Clematis danxiacola (Ranunculaceae), a new species from the Danxia landform area in Zhejiang Province, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
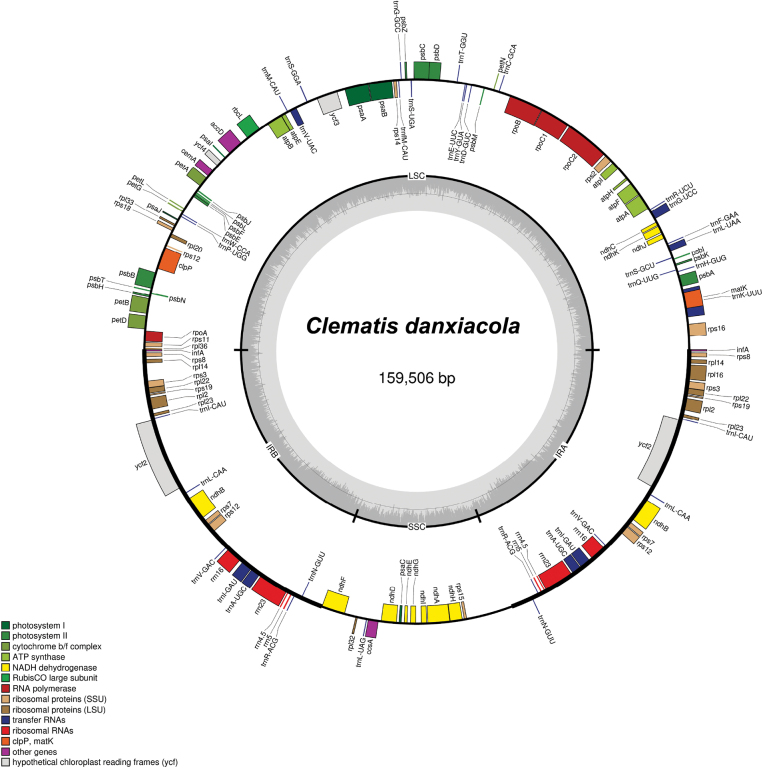 Figure 1
Figure 1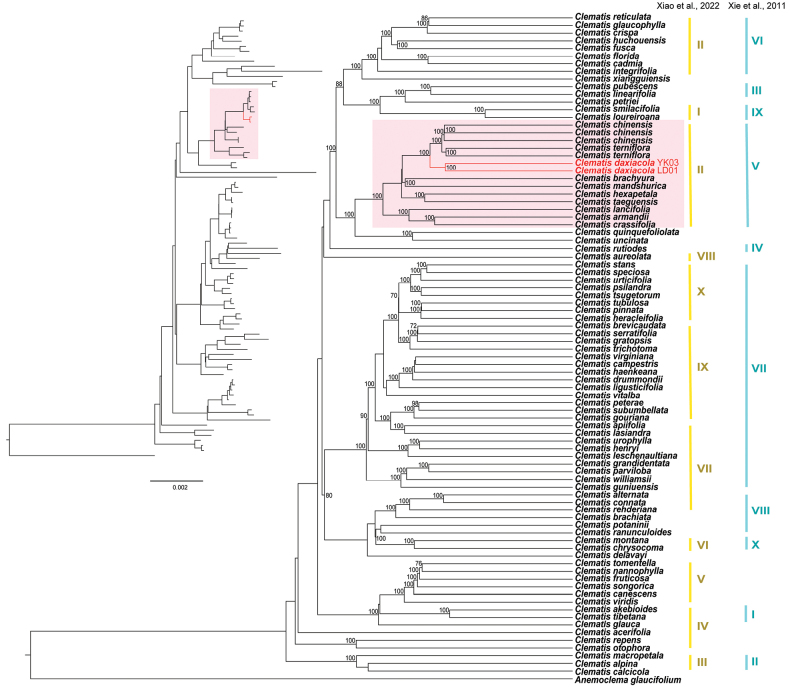 Figure 2
Figure 2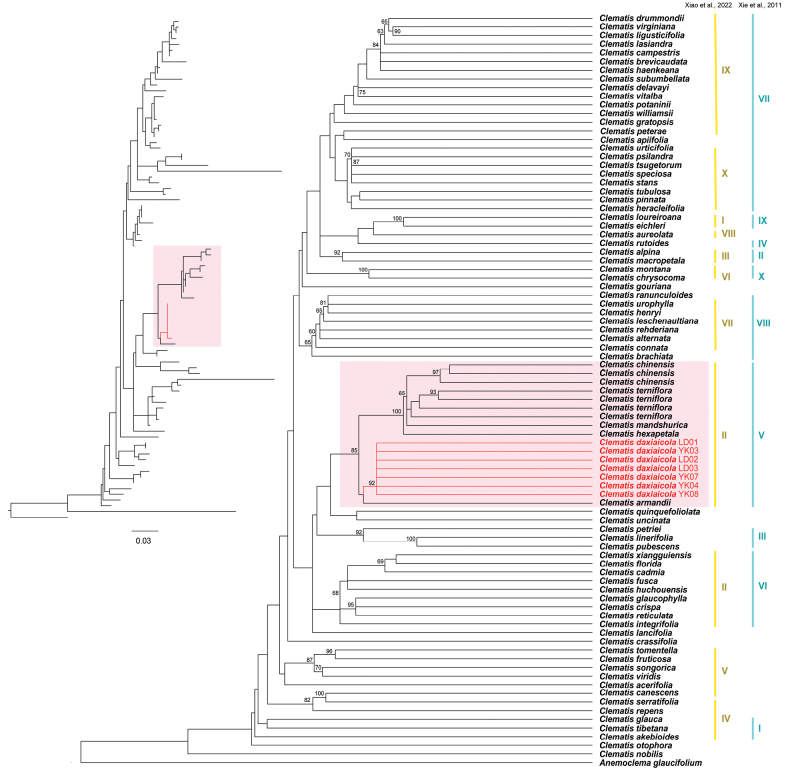 Figure 3
Figure 3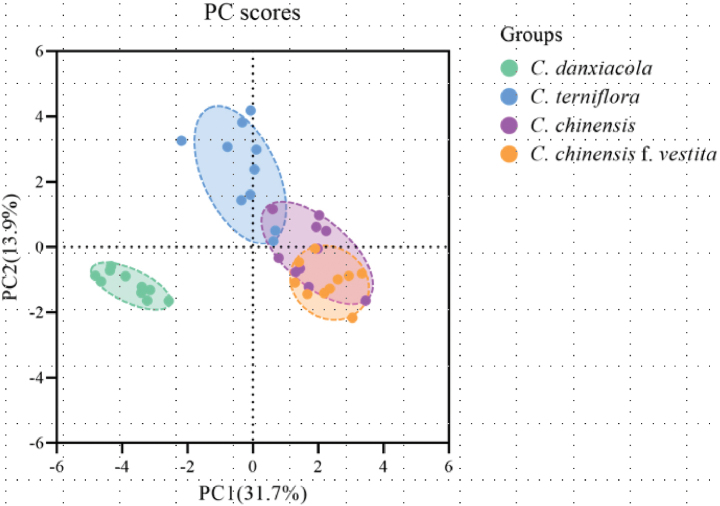 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6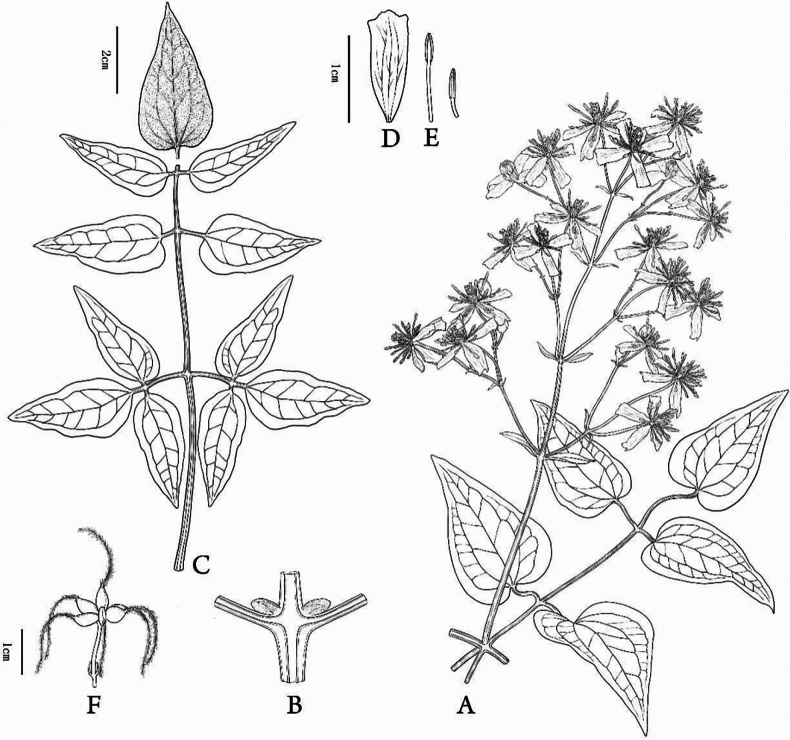 Figure 7
Figure 7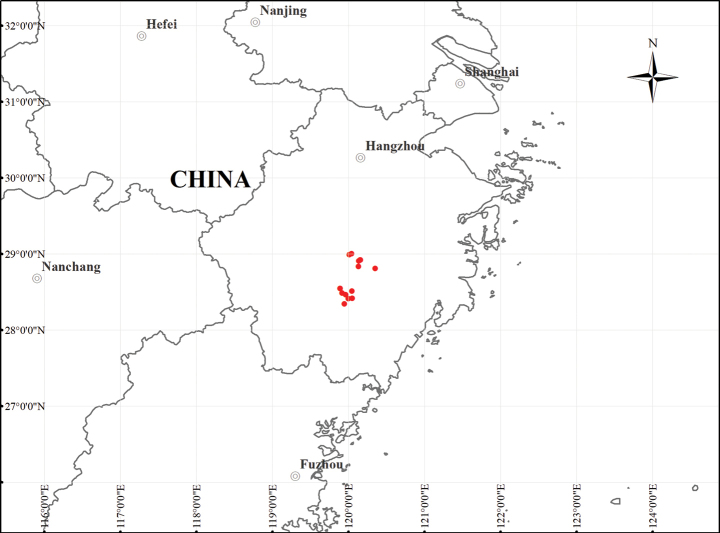 Figure 8
Figure 8| Molecular specimen ID | Herbarium specimen ID | Locality | Coordinates | Altitude (a.s.l.) |
|---|---|---|---|---|
| YK-03 | ZMNH0067437 | Feilongshan, Xicheng Subdistrict, Yongkang County, Jinhua City |
| 181 |
| YK-04 | ZMNH0067438 | Longqingkeng, Wangxitian Village, Xiangzhu Town, Yongkang County, Jinhua City |
| 246 |
| YK-07 | ZMNH0067439 | Qianlang Village, Shizhu Town, Yongkang County, Jinhua City |
| 175 |
| YK-08 | ZMNH0067436 PE02621603 KUN1644125 | Chiyantang Reservoir, Xianling Village, Zhiying Town, Yongkang County, Jinhua City |
| 172 |
| LD-01 | ZMNH0067440 | Nanbenjis, Baiyunshan, Baiyun Subdistrict, Liandu District, Lishui City |
| 176 |
| LD-02 | ZMNH0067441 | Huangnidan Village, Zijin Subdistrict, Liandu District, Lishui City |
| 98 |
| LD-03 | ZMNH0067442 | Huangcun Village, Huangcun Town, Liandu District, Lishui City |
| 129 |
| Variable | Data Type | PC1 | PC2 | PC3 |
|---|---|---|---|---|
| Specimen color after drying (blackened 1 / unblackened 0) | DV | 0.808 | −0.366 | 0.121 |
| Compound leaf length (cm) | CV | −0.46 | 0.185 | 0.564 |
| Compound leaf width (cm) | CV | −0.605 | 0.162 | 0.539 |
| Compound leaf length/width ratio | CV | 0.42 | −0.093 | −0.105 |
| Indumentum on leaf abaxial surface (densely pubescent 1 / glabrous or glabrescent 0) | DV | −0.643 | −0.612 | −0.187 |
| Terminal leaflet length (cm) | CV | −0.693 | −0.249 | 0.47 |
| Terminal leaflet width (cm) | CV | −0.215 | −0.064 | 0.367 |
