Vaccinium bidoupense (Ericaceae), a new species from Vietnam and Laos, with notes on V. petelotii and V. pseudobullatum

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2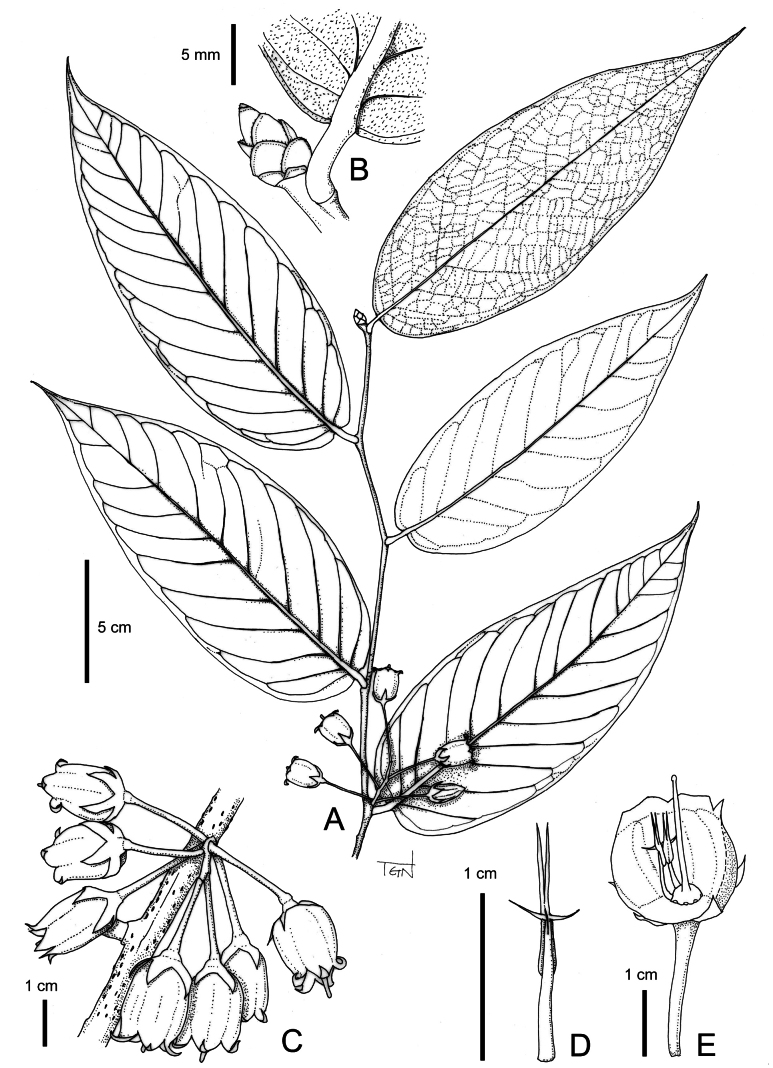 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6| Characters |
|
|
|
|---|---|---|---|
| References | this study | Dunn (1903), | |
| Leaf blade: shape | ovate-elliptic to lanceolate | oblong or narrowly | elliptic to ovate |
| Leaf blade: tertiary veins | prominent in submarginal area, stout | prominent throughout, slender | prominent throughout, stout |
| Petiole: length (mm) | 3–12 | 3–6 | 6–8 |
| Pedicel: length (mm) | 7–14 | 9–15 | 4–7 |
| Calyx: lobe length (mm) | 5–6 | 2.5–3 | 4–6 |
| Corolla: color | pale red to salmon pink | pale green (usually described as cream-colored) | red, pink or white |
| Corolla: length of tube (mm) | 11–13.5 | 3.5–4.5 | 8–9 |
| Stamen: length (mm) | 14–16 | 5–6.5 | 6.5–7 |
| Stamen: filament length (mm) | 8–9 | 2.5–3 | (1–)2–3(–4) |
| Stamen: length ratio of anther tubules to thecae | ca. 1.3 | 1.5–2 | 1.1–1.3(–3) |
| Fruiting pedicel | straight | curved | curved |
| Distribution | Laos (Sekong), S Vietnam (Kon Tum, Lam Dong, Quang Nam) | S China (SE Yunnan), N Vietnam (Lai Chau, Lao Cai) | S China (SE Yunnan, SW Guangxi), N Vietnam (Cao Bang, Ha Giang) |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Plant and Fungal Species Descriptions · Ginger and Zingiberaceae research
Introduction
Vaccinium L. is the largest genus of the tribe Vaccinieae in Ericaceae (Fang 1991; Fang and Stevens 2005; Vander Kloet and Dickinson 2009). The genus has a nearly cosmopolitan distribution and comprises about 450–500 species. Recent phylogenetic studies have demonstrated that Vaccinium and many of its traditionally defined sections are non-monophyletic (Kron et al. 2002; Becker et al. 2024). These findings, however, have not yet been reflected in the corresponding taxonomic rearrangements, and denser sampling is needed before a comprehensive reclassification of the tribe can be attempted (Becker et al. 2024). Pending updates of the formal classification based on phylogenetics, we here adopt the current circumscription and subdivision of Vaccinium.
