Comparative cytogenetics of the Physalaemus gracilis group (Anura, Leptodactylidae) with characterization of the karyotype of Physalaemus evangelistai Bokermann, 1967
Pedro Henrique Pacheco Mosquini, Lucas Henrique Bonfim Souza, Juan Martín Ferro, Luciana Bolsoni Lourenço

TL;DR
This study compares the karyotypes of several Physalaemus frog species, focusing on the location of NORs and repetitive DNA sequences to assess chromosome homology.
Contribution
The paper provides the first karyotype characterization of Physalaemus evangelistai and expands cytogenetic data for related species.
Findings
Physalaemus evangelistai has a 2n = 22 karyotype with meta- and submetacentric chromosomes.
An interstitial DAPI-positive band on NOR-bearing chromosomes supports homology among Physalaemus species.
Cytogenetic differences were observed between P. carrizorum populations from Palmas-PR and Misiones.
Abstract
The anuran species group Physalaemus gracilis comprises six species, and variation in the location of nucleolus organizer regions (NORs) was observed across the four species that have been karyotyped to date. The NORs are located interstitially on chromosome 8 of P. carrizorum Cardozo et Pereyra, 2018, and P. lisei Braun et Braun, 1977, terminally on chromosome 8 of P. gracilis (Boulenger, 1883), and terminally on chromosome 10 of P. barrioi Bokermann, 1967. To contribute to the comparative analysis of this group, including the assessment of the hypothesis of homology among these NOR-bearing chromosomes, we described the karyotype of P. evangelistai Bokermann, 1967, and expanded the cytogenetic analyses of P. carrizorum, P. lisei, and P. barrioi. We used classical cytogenetic techniques and mapped, by fluorescent in situ hybridization (FISH), two repetitive sequences: the PcP190…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
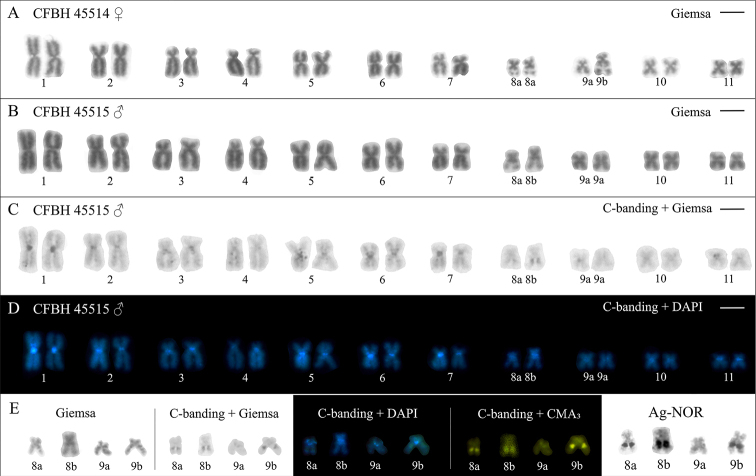 Figure 1
Figure 1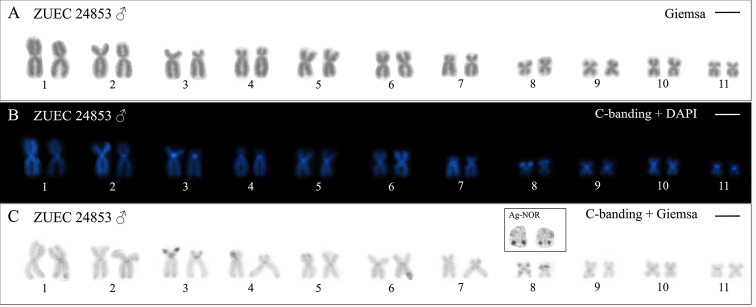 Figure 2
Figure 2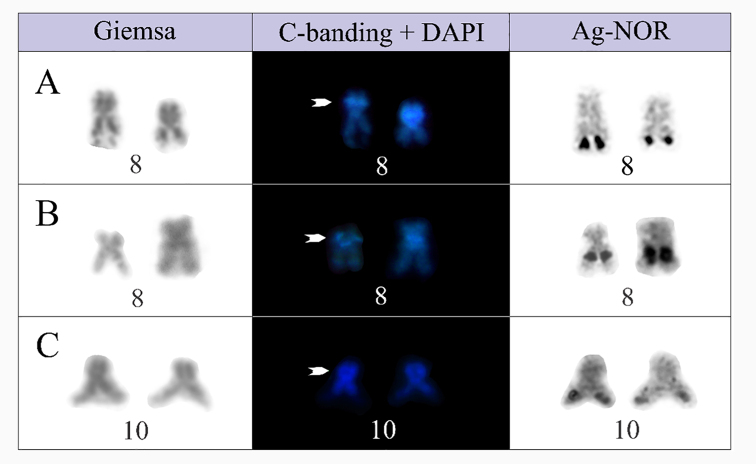 Figure 3
Figure 3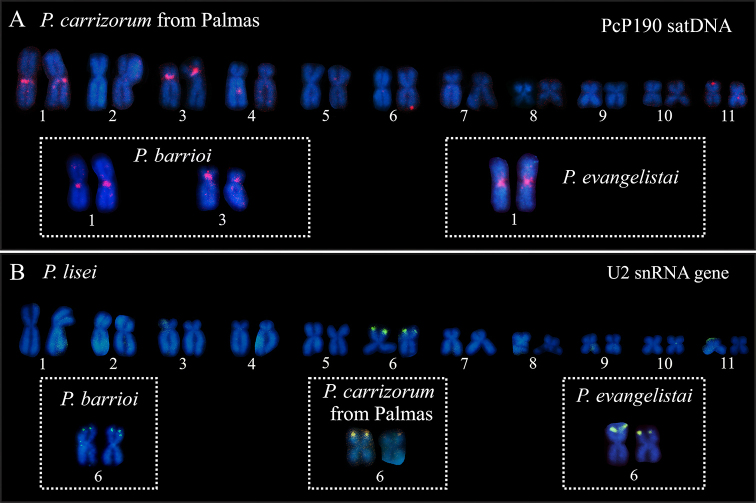 Figure 4
Figure 4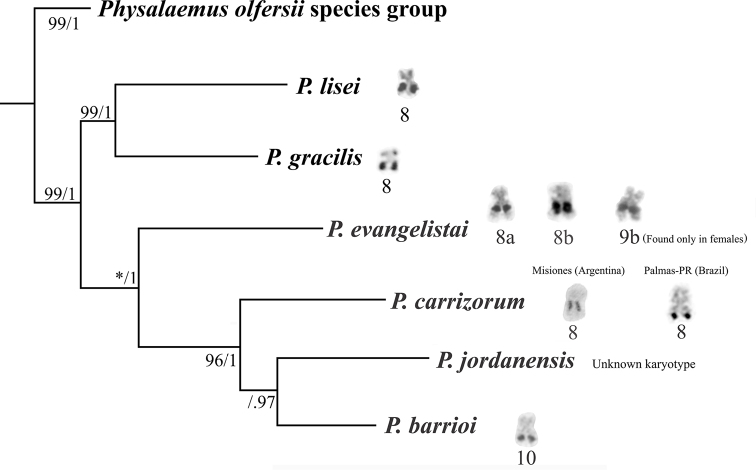 Figure 5