| Terminal leaflet length/width ratio | CV | −0.548 | −0.256 | 0.303 |
| Petiolule length (cm) | CV | −0.024 | 0.563 | 0.361 |
| Pedicel length (cm) | CV | 0.158 | 0.128 | 0.256 |
| Calyx apex morphology (truncate 1 / mucronate 0) | DV | −0.879 | −0.399 | −0.048 |
| Calyx length (mm) | CV | −0.4 | 0.596 | 0.254 |
| Calyx width (mm) | CV | −0.764 | 0.194 | −0.3 |
| Calyx length/width ratio | CV | 0.403 | 0.346 | 0.56 |
| Stamen number | CV | 0.379 | 0.262 | −0.048 |
| Fruit length (mm) | CV | −0.289 | 0.609 | −0.312 |
| Persistent style length (cm) | CV | 0.502 | −0.279 | 0.386 |
| Fruit length/persistent style ratio | CV | −0.296 | 0.512 | −0.508 |
| Taste of fresh leaves (pungent 1 / non-pungent 0) | DV | −0.808 | 0.366 | −0.121 |
| Habitat (ridge, hillside 1 / valley, plain or wetland 0) | DV | −0.879 | −0.399 | −0.048 |
| Characters |
|
|
| |
|---|---|---|---|---|
| Plant on drying | not turning black | not turning black | turning bright black | turning black |
| Branchlet | densely persistently appressed-puberulous; nodes with base of petiole and petiolules usually dark purple | puberulous; nodes with petiole and petiolules usually green | glabrous or sparsely puberulous; nodes with petiole and petiolules usually green | ± densely puberulous |
| Leaf | 1-pinnate, 5-foliolate; 1- or 2-pinnate on new shoots in summer-autumn | 1-pinnate, 5 (–7)-foliolate | 1-pinnate, 5-foliolate | 1-pinnate, 5-foliolate |
| leaflet blade | thick-papery or subleathery, 3.5–8.0 (–9.5) × 2.0–5.0 (–6.2) cm, abaxially densely persistently appressed-puberulous, | papery to subleathery, 2.5–8 × 1–4.2 cm, both surfaces sparsely puberulous, glabrescent | papery, 1.5–9.5 × 0.7–6.4 cm, both surfaces subglabrous or very sparsely puberulous only on basal veins | 3.5 (–5) × 2 (–2.5) cm, abaxially ± densely puberulous |
| Bract | narrowly lanceolate to linear, 0.7–1.5 (–2.3) cm long, petiolate, rarely leaflike | linear, elliptic or oblong, 0.8–3.5 (–5.0) cm long | petiolate and elliptic to oblong or sessile, small, and linear | petiolate and elliptic to oblong or sessile, small, and linear |