In Eastern Indochina, 31 species of Vaccinium have been recorded, all of which are known from Vietnam. Of these, 20 species were listed by Nguyen and Ho (1996) and Nguyen (2003), whereas the illustrated guide by Ho (1999) comprised 21 species through the inclusion of V. delavayi Franch. Subsequently, 10 species of Vaccinium were added to the known flora of Vietnam (Vander Kloet and Paterson 2000; Nguyen et al. 2008; Hoang et al. 2016; Hummer et al. 2017; Averyanov et al. 2020; Vu-Huynh et al. 2020; Tong et al. 2022). Three species are known from Laos (Newman et al. 2007), and two species from Cambodia (Cho et al. 2016).
Here, we describe a species of Vaccinium new to science based on our original gatherings, as well as historical collections, made in the Central Highlands of Vietnam and in southern Laos. To facilitate the taxonomic assessment of the new species, we revise the morphological variation of its two similar species, the Asian V. petelotii Merr. and V. pseudobullatum W.P.Fang & Z.H.Pan and rectify the contradictions in the understanding of their floral structure. Finally, we discuss the circumscription of Vaccinium sect. Calcicolus Kloet, which was earlier proposed to include the species treated here.
Materials and methods
Specimens of Vaccinium (or their images) kept in the following herbaria were examined: A, BRIT, DLU, FOF, FU, GXMG, GXMI, HITBC, HN, IBK, IBSC, K, KAG, KUN, LBG, LE, MO, MW, P, PE, SWFC, SZ, UC, US, VNM and WUK (herbarium acronyms follow Thiers, updated continuously). Small plant parts were observed with a hand magnifier. Terminology follows that of Fang and Stevens (2005), Beentje (2016) and Nuraliev et al. (2020). In each list of the examined specimens, the collections are arranged geographically (from NW to SE), and then chronologically. Distribution of the studied taxa in Vietnam is given in accordance with the official administrative division of the country as provided by the Vietnam Administrative Atlas (2015).
Taxonomic treatment
Vaccinium
petelotii
Taxon classificationPlantaeEricalesEricaceae
Merr., Univ. Calif. Publ. Bot. 13(6): 138 (1926).
5F42E032-4870-55BF-BAC8-728F76B5FA96
= Agapetes parviflora Dunn, J. Linn. Soc., Bot. 35: 515 (1903). Type: China. Yunnan Province • Meng-tze [Mengzi County], A. Henry 10488A [fr.] (lectotype, designated by Sleumer 1941: 446, US, 00116973, image!).
Type.
Vietnam. Lao Cai Province • Chapa [Sapa], 1500 m a.s.l., April 1925, P.A. Pételot 1772 [young fr.] (holotype: UC, UC259692, image!; isotypes: A, 00015962, image!; P, P00647843, image!; US, 00116935, image!).
Notes.
Based on the specimens cited in Appendix 1, we have clarified the morphological variation of the species as follows:
In the treatments by Fang (1991) and Fang and Stevens (2005), Vaccinium petelotii is described as having corollas 6–11 mm long. However, the specimens of this species examined in our study bear corollas 4–5 mm long, consistent with the description by, e.g., Huang and Fang (1991) who indicated corollas about 6 mm long. At the same time, the specimen P.I Mao 82-172 (SWFC, 00004475 & 00004478) from Yunnan assigned to V. petelotii by R.C. Fang in 1984 demonstrates remarkably larger flowers with corollas up to 12 mm long. We suppose that the indications of large corollas were based solely on this specimen. Apart from flower size, this specimen differs from typical V. petelotii in larger leaves and shorter pedicels that remain straight (rather than becoming curved) when fruiting. We therefore exclude this specimen from V. petelotii and consider it a possible hybrid between V. petelotii and V. pseudobullatum. Fang and Stevens (2005) indicated the stamen filaments of Vaccinium petelotii as being 0.5 mm long, in which case the anthers would seem to be nearly sessile. We could not detect any reason for this indication and consider it erroneous because the specimens of this species studied here uniformly have filaments 2.5–3 mm long. The ratio of anther tubules to thecae was indicated by Fang and Stevens (2005) as ca. 3 for Vaccinium petelotii, which contradicts our observations (1.6–2.0) as well as the treatments by Fang (1991) and Huang and Fang (1991), both of which state a ratio of 1.5.