Figure 5| Species | Locality | Voucher number in zoological collection | Chromosome preparation accession number |
|---|---|---|---|
|
| São José do Barreiro, SP, Brasil ( | ZUEC 18146 ♂ | |
| Piraquara, PR, Brasil ( | LGE 15329 | LGE 15329 | |
|
| Palmas, PR, Brasil ( | ZUEC 24848 ♂, ZUEC 24853 ♂ | |
| Arroyo de los Muertos, Misiones, Argentina ( | LGE 24608 ♂ 3 | LGE 24608 3 | |
|
| Salinas, MG, Brasil ( | CFBH 45513 ♂, CFBH 45514 ♀, CFBH 45515 ♂, CFBH 45516 ♂, CFBH 45517 ♂, CFBH 45518 ♀ | |
|
| Gravataí, RS, Brasil ( | LGE 28043 ♂ | |
|
| Gramado, RS, Brasil ( | ZUEC 24003 ♂ |
- —Fundação de Amparo à Pesquisa do Estado de São Paulo 501100001807 https://ror.org/02ddkpn78 http://doi.org/10.13039/501100001807
- —Consejo Nacional de Investigaciones Científicas y Técnicas 501100002923 https://ror.org/03cqe8w59 http://doi.org/10.13039/501100002923
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico 501100003593 https://ror.org/03swz6y49 http://doi.org/10.13039/501100003593
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChromosomal and Genetic Variations · Genetic and Clinical Aspects of Sex Determination and Chromosomal Abnormalities · Genomics and Phylogenetic Studies
Introduction
Karyotype comparisons can be highly informative for distinguishing certain clades, as they provide characters related to diploid number, chromosomal morphology and size, the distribution of repetitive elements, and other traits that may be specific to distinct lineages (Dobigny et al. 2004; Guerra 2008; Mezzasalma et al. 2023). The Neotropical genus Physalaemus Fitzinger, 1826 represents an interesting clade in terms of chromosome evolution, and karyotype data have proven informative for comparing phylogenetically related groups within this genus (see Tomatis et al. 2009; Vittorazzi et al. 2014b; Lourenço et al. 2015; Ferro et al. 2022; Souza et al. 2025 and references therein). In addition, this genus is characterized by a high frequency of chromosomal polymorphisms and a large number of cryptic species (e.g., Barrio 1965; Cassini et al. 2010; Nascimento et al. 2019), which often hinder species identification based solely on morphological characteristics.
