Novel pennate diatom symbionts support high N2 fixation rates
Bhavya S Panthalil, Angela Vogts, Mar Benavides, Matthew J Harke, Christiane Hassenrück, Ajit Subramaniam, Joseph P Montoya, Maren Voss

TL;DR
A new type of diatom with nitrogen-fixing symbionts may significantly contribute to nitrogen availability in the ocean.
Contribution
Discovery of active nitrogen fixation in pennate diatoms symbiotic with rhizobial and cyanobacterial partners in the Amazon River plume region.
Findings
Pennate diatoms like Mastogloia and Haslea host rhizobial and cyanobacterial symbionts capable of high N2 fixation rates.
NanoSIMS analysis confirmed significant nitrogen fixation in these diatom-symbiont associations.
This symbiosis may represent a new and unquantified source of biologically available nitrogen in oligotrophic oceans.
Abstract
Diazotrophy is the most important nitrogen source in the oligotrophic surface ocean, but the organisms involved and their contributions are incompletely understood due to limited observations. Only diazotrophic organisms possess the nifH gene to reduce dinitrogen to ammonium, but their distribution and activity can only be quantified through sampling and experiments during research cruises. Some recent studies document small diatoms with symbionts able to fix nitrogen, a new source of biologically available nitrogen in addition to the well-known cyanobacterial species such as Trichodesmium or symbionts of haptophytes (UCYN-A) and diatoms (Diatom–Diazotroph Associations, or DDAs). Here, we document a very active symbiosis between small pennate diatoms such as Mastogloia and Haslea with rhizobial and cyanobacterial symbionts in waters of the Western tropical North Atlantic influenced by…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
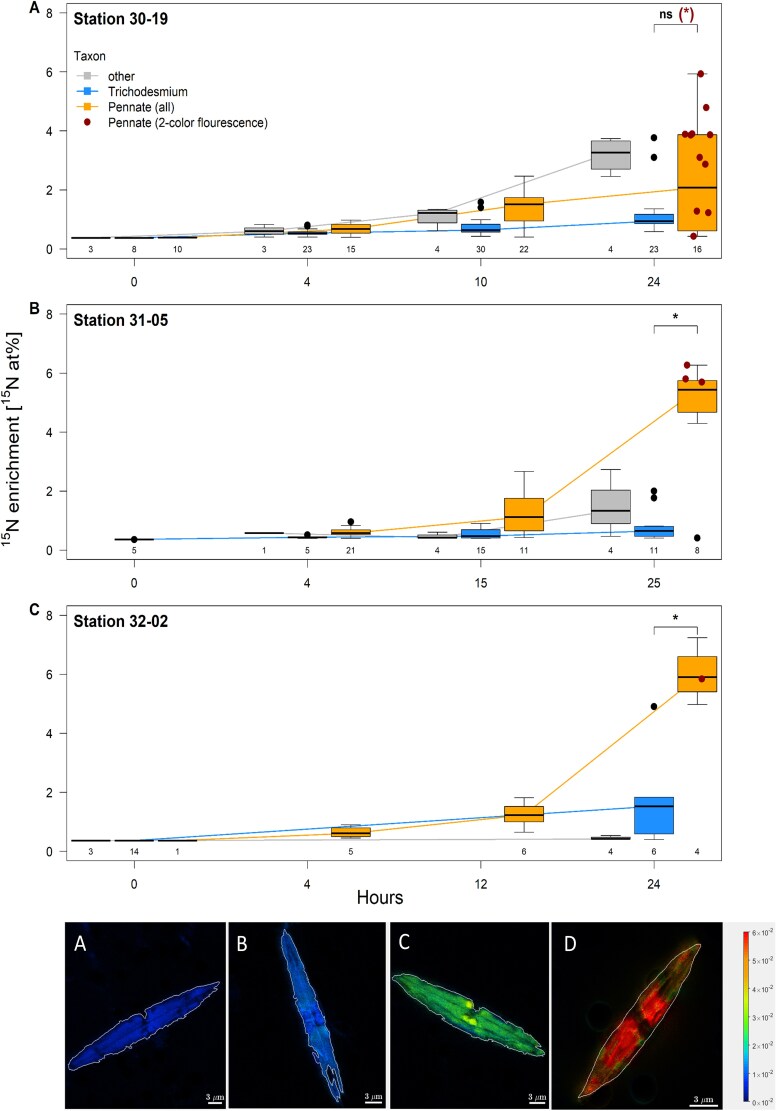 Figure 1
Figure 1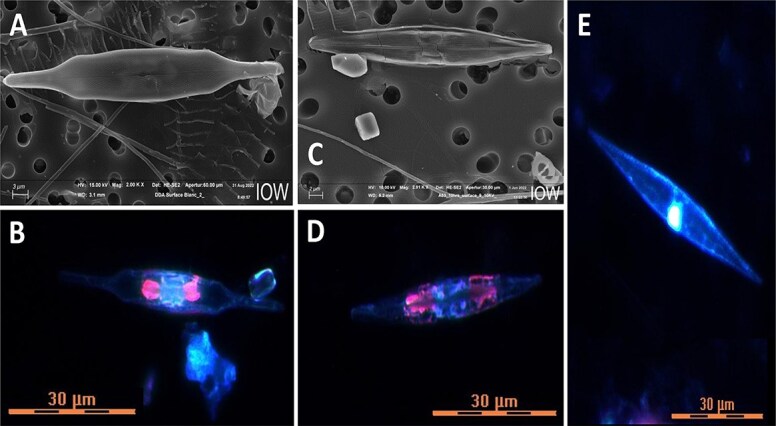 Figure 2
Figure 2- —MeNARP Project (Metabolism of Nitrogen in the Amazon River Plume project grant VO 487/14–1)
- —M174 N-Amazon
- —German Federal Ministry of Education and Research
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Community Ecology and Physiology · Diatoms and Algae Research
Dinitrogen (N_2_) fixation plays a key role in supplying reactive nitrogen to pelagic ecosystems, and filamentous cyanobacteria were long thought to be the only marine diazotrophs. In recent years, the application of fluorescence analysis, molecular and bioinformatic approaches, and single-cell mass spectrometry has generated increasing evidence for a significant contribution of new nitrogen by other groups including non-cyanobacterial diazotrophs [1] and newly discovered endosymbionts of haptophytes [2] and diatoms [3, 4]. Interestingly, these latter diazotrophs could represent an early stage of organellogenesis [5].