Distribution.
S China (SE Yunnan, SW Guangxi), N Vietnam (Lai Chau, Lao Cai).
Vaccinium petelotii. A. Branch with leaves and flowers; B. Leaf blade, abaxial view; C. Inflorescence, lateral view; D. Post-anthetic calyx and disk; E–G. Flower, lateral, oblique and apical view; H. Longitudinal section of flower; I. Flower with perianth removed to show androecium and pistil; J. Stamen, lateral view; K. Stamen, dorsal view. Photos by M.S. Nuraliev, based on M.S. Nuraliev 2359.
Vaccinium
pseudobullatum
Taxon classificationPlantaeEricalesEricaceae
W.P.Fang & Z.H.Pan, Acta Phytotax. Sin. 19(1): 108 (1981).
6103E0F1-F5D7-58A7-BF59-3E10CA25EDD5
Type.
China. Yunnan Province • Malipo County, Tiechang, 1200 m a.s.l., 20 February 1940, C.W. Wang 86988 [fl.] (holotype: PE; isotypes: IBK, IBK00302352!; IBSC, 0457816!; KUN, 12088909! & 12088910!; SZ, 00126560!; WUK, 0268189, image!).
Notes.
Based on the specimens cited in Appendix 1, we have clarified two androecial features of the species:
The length of the stamen filaments was indicated in the protologue (Fang and Pan 1981) as 1–1.5 mm, which was repeated in Fang and Stevens (2005). Fang (1991) and Huang and Fang (1991) considered the range as 1–2 mm. In most of the specimens studied here, however, the filaments are 3–4 mm long. Both flowering gatherings listed by Fang and Pan (1981; Wang 86988 and Feng 13490) bear flower buds (and no fully open flowers), and we suppose it to be the reason that they indicate shorter filaments than those observed by us. Their description was likely reproduced in the subsequent account without additional investigation of flower structure. The ratio of anther tubules to thecae was uniformly indicated as 2–3 for this species (Fang 1991; Huang and Fang 1991, including fig. 105 (9, 10); Fang and Stevens 2005), whereas we observed it to be mostly 1.1–1.3. We therefore treat this feature as more variable than earlier thought.
Distribution.
S China (SE Yunnan, SW Guangxi), N Vietnam (Cao Bang, Ha Giang). Until recently, the species was considered to be endemic to Yunnan; it was reported from Guangxi by Wei (2023) and from Vietnam by Averyanov et al. (2020).
Vaccinium pseudobullatum. A. Branch with leaves and flowers; B. Inflorescence, lateral view; C. Inflorescence, with front view of flowers; D. Bract and two bracteoles; E. Flower, lateral view; F. Androecium; G. Antepetalous (left) and antesepalous (right) stamens, dorsal view; H. Stamen, ventral view I. Three calyx lobes, ovary, disk and style; J. Cross section of ovary. Photos by Y.H. Tong, based on B.M. Wang TYH-965A.
Vaccinium
bidoupense
Taxon classificationPlantaeEricalesEricaceae
Smitinand ex Y.H.Tong, N.T.T.Huong & Tagane sp. nov.
FB85C397-5D32-5E61-BC6F-9A9F2F59C461
urn:lsid:ipni.org:names:77371640-1
Type.
Vietnam. Kon Tum Province • NW slopes of Ngoc Linh Mountain system at 1800–1900 m a.s.l., open places in primary evergreen dense mountain forest, 23 February 1995, L.V. Averyanov, N.T. Ban, N.Q. Binh, A. Budantzev, L. Budantzev, N.T. Hiep, D.D. Huyen, P.K. Loc, N.X. Tam, G. Yakovlev VH053 [fl.] (holotype: HN!; isotypes: BRIT, BRIT402341!; LE, LE01041972!; MO, 5168102!; P, P04485557! & P04483084!).
Diagnosis.