The anuran genus Physalaemus, which belongs to the family Leptodactylidae and comprises 50 species, is distributed from southern Venezuela and the Guianas to eastern Bolivia, Paraguay, southeastern Colombia, Uruguay, and a large portion of Brazil (Frost 2025). Phylogenetic inferences divide this genus into two major clades: the Physalaemus signifer clade and the Physalaemus cuvieri clade (Lourenço et al. 2015). The latter clade encompasses five species groups—P. gracilis, P. biligonigerus, P. cuvieri, P. henselii, and P. olfersii—as well as the species Physalaemus cicada Bokermann, 1966, which has not been allocated to any group (Lourenço et al. 2015).
Species of Physalaemus exhibit karyotypes with 2n = 22 and fundamental numbers (FN) of 42 or 44 (see discussion in Tomatis et al. 2009; Lourenço et al. 2015). The karyotypes of all karyotyped species of the P. signifer Clade have FN = 42 and are characterized by the presence of a telocentric chromosome pair 11, which was hypothesized to be a synapomorphy of this clade (Lourenço et al. 2015). Another notable cytogenetic feature in this genus is a heterochromatic band on the short arm of chromosome 5, which may represent a synapomorphy of the Physalaemus cuvieri group (Vittorazzi et al. 2014b; Lourenço et al. 2015).
The Physalaemus gracilis group is composed of six species: P. lisei Braun et Braun, 1977, P. gracilis (Boulenger, 1883), P. evangelistai Bokermann, 1967, P. carrizorum Cardozo et Pereyra, 2018, P. jordanensis Bokermann, 1967, and P. barrioi Bokermann, 1967 (Lourenço et al. 2015; Cardozo and Pereyra 2018). Four out of these six species have had their karyotypes previously described (Brum-Zorrilla and Saez 1968; De Lucca et al. 1974; Provete et al. 2012; Ferro et al. 2022), while the karyotypes of P. evangelistai and P. jordanensis remain unknown. Across the karyotyped species, variation in the position of the nucleolus organizer region (NOR) is observed: it is interstitial on chromosome 8 of P. carrizorum and P. lisei, terminal on chromosome 8 of P. gracilis, and terminal on chromosome 10 of P. barrioi (Brum-Zorrilla and Saez 1968; De Lucca et al. 1974; Provete et al. 2012; Ferro et al. 2022). Given that these NOR-bearing chromosomes are also similar in morphology and size, Ferro et al. (2022) proposed that they are homologous. However, the absence of further cytogenetic markers still prevents a comprehensive evaluation of this hypothesis.
Other interesting chromosomal markers observed in the P. gracilis group are sites enriched with the PcP190 satellite DNA (satDNA) (Vittorazzi et al. 2011), which is a repetitive DNA sequence widely found in anurans, reported in seven families of Hyloidea to date (Vittorazzi et al. 2011, 2014a; Gatto et al. 2016, 2018; Targueta et al. 2018; Gatto et al. 2019). In the analyzed species of the P. gracilis group, Ferro et al. (2022) identified a small number of chromosomal clusters of PcP190 per karyotype, distributed in apparently species-specific patterns. While in P. lisei a cluster of PcP190 was mapped to the (peri)centromeric region of chromosome 7, in P. carrizorum it was found on chromosome 3, and in P. gracilis, on chromosomes 2 and 9 (Ferro et al. 2022).
To enhance our understanding of chromosomal homologies and evolutionary relationships within the P. gracilis group, we described the karyotype of P. evangelistai, presented new data for P. carrizorum from a newly sampled locality, and conducted a comparative analysis of repetitive DNA clusters among the species with available chromosomal data in this species group.
Material and methods
Species, chromosome preparations, and conventional techniques
We analyzed samples of P. barrioi, P. carrizorum, P. evangelistai, P. gracilis, and P. lisei (Table 1). Physalaemus evangelistai specimens and P. carrizorum samples from Palmas, state of Paraná, were collected under the authorization of the Chico Mendes Institute for Biodiversity Conservation/Biodiversity and Information System (ICMBio/SISBIO - permit number 32483). The sampling sites and voucher numbers in scientific collections are provided for all analyzed specimens in Table 1.
Chromosome preparations were obtained from cell suspensions deposited in the scientific collection “Shirlei Maria Recco Pimentel” (SMRP), housed at LabEsC-UNICAMP. These suspensions were previously prepared from intestinal and testicular tissues, following, respectively, the protocols of King and Rofe (1976), with modifications outlined by Gatto et al. (2018), and Schmid (1978), replacing the hypotonic KCl solution with cold distilled water. All experiments were conducted in accordance with relevant guidelines and approved by the Committee for Ethics in Animal Use of the University of Campinas (CEUA/UNICAMP).