| Pedicel | 1–2 cm long, densely persistently appressed-puberulous | 0.5–3.0 cm long, puberulous | 1.4–3.0 cm long, sparsely puberulous | 1.4–3.0 cm long, sparsely puberulous |
| Sepal | cuneate or oblanceolate, 10–14 × 4–6 mm, apex subtruncate and premorse | obovate-oblong to oblong, 5–15 × 2–6 mm, apex acute to obtuse and entire | obovate-oblong, oblanceolate, or lanceolate, 6–13 × 1.8–3 (–4) mm, apex acute and entire | obovate-oblong, oblanceolate, 6–9 × 2–3 mm, apex acute and entire |
| Pistil | (5–) 8–12 | 4–7 | — | — |
| Achene | ovate, 5.4–6.3 × 3.0–3.7 mm, compressed and slightly swollen on middle both sides, margin not or slightly thickened | broadly elliptic to obovate, 6–9 × 3–6 mm, strongly compressed, margin distinctly thickened | elliptic, 5–7 × 3.5–4 mm, appressed puberulous | — |
| Habitat | Danxia landform, low hills, purple sandstone and conglomerate, arid habitats, alt. 50–550 m a.s.l. | various landforms, humid habitats, alt. below 600 m a.s.l. | various landforms, humid habitats, alt. 100–1500 m a.s.l. | thickets, alt. 300–1100 m a.s.l. |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Biological and pharmacological studies of plants · Plant Diversity and Evolution
Introduction
Clematis L. (1753: 543) is one of the few cosmopolitan genera in the buttercup family (Ranunculaceae), comprising approximately 300 species predominantly distributed across north temperate zones (Wang 1980; Ziman and Keener 1989; Tamura 1995; Johnson 1997; Grey-Wilson 2000; He et al. 2021). China hosts approximately 147 Clematis species, with 93 being endemic (Wang and Bartholomew 2001).
The taxonomy of Clematis has been notoriously difficult due to high species-level morphological variation (Johnson 1997; Brandenburg 2000; Grey-Wilson 2000; Wang and Li 2005). Multiple taxonomic schemes have emerged in recent years addressing genus delineation, infrageneric classification, and species delimitation in Clematis (Tamura 1995; Johnson 1997; Wang 1998, 2003, Grey-Wilson 2000; Wang and Li 2005; Xiao et al. 2022).