Vaccinium bidoupense is morphologically similar to V. petelotii and V. pseudobullatum in its relatively large leaf blades and umbel-like inflorescence with short rachis. It differs from V. petelotii in having longer calyx lobes (5–6 mm vs. 2.5–3 mm), pale red to salmon pink (vs. pale green) corolla, a longer corolla tube (11–13.5 mm vs. 3.5–4.5 mm), and longer stamens (14–16 mm vs. 5–6.5 mm). The new species differs from V. pseudobullatum in having leaf blades with somewhat less obvious tertiary veins, and longer pedicels (7–14 mm vs. 4–7 mm), corolla tube (11–13.5 mm vs. 8–9 mm) and stamen filaments (8–9 mm vs. 1–4 mm) (Table 1).
Vaccinium bidoupense. A. Flowering branch; B. Bud and base of leaf (abaxial side) showing dense appressed trichomes; C. Inflorescence; D. Stamen, dorsal view; E. Flower with part of corolla and some stamens removed to show disk and style. Drawn by S. Tagane.
Table 1.: A comparison of Vaccinium bidoupense, V. petelotii and V. pseudobullatum. The data on the latter two species were obtained from the cited literature sources and the examined specimens listed in Appendix 1.
Description.
Shrubs, epiphytic or epilithic, evergreen; shoots pendent, ca. 2–5(–10) m long, sometimes rooting at nodes. Branches more or less angled, glabrous, densely lenticellate. Leaves non-distichously alternate (but leaves of a branch often arranged in one plane due to twisted petioles), evenly scattered. Petiole 0.3–1.2 cm long, glabrous. Leaf blade ovate to lanceolate or sometimes elliptic, 12–22 × 4.7–11.7 cm, 2.0–3.7 times as long as wide, coriaceous; adaxial surface green, glabrous; abaxial surface pale green, densely covered with appressed dark brown trichomes (trichomes caducous, leaving faintly punctate scars); base rounded, subtruncate or shallowly cordate, rarely cuneate or prominently cordate; margin entire, slightly revolute, with 1 basal gland per side, glands 1–1.5 mm in diam.; apex acute to acuminate or shortly caudate; median vein and secondary veins impressed adaxially, strongly raised abaxially; secondary veins pinnately arranged, 10–14 on each side of median vein, usually meeting in a closed loop and forming an intramarginal vein; tertiary veins more prominent in submarginal area than in the rest of leaf blade, stout, reticulate, slightly impressed or sometimes flat adaxially, slightly raised abaxially. Inflorescences axillary, 1 per axil, on leafy shoots and older leafless branches, sessile (peduncle absent), racemose, umbel-like, determinate (developing mostly within the confines of the perennating buds), 4–10-flowered; rachis pale green, 0.5–2 cm long, glabrous; bracts caducous, crimson, ovate, 1.0–1.5 × 0.5–0.6 cm, thinly coriaceous, glabrous, margin entire, apex obtuse. Flowers pendent, articulated with pedicels, 5-merous. Pedicels red to pale green, 0.7–1.4 cm long, glabrous; bracteoles 2, attached at base of pedicel, subopposite, acicular, early caducous. Calyx (excluding ovary) pale red to salmon pink, becoming greenish with age, at base appressed to corolla and distally diverging to spreading, glabrous on both sides; tube ca. 1 mm long; lobes 5, triangular-lanceolate, 5–6 × 2–3 mm, distinctly 1-veined. Corolla pale red to salmon pink, glabrous outside and inside; tube cupulate, 11–13.5 × 13–14 mm; lobes 5, strongly recurved, triangular to broadly triangular, 3–4 × 4–5 mm. Stamens 10, dimorphic with respect to spurs, included in corolla, free from each other, 14–16 mm long; filaments white to pale pink, slightly incurved, 8–9 mm long, glabrous in proximal half and puberulent in distal half; anthers tightly appressed to style, golden yellow, dorsifixed, 7–9 mm long, each with 2 spurs; thecae 3–3.5 mm long, slightly papillate; tubules parallel, 4–6 mm long (ca. 1.3 times as long as thecae), opening by oblique ventral apical pores; spurs borne dorsally at base of tubules, strongly curved, ca. 1 mm long, those on antesepalous stamens extending laterally outside of antepetalous anthers and overlapping with spurs of next antesepalous stamens, those on antepetalous stamens strongly hooked outward below spurs of antesepalous stamens. Ovary inferior, pale red to salmon pink, obconic, terete, 1–2 × 2–2.5 mm, glabrous, pseudo-10-locular; disk pale yellow, annular, broadly dome-shaped, ca. 2.5 mm diam., glabrous; style 1.4–1.6 cm long, exserted from corolla for 2.5–4 mm, linear, glabrous; stigma truncate. Infructescences with rachis 1–3 cm long. Fruiting pedicel 1.2–2.4 cm long. Fruit (when young) globose, ca. 6 mm in diam., glabrous, with persistent calyx lobes 5–6 mm long. Mature fruits and seeds unknown.
