the shoot tip in April 2022 and 2023. Trees were monitored throughout the fall to compare the onset and rate of leaf abscission between genotypes. Leaf color was compared between genotypes as an indicator of leaf senescence in October. In the spring, the appearance of bud flush on the dominant stem and flowering were recorded. Floral buds and catkin emergence were observed during the spring of 2023 (late February) and 2024 (early March). The total number of trees that developed catkins was recorded prior to their removal from the field site per the APHIS regulation. Further information regarding the field trial monitoring, phenotyping, and sampling is shown in Figure S1.
Tissue sampling
Approximately 12–17 months after outplanting, increment cores (10 mm in length and 2 mm in diameter) were collected from the main stem approximately 1.3 meter above soil level using a micro‐coring device (Costruzioni Meccaniche Carabin C, San Vito di Cadore, Italy) according to Rossi et al. (2006). Stems were approximately 2–6 cm in diameter at the time of sampling. Increment cores were collected from the same stems during active growth in August 2022 (summer) and during dormancy in January 2023 (winter). Xylem and bark were separated from the cores in the field, snap‐frozen in liquid nitrogen before being transported back to laboratory facilities where they were stored at −80°C until sample processing occurred. Temperatures ranged from a night‐time low of 22°C to a daytime high of 31°C (daily average 25°C) on the day of summer sampling, and from a low of 3°C to a high of 18°C (daily average 11°C) on the day of winter sampling (Figure S1). Catkins were observed in late February 2023 after the second winter in the field. Three developmental stages of catkins from Cohort 1 trees were collected on March 1, 2023 for transcript and metabolite profiling. All tissue sampling occurred under sunny conditions between 11 AM and 2 PM. Tissues were snap‐frozen, ground under liquid nitrogen with a mortar and pestle, and stored at −80°C.
Controlled pollination
Branches containing flower buds were collected from WT and sut4 trees and incubated in 1/4 MS macro salt solution at room temperature in a confined laboratory to force flower. After catkin flowers opened, wild P. tremuloides pollen sourced from Minnesota in 2022 (with assistance of Kathy Haiby, Poplar Innovations Inc.) was brushed onto individual WT, Cas9, and sut4 catkins, which were then covered with glassine bags. After 7–10 days, the capsules opened, and the cottony fuss was collected for seed observation under a dissecting microscope. The length of catkins, seed size and number, and ovule size were recorded. The absence of seeds from pollinated sut4 flowers was first observed in 2023, and the controlled pollination experiments were repeated in 2024 and 2025 with reproducible results. Individual carpellate flowers collected 3–4 days after pollination in 2025 were flash‐frozen in liquid nitrogen for catechin analysis by GC–MS as described below.
Metabolite profiling
Tissues were lyophilized for 48 h (FreezeZone 2.5, Labconco), ground through a 40‐mesh sieve using a Wiley Mill (Thomas Scientific), and further ball‐milled to a fine powder in a S1600 Mini G Bead‐beater (Spex SamplePrep; Metuchen, New Jersey) for metabolite profiling by Gas Chromatography–Mass Spectrometry (GC–MS) as detailed previously (Harding et al., 2022; Tuma et al., 2024). Peak identities were assigned based on the NIST08, FiehnLib metabolomics library (Agilent) (Kind et al., 2009) and in‐house spectral libraries from authentic standards (Chen et al., 2014; Harding et al., 2022; Xue et al., 2013). A standard mix of succinic acid, sucrose, glutamic acid, fructose, glucose, and ascorbic acid was loaded at the beginning and end of each sample sequence to monitor derivatization and instrument performance. Peak intensities were normalized by the internal standard and reported on a sample dry weight basis (normalized peak area). Sucrose and hexose abundance (sum of fructose and glucose) were reported as μg/mg dry weight based on calibration curves.