Recent molecular phylogenetic studies suggest substantial convergence in floral and trophic characteristics in Clematis, challenging previous infrageneric classifications based on morphological characteristics (Miikeda et al. 2006; Xie et al. 2011), but conforms to Johnson’s broad concept that Clematis should encompass all previously recognized genera of subtribe Clematidinae (Johnson, 1997). Currently, ten clades within Clematis have been resolved as regional geographic groups (Xie et al. 2011).
During a field survey of the Danxia landform area in Zhejiang, China, we identified an unknown species of Clematis. The species displayed morphological similarities to C. terniflora DC. (1818: 137), but was notably distinct in habitat, and leaf and floral characteristics, prompting investigation of its taxonomic status.
Materials and methods
Morphological observation
From 2021 to 2024, we conducted field excursions to the Danxia landform area in Zhejiang Province, China, including Yongkang County, Wuyi County, and Wucheng District (Jinhua); Songyang County, Jinyun County, Qingtian County, and Liandu District (Lishui); Jiangshan County and Kecheng District (Quzhou); Zhuji County and Xinchang County (Shaoxin); and Yuyao County and Fenghau District (Ningbo), to observe and investigate this unknown species. Specimens in the principal herbaria in Zhejiang Province (HHBG, HTC, HZU, and ZM) (Thiers 2024) and specimen photographs from the Chinese Virtual Herbarium (CVH; www.cvh.ac.cn/) were examined to identify specimens similar to the unknown species. High-definition images of similar specimens from international herbaria (A, BM, E, FI, G, K, LD, LINN, MPU, and P) were also consulted. Based on field investigation and herbarium specimen evaluation, the morphology of this new species was documented through comparison with protologues from Greuter (1965) and de Candolle (1818).