Vaccinium bidoupense. A. Flowering branch; B–D. Inflorescences; E. Flower, apical view. Photos by L.V. Averyanov, based on Averyanov et al. AL2537.
Ecology and phenology.
Vaccinium bidoupense grows as an epiphyte on tree trunks or as a lithophyte on mossy rocks in broad-leaved and mixed forests at elevations of 1100–2200 m. It flowers from October to February; young fruits are documented from May to June.
Distribution.
Vaccinium bidoupense is known from Laos (Sekong Province) and Vietnam (provinces Kon Tum, Lam Dong, and Quang Nam). The species is therefore endemic to the area comprising the Central Highlands of Vietnam (also called Tay Nguyen Plateau) and their extension in Laos known as Dak Cheung (or Dakchung) Plateau, being broadly distributed across this region. The region comprises three main mountainous areas, i.e., Dak Lak Plateau, Kon Tum—Gia Lai Plateau and Langbian Plateau (Poyarkov et al. 2021), and V. bidoupense is documented to inhabit the latter two areas. As the known locations suggest, the species probably occurs throughout the Central Highlands, and its presence in the provinces Dak Lak, Dak Nong and Gia Lai is especially probable.
Vaccinium bidoupense. A. Habit; B. Leafy branch; C. Leaf blade, abaxial view; D. Base of leaf, abaxial view, showing appressed dark brown trichomes; E. Young leafy branch. Photos by S. Tagane, A. unvouchered; B. based on Toyama et al. V1857; C–E. based on Tagane et al. V4287.
Etymology and history of documentation of Vaccinium
bidoupense.
The species name “Vaccinium bidoupense” was initially proposed by Tem Smitinand on the determination slips of three of Poilane’s specimens kept in P (30292, 30763 and 30909, included below as paratypes of the species). Two of these specimens originated from the Bidoup Massif (and 30292 was collected nearby), which is reflected in the species epithet. Although such a name has never been formally published until now, it was used by Vander Kloet and Avery (2007) (as “V. bidoupense Smithin”) who included the specimen Poilane 30763 in a morphophenetic analysis of stamen characters in the tribe Vaccinieae. Here, we follow Smitinand’s choice of epithet to ensure the clearness of the species identity and to acknowledge his recognition of this distinct species.
Vaccinium bidoupense. A, B. Inflorescences; C. Flower with a half of perianth removed; D, E. Androecium. Photos by S. Tagane, based on Tagane et al. V4287.
Conservation status.
Vaccinium bidoupense is an integral element of highland vegetation of the Central Highlands of Vietnam (including their extension in Laos) with an expected total extent of occurrence (EOO) of about 55,000 km^2^, which does not fit any of the threatened IUCN Red List conservation categories (IUCN, Standards and Petitions Committee 2025). In intact habitats of montane forests, the species meets no damage factors. However, the vast degradation of primary vegetation across the distribution area of the species, including the legally protected territories, leads to a continuing decline in the extent of its occurrence, quality of habitat, number of populations, and number of mature individuals. In this connection, we preliminarily assess V. bidoupense as Near Threatened (NT); we also consider that the species is close to being qualified as Vulnerable (VU) in the near future.
Additional specimens examined (paratypes).