Starch
Starch content was determined by hydrolyzing starch to glucose using enzymatic digestion with α‐amylase and amyloglucosidase (Chow & Landhausser, 2004) using 10 mg of lyophilized, Wiley‐ and ball‐milled tissue powder as previously described (Tuma et al., 2024). Starch abundance was reported as μg/mg dry weight based on a glucose calibration curve.
Wood chemical composition and physical property analyses
Fifteen‐cm long pieces of the main stem were destructively harvested following bud flush in March 2023. For wood chemistry analysis, stem segments were debarked, dried at 55°C for 24 h, and ground to pass through a 40‐mesh screen using a Wiley mill. Samples were then run through 99 cycles of hot ethanol extraction using a Soxhlet extraction unit (Buchi) and air dried. Total lignin content and the ratio of lignin monomers in the stems (syringyl/guaiacyl, S/G ratio) were determined by pyrolysis‐molecular beam mass spectrometry according to Harman‐Ware et al. (2022). Total lignin content was also determined by the Klason method as described (Swamy et al., 2015). Three approximately 7.5 mm longitudinal wood sections were obtained from the main stem approximately 1 m above soil level for analysis of specific gravity, acoustic velocity, and modulus of elasticity as described (Dahlen et al., 2023; Tsai et al., 2020; Vieilledent et al., 2018).
RNA‐seq analysis
Snap‐frozen catkin tissues from Cohort 1 trees in 2023 were ground to a fine powder under liquid nitrogen and an aliquot was used for RNA extraction using the Direct‐zol RNA Miniprep kit (Zymo) with PureLink Plant RNA Reagent (Life Technologies) as described (Harding et al., 2018). Illumina low input RNA (with PolyA selection) library preparation and NovaSeq sequencing (NovaSeq XP V1.5 reagent kits, 10B flowcell, 2 × 151 run length) were performed at the Joint Genome Institute (proposal: 10.46936/10.25585/6000884). RNA‐seq reads were aligned to the Populus tremula haplotype of the 717 reference genome (Zhou et al., 2023) using STAR‐2.7.10 (Dobin & Gingeras, 2015) with the following parameters: ‐‐alignMatesGapMax 20 000, ‐‐alignIntronMax 10 000, ‐‐outFilterScoreMinOverLread 0.1, and ‐‐outFilterMatchNminOverLread 0.1. Expression levels were estimated using featureCounts from the Subread package v2.0.6 with the options ‐Q 2, ‐s 0, ‐T 12, ‐p, and ‐‐countReadPairs ‐C (Liao et al., 2014) and reported as FPKM (fragments per kilobase of transcript per million mapped reads). Differential gene expression analysis were performed using DESeq2 v1.34.0 (Love et al., 2014). Thresholds for significant differential expression were based on minimum FPKM ≥5 in all samples of at least one group, Q ≤ 0.005, and fold change ≥1.5. Gene ontology (GO) enrichment analysis of significantly upregulated or downregulated genes was performed using ShinyGO v0.77 (Ge et al., 2020) available at https://aspendb.uga.edu/ShinyGO/. Expression responses of selected genes were visualized using the HeatMapperPlus tool (https://bar.utoronto.ca/ntools/cgi‐bin/ntools_heatmapper_plus.cgi).
Statistics
Repeated measures analysis of variance (ANOVA) was used to determine significant differences in growth over time, genotypic effects, and their interactions using R v4.1.0 (https://www.r‐project.org/) in Rstudio v.2022.12.353. All other reported genotypic contrasts were assessed with Student's t‐test (α = 0.05).
Author Contributions
TTT, SAH, and CJT conceived the study and designed all experiments. TTT performed all experiments and analyzed the data, with assistance from HAM, HP, and WPB for transgenic plant production and amplicon sequencing, SAH for metabolite data analyses, BL for RNA extraction, AL, KWB, and RZ for RNA‐seq analysis, DLWW for diameter and flower monitoring, MD and RZ for controlled pollination, JD for wood property analysis, and SS and AEHW for wood chemistry analysis. TTT prepared the manuscript which was revised by CJT with contributions from SAH.