Sampling and sequencing
Material sampling
From August 10 to August 18, 2023, we collected one leaf sample each from 7 populations of the unknown taxon in Jinhua and Lishui cities of the Danxia landform area in Zhejiang Province (Table 1). Leaves were initially placed in non-woven bags, then stored in sealing bags containing allochroic silica gel for rapid drying. Herbarium specimens were preserved in the herbaria of Zhejiang Museum of Natural History (ZM); Institute of Botany, Chinese Academy of Sciences (PE); and Kunming Institute of Botany, Chinese Academy of Sciences (KUN) (Table 1).
Table 1.: List of analyzed samples of Clematis danxiacola sp. nov.
DNA extraction, PCR amplification, and sequencing
Whole-genomic DNA from silica-dried leaf tissue was extracted using a modified CTAB protocol (Chen et al. 2014). For plastome sequencing, high-quality DNA was fragmented into lengths of ≤800 bp. Fragment quality was verified using an Agilent Bioanalyzer 2100 (Agilent Technologies, Palo Alto, CA, USA). A short-insert (500 bp) paired-end library was constructed and sequenced by Nanopore Technology (Wuhan, China) on a DNBSEQ-T7 sequencer with 150 bp paired-end reads.
For amplification and sequencing of ribosomal DNA (rDNA) internal transcribed spacer (ITS) regions, primers were designed using ITS sequences of closely related species: forward, 5′-ATGCGATACTTGGTGTGAAT-3′; reverse, 5′-GACGCTTCTCCAGACTACA-3′. The amplification fragments encompass the ITS1, ITS2, 5.8 rDNA, and 26S rDNA regions. DNA amplification was conducted in 50 μL reactions containing 1 μL of total DNA (20 ng/μL), 5 μL of 10× buffer with 2.5 mM Mg^2+^, 1 μL of dNTP (10 mM), 1.5 μL of each primer (10 μM), and 1 μL of Taq polymerase (5 u/μL) in an ABI-2720 thermocycler (Applied Biosystems, Waltham, MA, USA). The PCR protocol consisted of initial denaturation at 95 °C for 5 min, followed by 35 cycles of denaturation at 95 °C for 30 s, annealing at 58 °C for 30 s, and extension at 72 °C for 90 s, with a final extension at 72 °C for 7 min. PCR products were purified by ethanol precipitation. The purified products were analyzed by electrophoresis on a 1% (w/v) agarose gel and spectrophotometric analysis (NanoDrop 2000; Thermo Fisher Scientific, Waltham, MA, USA) before Sanger sequencing using an ABI3730-XL sequencer (Applied Biosystems).
Data assembly and annotation
For plastome assembly and annotation, raw sequencing data was filtered using FastQC (www.bioinformatics.babraham.ac.uk/projects/fastqc/). Clean paired-end reads were de novo assembled using the GetOrganelle pipeline (Jin et al. 2020) using SPAdes 3.10.1 as the assembler (Bankevich et al. 2012). The plastome of C. terniflora (NC028000) served as the reference. The assembled plastome was imported into Geneious Prime (https://www.geneious.com) for annotation and identification of putative starts, stops, and intron positions through comparison with homologous genes of published Clematis plastomes. The circular plastome map of the new species was generated using OGDRAW (http://ogdraw.mpimp-golm.mpg.de/) (Greiner et al. 2019).
Phylogenetic analyses
For phylogenetic analyses, complete plastome sequences of 85 Clematis species and ITS sequences of 80 Clematis species were obtained from GenBank (Suppl. material 1: table S1). The sampling strategy encompassed all major clades within Clematis previously identified (Xie et al. 2011; Xiao et al. 2022). The plastome and ITS dataset were aligned independently using multiple alignment using fast Fourier transform (MAFFT) v7.490 implemented in Geneious Prime (Katoh and Standley 2013). Phylogenetic analyses based on ITS and plastome sequences (excluding one copy of the inverted repeat) were conducted to explore the evolutionary relationship among the new species and other Clematis species using the maximum likelihood method with RAxML-HPC v8.2.12 (Stamatakis 2014) on the CIPRES Science Gateway (http://www.phylo.org/). According to Jiang et al. (2017) and Xiao et al. (2022), Anemoclema glaucifolium (Franch.) W.T. Wang (Ranunculaceae) was chosen as an outgroup.
Morphological Analysis
Morphological characteristics were documented from 10 wild individuals of C. danxiacola sp. nov. during field surveys. For comparative analysis, 10 herbarium specimens each of C. terniflora, C. chinensis, and C. chinensis var. vestita were examined from the Chinese Virtual Herbarium (CVH; https://www.cvh.ac.cn/).
Twenty quantitative traits (Table 2) were selected to construct a morphological matrix. Principal component analysis (PCA) was performed for dimensionality reduction, and a multidimensional morphospace was established based on the first two principal components. All computations were conducted using GraphPad Prism v10.1.2 (Build 324).