Laos. Sekong Province • [Dak Cheung District,] new road along the border with Vietnam, 15°34′36.0′′N, 107°20′11.5′′E, 1224 m a.s.l., 17 July 2021, P. Souladeth, M. Soukhavong, N. Thongphakdee, T. Boutavong 1141 [st.] (FOF, FOF0009576; KAG, KAG181621). Vietnam. Quang Nam Province • Nam Giang District, Song Thanh Nature Reserve, 15°33′07′′N, 107°23′02′′E, 1100 m a.s.l., 6 May 2019, M.S. Nuraliev 2509 [young fr.] (BRIT, BRIT1150627; IBSC; MW, MW0758862, MW0758863 & MW0758864). Lam Dong Province • N of Dalat, 1500 m a.s.l., 2 September 1940, E. Poilane 30292 [young fr.] (L, L.3786302, image; P, P04484682, P04484683, P04484684, P04484685, P05244320 & P05244322, images) • Bidoup Massif, 2200 m a.s.l., 12 October 1940, E. Poilane 30763 [fl.] (L, L.3786303, image; P, P00647856, P04484686, P04484688, P05244702 & P05244703, images) • ibid., 2000 m a.s.l., 14 October 1940, E. Poilane 30909 [fl.] (P, P04484687, P05244317 & P05244321) • Lac Duong District, Da Chay Municipality, 29 km to NE from Dalat City, 12°6′N, 108°39′E, 1900–2000 m a.s.l., 23 March 1997, L.V. Averyanov, N.Q. Binh, P.K. Loc VH3118 [st.] (HN; LE, LE01041973; MO, MO-5172316) • ibid., 12°6′N, 108°39′E, 2150 m a.s.l., 1 May 1997, L.V. Averyanov, N.Q. Binh, N.T. Hiep, P.K. Loc, P. Lowry VH4463 [fr.] (HN; LE, LE01041976; MO, MO2080998; MW, MW1045967; P, P04483084) • Lac Duong District, Bidoup Nui Ba National Park, 12 October 2011, N.H. Xia, J.B. Ni, Y.H. Tong & X.R. Zheng TYH-1070 [st.] (IBSC) • ibid., SE of Giang Ly station, 12°11′07.7′′N, 108°41′14.6′′E, 1509 m a.s.l., 23 December 2012, A.N. Kuznetsov, S.P. Kuznetsova, A.N. Demidova, N.G. Prilepsky 404 [st.] (MW, MW0750311) • ibid., 12°10′21.03′′N, 108°41′49.76′′E, 1504 m a.s.l., 19 November 2014, H. Toyama, S. Tagane, V.S. Dang, H. Nagamasu, A. Naiki, H. Tran, C.J. Yang et al. V1857 [fl.] (FU; VNM) • ibid., 12°11′11.18′′N, 108°42′53.12′′E, 1639 m a.s.l., 23 February 2016, S. Tagane et al. V4287 [fl.] (DLU; FU; VNM) • ibid., 12°11′09.23′′N, 108°42′55.46′′E, 1637 m a.s.l., 22 December 2018, S. Tagane et al. V9614 [fl.] (DLU; FU; KAG, KAG127364) • ibid., 22 April 2019, T. Yahara et al. V9879 [young fr.] (DLU; FU; KAG, KAG182510) • ibid., 12°4.076′N, 108°38.927′E, 2000–2200 m a.s.l., 8 November 2023, L.V. Averyanov, V.C. Nguyen, B.V. Truong, T. Maisak AL2537 [fl.] (LE, LE01253935, photos LE 01124590).
Discussion
Notes on inflorescence diversity in Vaccinium
The entire genus Vaccinium is characterized by open racemose inflorescences except for the species with single axillary flowers (e.g. Stevens et al. 2004; Fang and Stevens 2005). At the same time, the inflorescences vary in the timing and pattern of their development. As highlighted by Vander Kloet and Dickinson (2005), species of the genus fall into two rather distinct groups differing in having indeterminate and determinate racemes. Vander Kloet and Dickinson (2005) defined the two types as follows: in indeterminate inflorescences, the rachis (termed “floral axis” by these authors) develops after the opening of the inflorescence bud, and in determinate inflorescences it develops within the inflorescence bud. This distinction is consistent with the ideas of Claßen-Bockhoff and Bull-Hereñu (2013; see also Claßen-Bockhoff et al. 2021) who considered “inflorescences sensu stricto” (with indeterminate apical meristem, although usually limited in its activity) and “floral units” (with determinate apical meristem that lacks stem cells). We suggest that a feature allowing a stricter discrimination of the two types of racemes and at the same time offering their easier recognition is the emergence of new flowers during the growth of the raceme beyond the confines of the inflorescence bud. In indeterminate racemes, the apical meristem continues to produce new flowers well after the bud opening, which often leads to a long duration of the raceme flowering and to markedly differing phenology of individual flowers along the rachis (including even the co-existence of young fruits at the base of the inflorescence with underdeveloped flower buds at its apex). Conversely, in determinate racemes all the flowers are formed by the time of bud opening. In this case, although the flower buds are sometimes pronouncedly unequal in size at the stage of opening of the inflorescence bud, the flowers function more or less synchronously.
An additional difference between the types of racemes (already highlighted by Vander Kloet and Dickinson 2005) is the tendency of the bracts subtending each pedicel to be leaf-like or scale-like and often small in indeterminate racemes, whereas the determinate racemes usually have elaborate and large bracts that act as scales of showy inflorescence buds. It should be noted that the features that define the two types are not related to the number of flowers per raceme, as this varies greatly within each type.