Conflict of Interest
The authors have not declared a conflict of interest.
Supporting information
Dataset S1. Mutation patterns of SUT5, SUT6, and SUT5/6 edited transgenic lines. Mutation patterns are color‐coden for WT‐like (green, no edit), indels (orange) or no amplification (gray) likely due to large deletions. The two parental alleles are labeled as a (alba) or t (tremula).
Dataset S2. Differentially expressed genes in elongating catkins. Differential expression (sut4/control) criteria: Q ≤ 0.005, fold change (FC) ≥ 1.5, FPKM (fragments per kilobase of transcript per million mapped reads) ≥ 5.
Table S1. Primers. Figure S1. Field trial timeline and temperature profiles.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ameglio, T. , Decourteix, M. , Alves, G. , Valentin, V. , Sakr, S. , Julien, J.L. et al. (2004) Temperature effects on xylem sap osmolarity in walnut trees: evidence for a vitalistic model of winter embolism repair. Tree Physiology, 24, 785–793.15123450 10.1093/treephys/24.7.785 · doi ↗ · pubmed ↗
- 2Appelhagen, I. , Nordholt, N. , Seidel, T. , Spelt, K. , Koes, R. , Quattrochio, F. et al. (2015) TRANSPARENT TESTA 13 is a tonoplast P 3A‐AT Pase required for vacuolar deposition of proanthocyanidins in Arabidopsis thaliana seeds. Plant Journal, 82, 840–849.10.1111/tpj.1285425891958 · doi ↗ · pubmed ↗
- 3Araguirang, G.E. & Richter, A.S. (2022) Activation of anthocyanin biosynthesis in high light – what is the initial signal? New Phytologist, 236, 2037–2043.36110042 10.1111/nph.18488 · doi ↗ · pubmed ↗
- 4Bewg, W.P. , Harding, S.A. , Engle, N.L. , Vaidya, B.N. , Zhou, R. , Reeves, J. et al. (2022) Multiplex knockout of trichome‐regulating MYB duplicates in hybrid poplar using a single g RNA. Plant Physiology, 189, 516–526.35298644 10.1093/plphys/kiac 128PMC 9157173 · doi ↗ · pubmed ↗
- 5Calinger, K. & Curtis, P. (2023) A century of climate warming results in growing season extension: delayed autumn leaf phenology in north central North America. P Lo S One, 18, e 0282635.36867631 10.1371/journal.pone.0282635 PMC 9983848 · doi ↗ · pubmed ↗
- 6Chen, H.‐Y. , Babst, B.A. , Nyamdari, B. , Hu, H. , Sykes, R. , Davis, M.F. et al. (2014) Ectopic expression of a loblolly pine class II 4‐coumarate:Co A ligase alters soluble phenylpropanoid metabolism but not lignin biosynthesis in Populus. Plant Cell Physiology, 55, 1669–1678.25016610 10.1093/pcp/pcu 098 · doi ↗ · pubmed ↗
- 7Chincinska, I. , Gier, K. , Krügel, U. , Liesche, J. , He, H. , Grimm, B. et al. (2013) Photoperiodic regulation of the sucrose transporter St SUT 4 affects the expression of circadian‐regulated genes and ethylene production. Frontiers in Plant Science, 4, 26.23429841 10.3389/fpls.2013.00026 PMC 3576705 · doi ↗ · pubmed ↗
- 8Chincinska, I.A. , Liesche, J. , Krugel, U. , Michalska, J. , Geigenberger, P. , Grimm, B. et al. (2008) Sucrose transporter St SUT 4 from potato affects flowering, tuberization, and shade avoidance response. Plant Physiology, 146, 515–528.18083796 10.1104/pp.107.112334 PMC 2245842 · doi ↗ · pubmed ↗