Mitotic metaphases were stained with 10% Giemsa, C-banded following Sumner (1972), with modifications described in Siqueira et al. (2008), and silver-impregnated using the Ag-NOR method following Howell and Black (1980). Some C-banded metaphases were sequentially stained with 4’-6 diamidine-2-phenylindole (DAPI, 0.5 µg/mL) and Chromomycin A_3_ (CMA_3_, 0.5 mg/mL) after removal of Giemsa-staining in 50% acetic acid for five minutes. Mitotic metaphases were observed using an Olympus BX-60 microscope (Olympus, Tokyo, Japan). Images were captured with a Quick Start FL-20 camera, using Mosaic 2.4 (Tucsen Photonics, Fuzhou, China). Brightness and contrast adjustments were made using Adobe Photoshop 2021 (Adobe Systems, San Jose, CA, USA).
Isolation and analysis of nucleotide sequences of PcP190 satellite DNA
Genomic DNA samples were obtained from liver tissue of one specimen of P. evangelistai (CFBH 45518) and one specimen of P. lisei (ZUEC 24003). DNA extraction was performed following the TNES method as employed by Medeiros et al. (2013), or alternatively using the QIAamp Fast DNA Tissue Kit (QIAGEN, Hilden, Germany), following the manufacturer’s instructions. The PcP190 satDNA was isolated by PCR using the specific set of primers P190F and P190R (Vittorazzi et al. 2011). The PCR products were purified using the Wizard SV Gel and PCR Clean-Up System (Promega Corporation, Madison, WI, USA), following the manufacturer’s instructions. Samples of the purified products were sequenced by the sequencing facility of the Human Genome and Stem Cell Research Center/IB-USP. The obtained sequences were edited using the BioEdit Sequence Alignment Editor v.7.7.1 (Hall 1999) and compared with sequences available in the NCBI database using BLASTn. After BLASTn indicated similarity between the isolated sequences and those of PcP190 type 1a (see Gatto et al. 2016 for details), we aligned the obtained sequences with all complete PcP190 1a monomers, available in the NCBI database. We excluded monomers derived from P. albonotatus (Steindachner, 1864), which has a 7 bp deletion. The average p-distance value was calculated using the MEGA v.11.0.13 software (Tamura et al. 2021), treating alignment gaps and missing data as pairwise deletions. Alignment figures were generated using Geneious v.7.1.3 (https://www.geneious.com).
Fluorescent in situ Hybridization (FISH) of PcP190 Satellite DNA and U2 snRNA gene
PcP190 satDNA and U2 snRNA gene sequences were obtained from cloned fragments available in the SMRP collection, previously isolated from Physalaemus cuvieri Fitzinger, 1826 (KM361675.1, Vittorazzi et al. [2014a]) and P. ephippifer (Steindachner, 1864) (PQ200500, Souza et al. [2025]), respectively. The PcP190 satDNA was labeled using a PCR Dig Probe Synthesis Kit (Roche) with the universal primers SP6 and T7. The U2 snRNA gene probe was obtained using the primers U2-F and U2-R, designed by Souza et al. (2025). The resulting probes were hybridized with chromosome preparations following the protocol by Viegas-Péquignot (1992). The probes were detected using an anti-digoxigenin-rhodamine antibody (0.06 μg/mL), the chromosomes were counterstained with DAPI (0.5 μg/mL), and the preparations were mounted in Vectashield Antifade Mounting Medium (Vector Laboratories, Burlingame, CA, USA). Microscope analyses and image acquisition were performed as mentioned above.
Results
The karyotype of Physalaemus
evangelistai
The P. evangelistai karyotype had a diploid number of 22 and a fundamental number of 44 (Fig. 1). Chromosome pairs 1, 5, 6, and 10 were metacentric, while pairs 2–4, 7–9, and 11 were submetacentric (Fig. 1A–D). Small heterochromatin blocks were revealed by C-banding in the centromeric region of all chromosomes (Fig. 1C). All these bands were also evidenced by DAPI in metaphases previously subjected to C-banding (Fig. 1D). An interstitial C-band was found on the long arm of chromosome 8, which was DAPI-negative, CMA_3_-positive, and colocalized with the NOR (Figs 1D, E, 3B). In some metaphases, a small C-band was noted on the short arm of chromosome 8, particularly after DAPI staining (Figs 1E, 3A).
Chromosomes of Physalaemus evangelistaiA female karyotype stained with Giemsa B–D male karyotype sequentially subjected to Giemsa-staining (B), conventional C-banding (C), and DAPI-staining (D) E chromosomes 8 and 9 subjected to Giemsa-staining, conventional C-banding, C-banding with DAPI- and CMA₃-stainings, and silver-impregnation using the Ag-NOR method. Scale bars: 5 µm.