Although the distinctness of the two types of racemes was confirmed for the species of Vaccinium closely examined so far (e.g., by Vander Kloet and Dickinson 2005, 2009; Nuraliev et al. 2020; Ye et al. 2021; Tong et al. 2022), it is still to be tested on a broader sampling, and an assessment is needed as to the extent of correspondence with the sections of Vaccinium and phylogeny of the tribe Vaccinieae. For the moment, and based on the clarification provided above, we here reconsider the racemes of V. glaucorubrum (C.Y.Wu) Y.H.Tong & P.W.Fritsch described as indeterminate in our earlier study (Tong et al. 2022) as being determinate. In particular, all the flowers are formed (and readily visible) in this species by the time of the opening of the inflorescence bud and are subtended by brightly colored rhomboid bracts ca. as long as the flowers.
Vander Kloet and Dickinson (2005) recognized three groups of species within the tribe Vaccinieae based on their morphophenetic analysis of inflorescence characters. These authors stated that the groups (designated as A, B and C) are largely in agreement with the primary subdivisions (III, II and I, correspondingly) in the polytomic key to the sections of Vaccinium compiled by Sleumer (1941, p. 413). However, the morphological characterizations provided by Vander Kloet and Dickinson (2005) for their groups differ considerably from Sleumer’s couplets; in particular, Sleumer did not mention the indeterminate vs. determinate racemes. Most importantly, we uncovered about a dozen sections of Vaccinium whose placement represent discrepancies between Vander Kloet and Dickinson’s groups and Sleumer’s key.
Taxonomic distinctness of Vaccinium
bidoupense
Vaccinium bidoupense differs from most of its congeners in having the following combination of morphological traits: non-tuberous stem bases or roots, leaf blades with entire margins and without glandular setae on the median vein abaxially, and determinate racemes that are umbel-like from the pedicels being approximately as long as the rachis. It shares this set of features with only two species of the genus, viz., V. petelotii and V. pseudobullatum. Comparison of the three species is provided in the diagnosis. It is remarkable that the close similarity of the new species with V. petelotii was also assumed by Pham Hoang Ho, who identified the P specimens in sched. as “Vaccinium petelotii Merr. var. bidoupense Smitin. ex Pham-Hoang” (this varietal name, however, has never appeared in any publications).
Several collections assigned here to Vaccinium bidoupense (Averyanov et al. VH053, VH3118, VH4463, and Poilane 30909) were earlier identified in sched. as V. dunalianum var. megaphyllum Sleumer. However, V. bidoupense, and in particular the specimens in question, readily differs from the entirety of V. dunalianum Wight (Fang and Stevens 2005) in having umbel-like racemes with the rachis not longer than 2 cm and pedicels 0.7–1.4 cm long (vs. elongate racemes with rachis 3–7 cm long and pedicels 0.5–0.8 cm long) and much longer corolla tubes (11–13.5 mm vs. 3.5–4.5 mm long).
History of Vaccinium
sect. Calcicolus, with special reference to V. bidoupense
When establishing Vaccinium sect. Calcicolus, Vander Kloet and Dickinson (2005) placed eight species in the section, including V. dunalianum and V. petelotii discussed above. Shortly thereafter, Vander Kloet and Avery (2007) listed “V. bidoupense” as belonging to this section. Vander Kloet and Dickinson (2005, 2009) stated that the section is characterized by an evergreen habit, dimorphic perennating buds (the inflorescence buds at least three times as large as the vegetative buds), a determinate inflorescence (developing entirely within the confines of an enlarging perennating bud), large caducous inflorescence bracts, a pseudo-10-locular ovary, 2–5 seeds per locule, and soft seed testa (see also discussion of the section by Nuraliev et al. 2020). Vaccinium pseudobullatum (considered above as one of the two species most similar to V. bidoupense) was not mentioned by Vander Kloet and Dickinson (2005, 2009), nor by Vander Kloet and Avery (2007), but is consistent with Vaccinium sect. Calcicolus morphologically.
Vander Kloet and Dickinson (2009) emphasized the morphological similarity of Vaccinium lanigerum Sleumer to certain species of Vaccinium sect. Calcicolus. Consequently, they united Vaccinium sect. Calcicolus with Vaccinium sect. Pseudocephalanthos C.Y.Wu & R.C.Fang (but erroneously chose the former name for the expanded section, although the latter name has a nomenclatural priority). Vaccinium lanigerum, however, is characterized by swollen stem bases or roots (Y.H. Tong, personal observation in natural habitat), markedly differing in this respect from Vaccinium sect. Calcicolus (see Nuraliev et al. 2020). For this reason, we do not here follow the merger proposed by Vander Kloet and Dickinson (2009).