Results
Characteristics of the plastome
The plastome size of Clematis danxiacola sp. nov. is 159,506 bp and comprises two 31,040 bp inverted repeats, a 18,407 bp small single-copy region, and a 82,836 bp large single-copy region. The overall GC content is 38%, and the GC contents of the large single-copy, small single-copy, and inverted repeat regions are 36.3%, 31.4%, and 42.0%, respectively. The complete plastome contains 136 functional genes, including 92 protein-coding genes, 36 tRNA genes, and 8 rRNA genes (Fig. 1, Suppl. material 1: table S2). The complete plastome sequence of C. danxiacola after annotation was deposited in GenBank (PQ246280).
Plastome map of Clematis danxiacola sp. nov. Genes inside the circle are transcribed clockwise, while those outside are transcribed counterclockwise. In the inner circle, light gray represents the AT content, and dark gray represents the GC content. Different colors indicate genes belonging to distinct functional groups.
Molecular analysis
Phylogenetic analyses based on both plastome and ITS datasets revealed that individuals of C. danxiacola were clustered with high bootstrap support values (Figs 2–3). The new species is closely related to C. terniflora and C. chinensisOsbeck (1757: 205) and belongs to clade V (sensu Xie et al. 2011) or clade II (sensu Xiao et al. 2022) of Clematis, which comprises species of sect. Flammula DC., sect. Viticella DC., and sect. Viorna (sensu Tamura 1995).
Maximum likelihood tree inferred from Clematis plastomes to elucidate the phylogenetic position of C. danxiacola sp. nov. Numbers above the branches indicate the bootstrap values (≥50%) of the maximum likelihood analysis. Branches and names in red indicate the new species (C. danxiacola sp. nov). Light red shading highlights the clade containing the new species, as revealed by phylogenetic analyses in Xie et al. (2011) and Xiao et al. (2022). Numerals on the right indicate the ten Clematis clades recognized in Xie et al. (2011) and Xiao et al. (2022): yellow, Xiao et al. (2022); blue, Xie et al. (2011).
Maximum likelihood tree inferred from ITS sequences to elucidate the phylogenetic position of C. danxiacola sp. nov. Numbers above the branches indicate the bootstrap values (≥50%) of the maximum likelihood analysis. Branches and names in red indicate the new species (C. danxiacola sp. nov). Light red shading highlights the clade containing the new species, as revealed by phylogenetic analyses in Xie et al. (2011) and Xiao et al. (2022). The GenBank accession numbers used for the analysis are indicated after the species names. Numerals on the right indicate the ten Clematis clades recognized in Xie et al. (2011) and Xiao et al. (2022): yellow, Xiao et al. (2022); blue, Xie et al. (2011).
Morphological comparison
Morphological comparisons between C. danxiacola sp. nov., C. terniflora, C. chinensis, and C. chinensis var. vestita (Rehder & E.H.Wilson) W.T. Wang (1998: 158) are presented in Table 2. This new species shares characteristics with C. terniflora and C. chinensis in having climbing stems, entire leaflet, cymes, small flowers, white and spreading sepals, and glabrous stamens; however, it is distinctly different from them in habitat, texture and indumentum of leaflet blades, and sepal apex shape (Figs 5–7).
PCA
Multivariate analysis provided a quantitative assessment of morphological differentiation among the four species. The analysis revealed distinct variation patterns along the first two axes (Fig. 4). Two clearly separated clusters emerged: one comprising all specimens of C. danxiacola sp. nov., and another containing its allied species. C. danxiacola demonstrated a discontinuous distribution in the principal component (PC) 1–PC2 morphospace compared with the other three taxa, forming statistically distinct clusters (Fig. 4).
Principal component axes 1 and 2 representing morphological data of C. danxiacola sp. nov. and three other similar species.
PCA revealed that the first seven principal components derived from the 20 morphological traits collectively explained 85.16% of the total variation. PC1 accounted for 31.72% of the variance, while PC2 explained 13.93%.
The primary traits associated with PC1 included specimen color after drying, sepal apex characteristics, habitat preferences, fresh leaf taste, and sepal width. PC2 was predominantly influenced by leaf abaxial surface indumentum, fruit length, sepal length, and petiolule length (Table 2).
Taxonomic treatment
Clematis
danxiacola
Taxon classificationPlantaeRanunculalesRanunculaceae
J.P.Li, P.Li & Z.H.Chen sp. nov.
BE9642EF-85BA-5797-9DF4-9A3E9078CE64
urn:lsid:ipni.org:names:77371642-1
Type.
China • Zhejiang Province, Jinhua City, Yongkang County, Zhiying Town, Xianling Village, Chiyantang Reservoir, purple glutenite in the Danxia landform area, in bushes at the forest margin at foothills, alt. 172 m a.s.l., 28°54'29.28"N, 120°8'7.86"E, August 18, 2023, He-Ping Chen, Jun-Ping Li et Zheng-Hai Chen YK23081807 (holotype: ZM barcode ZMNH0067436! isotypes: HZU!, KUN!, PE!).
Diagnosis.