In all analyzed specimens, chromosome 8 exhibit an interstitial NOR on the long arm (Figs 1E, 3B), coinciding with a secondary constriction observed in Giemsa-stained metaphases (Figs 1E, 3B). However, variation in NOR size allowed the identification of two morphotypes: 8a, bearing a small NOR, and 8b, carrying a large NOR (Fig. 1E). Three males (CFBH 45513, CFBH 45515, and CFBH 4516) were heterozygous with respect to this condition (8a8b), while one male (CFBH 45517) was homozygous for the smaller NOR (8a8a). Both females (CFBH 45514 and CFBH 45518) were homozygous for the smaller NOR (8a8a).
In addition to chromosome pair 8, one homologue of chromosome pair 9 also carried a NOR in both analyzed females (Fig. 1E), coinciding with a secondary constriction in Giemsa-stained metaphases and being DAPI-negative and CMA_3_-positive (Fig. 1E). The NOR-bearing chromosome 9 (morph 9b) was larger than its counterpart (morph 9a), which lacks a NOR. All four analyzed males had no NOR on this chromosome pair.
Physalaemus carrizorum from Palmas-PR presented a karyotype with FN = 44 and 2n = 22 (Fig. 2A–C). DAPI-positive C-bands were detected in all centromeric regions (Fig. 2B, C). Heterochromatic bands were also present pericentromerically on the short arm of chromosome 3 and on the long arm of chromosome 7, interstitially on the short arm of chromosome 8, and terminally on the short arm of chromosome 3 (Fig. 2B, C). This terminal C-band was larger in one homologue of chromosome pair 3 and more noticeable in C-banded metaphases stained with Giemsa (Fig. 2C). Chromosome 8 had a terminal NOR on the long arm (inset in Figs 2C, 3A), coinciding with a secondary constriction observed in Giemsa-stained metaphases (Figs 2A, 3A). This chromosome also presented a small interstitial C-band on the long arm adjacent to the NOR, barely seen in some metaphases (Fig. 2C).
Karyotype of Physalaemus carrizorum from Palmas-PR A–C Metaphase of male ZUEC 24853 after Giemsa-staining (A), C-banding with DAPI-staining (B), and C-banding with Giemsa-staining (C). In the inset in C, NOR-bearing chromosomes after the Ag-NOR method. Scale bars: 5 µm.
Additionally, the reanalysis of P. barrioi specimens allowed us to identify a faint DAPI-stained C-band on the short arm of the NOR-bearing chromosome 10, in a position similar to that observed in the NOR-bearing chromosomes 8 of P. evangelistai and P. carrizorum (Fig. 3).
NOR-bearing chromosomes of Physalaemus carrizorum (A), P. evangelistai (B), and P. barrioi (C), sequentially subjected to Giemsa-staining, C-banding and DAPI-staining, and the Ag-NOR method. Arrowheads indicate an interstitial DAPI-positive C-band present in all species. Note that, although less evident, it is also present in the NOR-bearing chromosome of P. barrioi.
Chromosomal mapping of PcP190 satDNA and the U2 snRNA gene in the P.
gracilis group
The PcP190 satDNA nucleotide sequences isolated from specimens of P. evangelistai and P. lisei (GenBank accession numbers PV583974 and PV983975, respectively) were 87.5% similar to each other. Based on a comparison with PcP190 sequences available in GenBank, both sequences were assigned to the type 1a of PcP190 satDNA, as they were highly similar to previously described sequences of this type (Suppl. material 1). The mean p-distances between the PcP190 sequences of P. evangelistai and P. lisei and the previously described PcP190 1a sequences, excluding primer regions, were 0.0781 and 0.126, respectively (Suppl. material 1).
Hybridization of the PcP190 satDNA probes revealed clusters of this sequence in the (peri)centromeric regions of the long arm of chromosome 1 and short arm of chromosome 3 in P. carrizorum from Palmas (Fig. 4A) and P. barrioi (inset in Fig. 4A), while in P. evangelistai only the short arm of chromosome 1 exhibited cluster of this sequence (inset in Fig. 4A). Chromosome mapping of the U2 snRNA gene revealed terminal clusters on the short arm of chromosome 6 in P. barrioi, P. carrizorum, P. evangelistai, and P. lisei (Fig. 4B).
Chromosome mapping of PcP190 satDNA (in red) and U2 snRNA gene (in green) in species of the Physalaemus gracilis group A chromosomes of P. carrizorum from Palmas, P. barrioi, and P. evangelistai hybridized with PcP190 probe B mapping of the U2 snRNA gene on chromosomes of P. lisei, P. barrioi, P. carrizorum from Palmas, and P. evangelistai. Chromosomes 1 and 3 of P. barrioi shown in the figure were obtained from different metaphases and subsequently arranged in the figure.