Among the enigmatic marine diazotrophs are spheroid bodies containing N_2_ fixation genes [6] in rhopalodiacean diatoms such as Rhopalodia gibba [7]. Similarly, Nakayama [8] found non-photosynthetic intracellular cyanobacteria in the rhopalodiacean Epithemia turgida [9], and N_2_ fixation has been documented in other rhopalodiaceans [4]. More recently, Tschitschko et al. [3] have described a rhizobial symbiont, Candidatus Tectiglobus diatomicula, of biraphid diatoms belonging to the genus Haslea. The rhizobial symbiont appears to be broadly distributed in the world ocean [3], but its host specificity and activity remain poorly characterized. Taken together, these different lines of evidence suggest an overlooked contribution of symbiosis between diatoms and multiple diazotroph lineages to nitrogen supply in the ocean.
Here, we report unexpectedly high ^15^N enrichment within pennate diatoms tentatively identified as Mastogloia spp. and Haslea spp. from ^15^N_2_—tracer experiments carried out in the northern Amazon River Plume (Figs 1 and 2, Supplementary Fig. S1, Supplementary Tables S1–S3). We could not visualize the rhizobial symbionts reported from of Haslea [3] and Epithemia [4], but many of the diatoms we observed contained symbionts with phycoerythrin fluorescence flanking the nucleus axially, in contrast to the lateral arrangement of the previously described rhizobial symbionts (Fig. 2, Supplementary Fig. S3). Our Nano-SIMS analyses of ^15^N in individual diatoms provide definitive evidence of direct assimilation of ^15^N_2_ by cells containing these endosymbionts (Fig. 1).
Although pennate diatoms are known to acquire reactive nitrogen from sympatric, free-living diazotrophs [4], this pathway cannot generate ^15^N enrichments greater than those in the free-living diazotrophs. In all, we identified 108 pennate diatoms that became enriched in ^15^N, of which 56 contained two-color fluorescent inclusions indicative of cyanobacteria (Supplementary Table S4). At the end of our incubations, many pennate diatoms had significantly higher ^15^N enrichments than Trichodesmium and other Diatom-Diazotroph Associations previously known to supply reactive nitrogen to the Amazon River Plume [9–11] (Fig. 1, Supplementary Tables S4–S6).
The high ^15^N enrichment of the pennate diatoms with fluorescent symbionts implies that they were not acquiring ^15^N secondarily from other diazotrophs via a recycling pathway. As a quantitative test, we estimated that the maximum possible enrichment of ^15^N in pennates via release of labeled ^15^N-NH_4_^+^ by other active diazotrophs in the surrounding water is ~1.15 at% ^15^N, a third of the median enrichment of 3.89 at% (n = 28) we found in pennates after 24 h of incubation (SI-Methods). The ^15^N enrichment of pennate diatoms with symbionts increased through our 24 h incubations to as high as 7.24 at% ^15^N (Fig. 1, Supplementary Table S4), while the significantly lower ^15^N enrichments of pennates without symbionts are consistent with acquisition of ^15^N via a recycling pathway (Dunn’s test, z = 4.12, P < .001).
Because these findings were serendipitous, we lack molecular analyses allowing us to unambiguously resolve the identity of these fluorescent endosymbionts. However, we collected samples for metatranscriptomics and interrogated them for community-level signals of nifH gene transcription as well as diatom taxonomy to explore the potential identity of both symbionts and hosts. For taxonomic assessment, Kaiju, a protein-level classification tool [12], could classify between 13.6% and 45.7% of metatranscriptomic reads (Supplementary Table S3) revealing relatively similar diatom communities across the sampled region with site-by-site variability (Supplementary Fig. S4).