This new species resembles C. terniflora but differs in several characteristics including leaflet blades abaxially densely persistently appressed-puberulous (vs. sparsely puberulous and glabrescent), sepals cuneate or oblanceolate, apex subtruncate and premorse (vs. obovate–oblong to oblong, apex ± acute to obtuse and entire), pistils (5–) 8–12 (vs. 4–7), achene ovate, compressed but slightly swollen in the middle, margin not or slightly thickened (vs. broadly elliptic to obovate, strongly compressed, margin distinctly thickened).
Morphological description.
Evergreen woody vine. Stem with shallowly 6–12-grooved; branchlets densely gray appressed-puberulous, glabrescent except at nodes; axillary buds triangular–ovate, densely gray puberulous. Leaves opposite, 1-pinnate, usually 5-foliolate, sometimes 2-pinnate on new shoots in summer–fall; petiole 3–7 cm long, base with nodes usually dark purple or gray, puberulous; leaflet blades thick-papery or subleathery, ovate, 3.5–8.0 (–9.5) × 2.0–5.0 (–6.2) cm, apex acuminate, base broadly cuneate, subrounded to shallow cordate, margin entire, slightly revolute when drying, adaxially appressed-puberulous when young, glabrescent except for basal veins, lustrous, basal veins slightly prominent, abaxially densely persistently appressed-puberulous, basal veins prominent when dry; leaflets of summer–fall shoots thick leathery, ovate–lanceolate or lanceolate, 2.5–10.0 × 1–3 cm, apex long acuminate, base cuneate; petiolules 0.7–2.0 cm long, base usually dark purple, sometimes tendrilous. Paniculate cymes axillary or terminal, usually many-flowered, 5–15 (–30) cm long; bracts usually narrowly lanceolate to linear, 0.7–1.5 (–2.3) × 1.5–4.0 mm, petiolate, rarely leaflike, both surfaces appressed-puberulous. Flowers 2–3 cm in diameter, pungent aromatic, bisexual, erect; buds ovoid, apex acute. Pedicel 1–2 cm long, together with peduncle and rachis densely persistently appressed-puberulous. Sepals 4, white, spreading, cuneate or oblanceolate, 10–14 × 4–6 mm, apex subtruncate and premorse, adaxially glabrous, abaxially densely puberulous, margin velutinous. Stamens 26–49, white, glabrous, 6–11 mm long; filaments 3–7 mm long; anthers linear, 2–4 mm long, apex obtuse or minutely apiculate. Pistils (5–) 8–12; ovaries pubescent; style densely villous. Achenes usually 3–7, brown, ovate, compressed but slightly swollen in the middle, 5.4–6.3 × 3.0–3.7 mm, densely ascending pubescent, margin not or slightly thickened; persistent styles 2.5–3.0 cm long, plumose, whitish (Figs 5–7).
Habitat and habitus of C. danxiacola sp. nov. A. Habitat; B. Leaves of spring new shoot; C. Inflorescence of spring new shoot; D. Infructescence of spring new shoot; E. Branchlet and leaves of summer–fall new shoot; F. Inflorescence of summer–fall new shoot. Images captured by Dong-Hao Wu (A), Zheng-Hai Chen (B–E), Jun-Ping Li (F).
Morphological characteristics of C. danxiacola sp. nov. A. Flowering branch (b, summer–fall new shoot); B. Branchlet and axillary bud; C. Leaf (a, spring new shoot; b, summer–fall new shoot); D. Leaflet blade; E. Bud; F. Flower; G. Dissected flower (showing sepals); H. Fruiting branch; I. Infructescence; J. Achene and persistent style. Images captured by Zheng-Hai Chen (A, B, Cb, D–F, H, I) in the field; Jun-Ping Li (Ca, G, J) in the laboratory.
Line drawing of Clematis danxiacola sp. nov. A. Flowering branch; B. Stem; C. Leaf; D. Sepal; E. Stamen; F. Infructescence. Drawn by Yi-Chen Wang.
Phenology.
Flowering occurs from May to November, with fruiting from September to January of the following year.
Etymology.
The specific epithet refers to the Danxia landform where the species occurs.
Distribution and ecology.
C. danxiacola occurs in Yongkang County of Jinhua City, and Qingtian County, Jinyun County, and Liandu District of Lishui City, Zhejiang Province, China (Fig. 8). The species inhabits sunny slopes or ridges of low hills of the Danxia landform within the elevation range 50–550 m a.s.l, typically climbing on tree canopies or shrubs at forest edges or roadsides. The substrate consists of soil derived from weathered purple sandy conglomerate.
Geographic distribution of Clematis danxiacola in Zhejiang Province (red dots).
Conservation assessment.
The currently known 19 populations of C. danxiacola are distributed across two prefecture-level cities, four counties, and 14 towns. The total distribution area is approximately 100 km^2^ with an estimated population exceeding 1000 mature individuals. Despite occurring in low-hill areas characterized by frequent human activities and habitat fragmentation, the species demonstrates resilience due to its drought tolerance, arid soil adaptability, climbing capabilities, high reproductive success, and extensive potential habitat availability. Furthermore, the species currently faces minimal anthropogenic pressure as its economic value remains undetermined, resulting in limited local exploitation. Based on these factors, we propose that C. danxiacola be classified as a “Least Concern (LC)” species according to IUCN Red List criteria (IUCN 2024).