Discussion
The diploid number of P. evangelistai (2n = 22) is consistent with that reported for all other Physalaemus species karyotyped to date (Perkins et al. 2019), including members of the P. gracilis group (Provete et al. 2012; Ferro et al. 2022). Additionally, P. evangelistai has a FN of 44, a trait shared by all cytogenetically analyzed species within the P. cuvieri Clade, except P. fernandezae (Müller, 1926), which has a FN of 42 due to its smallest chromosome pair being telocentric (Tomatis et al. 2009; Lourenço et al. 2015).
All P. evangelistai specimens had a NOR-bearing chromosome pair 8, and an additional NOR was observed on one of the homologues of chromosome pair 9 in the two females analyzed. This additional NOR was not observed in any of the four males analyzed, nor in any other karyotyped species of the P. gracilis group (Provete et al. 2012; Ferro et al. 2022, Fig. 5), which may suggest the presence of a ZZ/ZW sex determination system in this species. NORs have previously been identified as distinguishing markers between sex chromosomes in some anuran species, including Physalaemus ephippifer (Nascimento et al. 2010), Gastrotheca riobambae (Schmid et al. 1983), Buergeria buergeri (Schmid et al. 1993), Hyla femoralis (Schmid and Steinlein 2003; Wiley 2003), and Engystomops petersi (Targueta et al. 2010). In the cases of B. buergeri (Schmid et al. 1993) and H. femoralis (Schmid and Steinlein 2003; Wiley 2003), a NOR was the sole cytogenetic feature differentiating the sex chromosomes, being found on chromosome Z in B. buergeri, colocalized with heterochromatin, and on chromosome X in H. femoralis. On the other hand, it is worth noting that polymorphisms related to the presence/absence of NORs are relatively common among anurans (e.g., Medeiros et al. 2003; Barth et al. 2009; Zaleśna et al. 2017, among several others). Therefore, the limited size of the P. evangelistai sample analyzed here, particularly with respect to females, prevents us from determining whether the observed heteromorphism is sex-linked or represents an autosomal polymorphism. Further studies involving a larger number of individuals would provide valuable insights into this issue.
NOR-bearing chromosomes in the P. gracilis species group. The phylogenetic relationships are based on Lourenço et al. (2015) and Cardozo and Pereyra (2018). To date, in the P. olfersii group, NORs have been observed on chromosomes 3, 4, or 7, but not on chromosome 8 (Milani et al. 2010).
When comparing NOR locations across the P. gracilis group, some interesting patterns emerge. As in P. evangelistai, chromosome 8 carries a NOR in all the remaining species of the P. gracilis group, except P. barrioi, in which the NOR-bearing chromosome is chromosome 10 (Provete et al. 2012; Ferro et al. 2022, Fig. 5). In the karyotype description of P. barrioi, the first species of this group to have its chromosomes studied, the authors based the numerical classification of the chromosomes on their relative sizes. However, Ferro and collaborators, when describing the karyotypes of P. gracilis, P. lisei, and P. carrizorum, chose to designate the small metacentric chromosome bearing NOR in these species as chromosome 8, prioritizing the establishment of chromosome homology based on the presence of the NOR rather than relative size. This hypothesis, however, still demands validation, as does the possibility that the NOR-bearing chromosome of P. barrioi is also homologous to chromosome 8 of the other species in the group. Given that chromosomes 8–11 in each of these karyotypes are very similar in size and morphology, this analysis of homology across the species has been challenging, requiring additional chromosome markers. The presence of an interstitial C-band on the short arm of the NOR-bearing chromosomes 8 in P. evangelistai and P. carrizorum from Palmas-PR, and chromosome 10 of P. barrioi, as detected in this study, may support the homology of these chromosomes (see Fig. 5). Considering it is a small band, hardly revealed in some C-banded metaphases, we cannot discard the possibility that it is also present on chromosome 8 of P. lisei and P. gracilis, which comprise the sister clade of the remaining four species of the P. gracilis group, including P. evangelistai, P. carrizorum, and P. barrioi (see Fig. 5).
With respect to the intrachromosomal location, the NOR on chromosome 8 of P. evangelistai resembles that of P. lisei and P. carrizorum from Misiones, as it is interstitial on the long arm, although in P. carrizorum from Misiones the interstitial NOR extends up to the pericentromeric region. In a sharper contrast, the NOR is distal in P. gracilis (Ferro et al. 2022), P. barrioi (Provete et al. 2012), and in P. carrizorum from Palmas-PR. Assuming the hypothetical homology for these NOR-bearing chromosomes mentioned above, the observed variation could be attributed to intrachromosomal rearrangements, such as inversions and transposition. However, additional chromosomal markers are still required to further assess the extent of similarity among these NOR-bearing chromosomes and to accurately identify the mechanisms underlying the observed changes. Moreover, based on the phylogenetic relationships currently recognized for these species (see Fig. 5), it is not possible to confidently establish the apomorphic condition(s) of NOR position (i.e., interstitial, pericentromeric, or terminal), nor to infer the evolutionary direction of any putative chromosomal changes.