Among the diatoms, transcripts attributed to Mastogloia spp. occurred at all stations with the highest relative abundance at station 29-04 (Supplementary Fig. S4B), while transcripts attributed to Haslea spp. were highest at station 31-05 (Supplementary Fig. S4B), suggesting broad presence but relatively low contribution of these pennates to the diatom community. Pennate diatoms of these genera benefit from elevated silicate concentrations e.g. in the river plumes like that of the Amazon (Supplementary Table S1) [11, 13] and may be observed elsewhere.
Although we could not identify the symbionts of our diatoms unambiguously, an evaluation of putative nifH gene expression within the assemblage using a combination of blast searches versus curated and public databases revealed the presence of the rhizobial symbiont, Candidatus Tectiglobus diatomicola, in our samples despite its absence in our microscopic observations (Supplementary Fig. S5). Overall, the highest nifH transcript proportion was observed at Station 28-01 for transcripts attributed to Trichodesmium (3.84 TPM), while transcripts belonging to the recently identified rhizobial symbiont (Candidatus Tectiglobus diatomicola, [3]) showed the highest expression at Station 29-04 (1.87 TPM; Supplementary Fig. S5). Our samples also contained transcripts attributed to Richelia, Crocosphaera, UCYN-A, and an unclassified cyanobacterium (Supplementary Fig. S5). Although we cannot yet identify the inclusions associated with high rates of assimilation of ^15^N by pennate diatoms, they have morphological and fluorescence characteristics of cyanobacteria. Our methods did not allow us to visualize the previously described rhizobial symbionts of Haslea, but our molecular data show them to be present (Fig. S5). Our observations thus provide strong evidence that pennate diatoms may form symbiotic associations with both rhizobia and currently unidentified cyanobacteria.
In summary, multiple broadly distributed diatom genera may be associated with rhizobial and/or cyanobacterial symbionts, but the small size of the pennate diatom host and the challenge of characterizing Diatom–Rhizobial Associations (DRAs) and some Diatom–Cyanobacterial Associations (DCAs) by light or fluorescence microscopy has led to them being largely overlooked. Pennate diatoms are abundant and broadly distributed but whether they play an important role in supplying new nitrogen to tropical waters of the North Atlantic cannot yet be assessed. Further field studies are needed to explore the possibility that DRAs and DCAs with pennate diatom hosts are active in other ocean basins, and to quantify their overall contribution to the oceanic nitrogen budget.
Supplementary Material
Panthalil_Vogts_ISME_Comm_SI_final_ycaf190
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Turk-Kubo KA, Gradoville MR, Cheung S. et al. Non-cyanobacterial diazotrophs: global diversity, distribution, ecophysiology, and activity in marine waters. FEMS Microbiol Rev 2022;47:25. 10.1093/femsre/fuac 046PMC 1071906836416813 · doi ↗ · pubmed ↗
- 2Thompson AW, Foster RA, Krupke A. et al. Unicellular cyanobacterium symbiotic with a single-celled eukaryotic alga. Science. 2012;337:1546–50. 10.1126/science.122270022997339 · doi ↗ · pubmed ↗
- 3Tschitschko B, Esti M, Philippi M. et al. Rhizobia–diatom symbiosis fixes missing nitrogen in the ocean. Nature. 2024;630:899–904. 10.1038/s 41586-024-07495-w 38723661 PMC 11208148 · doi ↗ · pubmed ↗
- 4Schvarcz CR, Wilson ST, Caffin M. et al. Overlooked and widespread pennate diatom-diazotroph symbioses in the sea. Nat Commun 2022;13:799. 10.1038/s 41467-022-28065-635145076 PMC 8831587 · doi ↗ · pubmed ↗
- 5Nakayama T, Inagaki Y. Genomic divergence within non-photosynthetic cyanobacterial endosymbionts in rhopalodiacean diatoms. Sci Rep 2017;7:13075. 10.1038/s 41598-017-13578-829026213 PMC 5638926 · doi ↗ · pubmed ↗
- 6Prechtl J, Kneip C, Lockhart P. et al. Intracellular spheroid bodies of Rhopalodia gibba have nitrogen-fixing apparatus of cyanobacterial origin. Mol Biol Evol 2004;21:1477–81. 10.1093/molbev/msh 08614963089 · doi ↗ · pubmed ↗
- 7Geitler L . On the life history of the epithemiaceae epithemia, rhopalodia and denticula (diatomophyceae) and their presumably symbiotic sphaeroid bodies. Plant Syst Evol 1977;128:259–75. 10.1007/BF 00984562 · doi ↗
- 8Nakayama T, Kamikawa R, Tanifuji G. et al. Complete genome of a nonphotosynthetic cyanobacterium in a diatom reveals recent adaptations to an intracellular lifestyle. Proc Natl Acad Sci 2014;111:11407–12. 10.1073/pnas.140522211125049384 PMC 4128115 · doi ↗ · pubmed ↗