More recently, Ye et al. (2021) transferred Vaccinium glaucophyllum C.Y.Wu & R.C.Fang to Vaccinium sect. Calcicolus based on its overall morphology, and Tong et al. (2022) segregated V. glaucorubrum from V. gaultheriifolium (Griff.) Hook.f. ex C.B.Clarke, the type species of Vaccinium sect. Calcicolus. Finally, Tong et al. (2021) and Guo et al. (2023) placed their newly described V. motuoense Y.H.Tong & Y.J.Guo and V. usneoides Y.H.Tong, Y.J.Guo & Ting Zhang in the section. Altogether, 14 species have been explicitly placed in Vaccinium sect. Calcicolus. The morphological basis for these placements, however, largely remains questionable, and at the same time more than a dozen Asian species of Vaccinium possess the set of morphological characters outlined above but have not been analyzed with respect to their possible inclusion in Vaccinium sect. Calcicolus. Given all the uncertainties outlined above, we here refrain from assigning V. bidoupense to either Vaccinium sect. Calcicolus or any other formal infrageneric subdivision.
Becker et al. (2024) demonstrated polyphyly of Vaccinium sect. Calcicolus (sensu Vander Kloet et al.): of the four species sampled in their phylogenetic study, the clade V. gaultheriifolium + V. glaucoalbum Hook.f. ex C.B.Clarke is sister to Vaccinium sect. Aethopus Airy Shaw, and the clade V. chunii Merr. ex Sleumer + V. dunalianum groups with the species of Vaccinium sect. Conchophyllum Sleumer and Vaccinium sect. Galeopetalum J.J.Sm. (but Becker et al. 2024 erroneously placed V. glaucoalbum into Vaccinium sect. Galeopetalum at least as based on Vander Kloet and Dickinson 2005). Because no phylogenetic data are available for V. bidoupense, V. petelotii or V. pseudobullatum, it is currently not possible to establish their relationships. Phylogenetic reconstruction with expanded species sampling is strongly needed for comprehensive reassessment of Vaccinium sect. Calcicolus and the other sections of Asian tropical Vaccinium.
Supplementary Material
XML Treatment for Vaccinium petelotii
XML Treatment for Vaccinium pseudobullatum
XML Treatment for Vaccinium bidoupense
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Averyanov LV Nguyen KS Tran TH Averyanova AL Maisak TV Nguyen HT (2020) Plant Diversity, Flora and Vegetation of Bat Dai Son Mountain Area, Northern Vietnam. Strata, Saint-Petersburg, 1–561.
- 2Becker AL Crowl AA Luteyn JL Chanderbali AS Judd WS Manos PS Soltis DE Smith SA Goncalves DJP Dick CW Weaver WN Soltis PS Cellinese N Fritsch PW (2024) A global blueberry phylogeny: Evolution, diversification, and biogeography of Vaccinieae (Ericaceae). Molecular Phylogenetics and Evolution 201: 108202. 10.1016/j.ympev.2024.10820239288897 · doi ↗ · pubmed ↗
- 3Beentje H (2016) The Kew Plant Glossary, Second Edition. Royal Botanic Gardens, Kew, London, 1–184.
- 4Cho SH Chhang P Kim YD (2016) A checklist for the seed plants of Cambodia. National Institute of Biological Resources, Ministry of Environment, Incheon, 1–272.
- 5Claßen-Bockhoff R Bull-Hereñu K (2013) Towards an ontogenetic understanding of inflorescence diversity.Annals of Botany 112(8): 1523–1542. 10.1093/aob/mct 00923445936 PMC 3828942 · doi ↗ · pubmed ↗
- 6Claßen-Bockhoff R Ronse De Craene LP Becker A (2021) From meristems to floral diversity: Developmental options and constraints. Frontiers in Ecology and Evolution 9: 637954. 10.3389/fevo.2021.637954 · doi ↗
- 7Dop P Trochain-Marquès J (1931) Les Vaccinium du groupe Dunalianum en Indo-Chine. Bulletin du Muséum National d’Histoire Naturelle, Ser 2, 3(7): 698–699.
- 8Dunn ST Wright CH (1903) Descriptions of new Chinese plants. Journal of the Linnean Society.Journal of the Linnean Society of London, Botany 35(247): 483–518. 10.1111/j.1095-8339.1903.tb 00699.x · doi ↗