In addition to the difference in NOR-bearing chromosomes, other notable interpopulational variation was noted in P. carrizorum. The karyotype of specimens from Misiones also differs from that of individuals from Palmas-PR in the distribution of PcP190 satDNA chromosomal clusters. Taken together, these findings highlight the need for further analyses to properly assess whether these cytogenetic differences represent interspecific or intraspecific variation (see further discussion in the next section).
Another interesting feature in the karyotype of P. carrizorum from Palmas-PR is the terminal C-band on the short arm of chromosome 3, which varied notably in size, even between homologues of the same individual. Considering that the smaller band was barely detectable in our analysis, we cannot discard the possibility that it also exists in P. carrizorum from Misiones, although it was not observed in the sample analyzed by Ferro et al. (2022). Polymorphisms involving terminal C-bands are not uncommon and have been reported in other Physalaemus species, such as P. centralis (Vittorazzi et al. 2014b), P. cuvieri (Silva et al. 1999), and even P. lisei, a species of the P. gracilis group, which showed polymorphic terminal C-bands on chromosome 1 (Ferro et al. 2022).
When comparing the distribution of heterochromatin within the P. gracilis group, one notable feature is the reduced amount of constitutive heterochromatin present in all karyotyped species, which is mostly restricted to the centromeric regions (Provete et al. 2012; Ferro et al. 2022, present work). In addition, all the four previously karyotyped species show a remarkable C-band in the pericentromeric region of chromosome 3 (Provete et al. 2012; Ferro et al. 2022), a feature that was not observed in P. evangelistai. Nonetheless, the sequence composition of this heterochromatic band likely differs among the species that possess it, as suggested by the chromosome mapping of PcP190 satDNA. The pericentromeric band on chromosome 3 is enriched with this satDNA in P. carrizorum and P. barrioi, but not in P. gracilis and P. lisei (Ferro et al. 2022; present study, further discussion in the next section).
Finally, other remarkable non-centromeric C-bands detected in the karyotype of P. evangelistai were observed on chromosomes 8 and 9, colocalized with the NORs. The association between NOR and heterochromatin is common in anurans and have been reported for species from different families (examples in bufonid and hylid species in Schmid [1978], and in leptodactylid species in Vittorazzi et al. [2016]). Among the remaining karyotyped species of the P. gracilis group, only P. gracilis also exhibited NOR-associated heterochromatin (Ferro et al. 2022). The molecular composition of these regions, however, remain to be elucidated.
PcP190 satellite DNA in the Physalaemus
gracilis group
Heterochromatic blocks can serve as valuable markers for the identification of chromosome pairs and may even aid in inferring homologies between closely related species, as well as in distinguishing specific features of populations or species (e.g., Carvalho et al. 2005; Targueta et al. 2010; Guzmán-Markevich et al. 2022). The interstitial C-band on the short arm of the principal NOR-bearing chromosomes of the P. gracilis group, discussed in the previous section, is one such example. However, this type of data should be analyzed with caution, as heterochromatin includes satDNAs or other repetitive sequences characterized by accelerated evolutionary rates (Ugarković and Plohl 2002; Mehrotra and Goyal 2014; Garrido-Ramos 2017). In this context, the characterization of satDNA can be helpful.
The PcP190 satDNA has been mapped to the karyotypes of several Hyloidea species, contributing significantly to comparative cytogenetics within related clades (Vittorazzi 2014a, 2016; Gatto et al. 2018; Bueno et al. 2021). In our study, by analyzing specimens of P. carrizorum from Palmas-PR (Brazil), we found a chromosomal distribution of PcP190 satDNA clusters that differs from that previously reported for individuals from Misiones, Argentina (approximately 242 km from Palmas-PR). While the specimens from Misiones showed PcP190 satDNA clustered on chromosome 3, in specimens from Palmas this satDNA is clustered on chromosomes 1 and 3. Interpopulational variation in the number of conspicuous chromosomal satDNA clusters was previously reported by Vittorazzi et al. (2014a) for a genetic lineage of P. cuvieri usually referred to as lineage 2 (Lourenço et al. 2015; Nascimento et al. 2019). In specimens of lineage 2 from two localities (i.e., Uberlândia-MG and Três Lagoas-MS), PcP190 clusters were present in the (peri)centromeric region of all chromosomes except for chromosome 8, whereas specimens from Passo Fundo displayed strong signals of a PcP190 probe on chromosomes 1–5 and weak signals on some small chromosomes (Vittorazzi et al. 2014a). On the other hand, the number of PcP190 satDNA chromosomal clusters revealed by FISH in P. ephippifer is remarkably different from those observed in all genetic lineages recognized for P. cuvieri (Vittorazzi et al. 2014a).
In the case of P. carrizorum from Misiones and Palmas, in addition to the difference in the number of PcP190 chromosomal clusters, specimens from these localities also differ in the intrachromosomal position of the NOR and the distribution of heterochromatic blocks (as discussed above). Therefore, further taxonomic analyses are still needed to evaluate whether the cytogenetic differences reported here are due to intraspecific variation or reflect cryptic species. A PcP190 cluster on chromosome 1, similar to that observed in P. carrizorum from Palmas, was also found in P. evangelistai and P. barrioi, species closely related to P. carrizorum, whereas it was not found in P. lisei and P. gracilis (Ferro et al. 2022), which composed another clade in the P. gracilis group (see Fig. 5). Based on these findings, it is likely that the PcP190 cluster on chromosome 1 was lost in the P. carrizorum population from Misiones. However, whether the differential cytogenetic features observed in specimens from Misiones and Palmas play any role in the evolutionary divergence of these frogs remains to be elucidated.
Chromosome mapping of the U2 snRNA gene in the P.
gracilis group
Finally, the mapping of U2 snRNA gene also allowed a better characterization of the karyotypes in the P. gracilis group. Sites carrying U2 snRNA gene have already been mapped at the chromosomal level in other leptodactylid species, including species of Physalaemus (Souza et al. 2025) and Leptodactylus (Gazoni et al. 2021). In the P. gracilis group, U2 snRNA gene clusters are restricted to the terminal region of the short arm of chromosome 6 in the four analyzed species. Moreover, this condition (i.e., the presence of U2 snRNA gene cluster on chromosome 6) is also observed in other Physalaemus and Leptodactylus species (Gazoni et al. 2021; Souza et al. 2025), supporting the hypothesis proposed by Souza et al. (2025) of homology between chromosome 6 among these species. However, mapping this sequence in other genera of the family is still required to draw more robust conclusions regarding the potential of U2 snRNA gene cluster as a reliable marker for identifying the homology of chromosome 6 in Leptodactylidae.
Conclusion
The presence of a constitutive pericentromeric heterochromatin band on the short arm of the NOR-bearing chromosomes may reinforce the hypothesis of homology between chromosome 10 of P. barrioi and chromosome 8 of the other species of the P. gracilis group. In contrast to the conserved location of a cluster of the U2 snRNA gene on chromosome 6, the NOR and PcP190 markers varied across the lineages within this group, even between two different populations of P. carrizorum. These findings highlight the need for further analysis of the diversity harbored by this species.
Author contributions
PHPM: Conceptualization, Investigation, Methodology, Validation, Visualization, Writing - original draft, review and editing. LHBS: Conceptualization, Methodology, Writing - review and editing. JMF: Supervision, Writing - review and editing. LBL: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing - review and editing. All authors approved the final version of the manuscript.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barrio A (1965) El género Physalaemus (Anura, Leptodactylidae) en la Argentina.Physis 25(70): 421–448.
- 2Barth A SoléM Costa MA (2009) Chromosome polymorphism in Phyllomedusa rohdei populations (Anura: Hylidae).Journal of Herpetology 43: 676–679. 10.1670/08-210.1 · doi ↗
- 3Bueno GP Gatto KP Gazolla CB Leivas PT Struett MM Moura M Bruschi DP (2021) Cytogenetic characterization and mapping of the repetitive DN As in Cycloramphus bolitoglossus (Werner, 1897): More clues for the chromosome evolution in the genus Cycloramphus (Anura, Cycloramphidae). P Lo S ONE 16(1): e 0245128. 10.1371/journal.pone.0245128 PMC 780616433439901 · doi ↗ · pubmed ↗
- 4Brum-Zorrilla N Saez FA (1968) Chromosomes of Leptodactylidae (Amphibia Anura).Experientia 24(9): 969. 10.1007/BF 021386895709058 · doi ↗ · pubmed ↗
- 5Cardozo DE Pereyra MO (2018) A new species of Physalaemus (Anura, Leptodactylidae) from the Atlantic Forest of Misiones, northeastern Argentina.Zootaxa 4387(3): 580–590. 10.11646/zootaxa.4387.3.1029690482 · doi ↗ · pubmed ↗
- 6Carvalho R Soares Filho WS Brasileiro-Vidal AC Guerra M (2005) The relationships among lemons, limes and citron: A chromosomal comparison.Cytogenetic and Genome Research 109(1–3): 276–282. 10.1159/00008241015753587 · doi ↗ · pubmed ↗
- 7Cassini CS Cruz CAG Caramaschi U (2010) Taxonomic review of Physalaemus olfersii (Lichtenstein & Martens, 1856) with revalidation of Physalaemus lateristriga (Steindachner, 1864) and description of two new species.Zootaxa 2491(1): 1–33. 10.11646/zootaxa.2491.1.1 · doi ↗
- 8De Lucca E Jim J Foresti F (1974) Chromosomal studies in twelve species of Leptodactylidae and one Brachycephalidae.Caryologia 27(2): 183–192. 10.1080/00087114.1974.10796573 · doi ↗