Additional specimens examined (paratypes).
China • Zhejiang Province, Jinhua City, Yongkang County, Xicheng Subdistrict, Feilongshan, purple glutenite in the Danxia landform area, the forest margin on slope, alt. 182 m a.s.l., June 29, 2023, Jun-Ping Li, Jian-Ping Zhong et Zheng-Hai Chen YK23062907 (ZM); • same locality, October 9, 2023, Jun-Ping Li, Shang-Jiao Ying et Zheng-Hai Chen YK23100904 (ZM); • same locality, alt. 181 m a.s.l., 28°59'23.88"N, 120°0'41.40"E, August 18, 2023, Zheng-Hai Chen, Jun-Ping Li et He-Ping Chen YK23081801 (ZM, HZU); • Xiangzhu Town, Wangxitian Village, Longqingkeng, purple glutenite in the Danxia landform area, on open land at the forest margin on ridge, alt. 246 m a.s.l., 29°0'8.40"N, 120°2'33.06"E, August 18, 2023, Shang-Jiao Ying et Xia-Shuo Lei YK23081802 (ZM, HZU); • same locality and date, Jun-Ping Li, He-Ping Chen et Zheng-Hai Chen YK23081808 (ZM); • Zhiying Town, Xianling Village, Chiyantang Reservoir, purple glutenite in the Danxia landform area, on bushes at the forest margin on slope, alt. 199 m a.s.l., July 13, 2023, Jun-Ping Li YK23071302 (ZM); • same locality, alt. 177 m a.s.l., October 8, 2023, Jun-Ping Li et Zheng-Hai Chen YK23100801 (ZM); • same locality and date, on ridge bushes, alt. 256 m a.s.l., October 8, 2023, Jun-Ping Li et Zheng-Hai Chen YK23100802 (ZM); • same locality and date, on ridge bushes, alt. 270 m a.s.l., October 8, 2023, Jun-Ping Li et Zheng-Hai Chen YK23100803 (ZM); • same locality and date, on bushes near the hilltop, alt. 277 m a.s.l., October 8, 2023, Jun-Ping Li et Zheng-Hai Chen YK23100804 (ZM); • same locality and date, on ridge bushes, alt. 271 m a.s.l., October 8, 2023, Jun-Ping Li et Zheng-Hai Chen YK23100805 (ZM); • Qianshanyangxia Village, purple glutenite in the Danxia landform area, on ridge bushes, alt. 236 m a.s.l., 28°55'15.13"N, 120°9'9.49"E, November 8, 2023, Zheng-Hai Chen, Jun-Ping Li et Wen-Yuan Xie YK23110802 (ZM); • same locality and date, alt. 268 m a.s.l., 28°55'10.71"N, 120°9'32.15"E, Zheng-Hai Chen, Jun-Ping Li et Wen-Yuan Xie YK23110808 (ZM); • Shizhu Town, Qianlang Village, purple glutenite in the Danxia landform area, on bushes at the forest margin on slope, alt. 175 m a.s.l., 28°49'59"N, 120°7'14.50"E, August 18, 2023, Jun-Ping Li, He-Ping Chen et Zheng-Hai Chen YK23081805 (ZM); • same locality and date, on bushes at the forest margin on ridge, alt. 230 m a.s.l., He-Ping Chen, Jun-Ping Li et Zheng-Hai Chen YK23081806 (ZM); • same locality, November 7, 2023, Zheng-Hai Chen, Jun-Ping Li et Wen-Yuan Xie YK23110702 (ZM). • Lishui City, Liandu District, Nanmingshan Subdistrict, Jinshanxia Village, Duimianshan, purple glutenite in the Danxia landform area, in bushes beside the highway, alt. 160 m a.s.l., July 23, 2023, Dong-Hao Wu LD23072301, LD23072302, LD23072303 (ZM); • Baiyun Subdistrict, Baiyunshan, Nanbenjishan, purple glutenite in the Danxia landform area, in bushes by a trail, alt. 176 m a.s.l., 28°28'54.4"N, 119°55'4.71"E, July 29, 2023, Dong-Hao Wu LD23072901 (ZM); • same locality, August 10, 2023, Wen-Yuan Xie, Jun-Feng Wang, Zheng-Hai Chen et al. LD23081001 (ZM, HZU); • Beibenjishan, purple glutenite in the Danxia landform area, in roadside bushes at the mountain foot, alt. 155 m a.s.l., July 29, 2023, Dong-Hao Wu LD23072902 (ZM); • Zijin Subdistrict, Hebian Village, Bogushan, purple glutenite in the Danxia landform area, in roadside bushes at the mountain foot, alt. 90 m a.s.l., August 5, 2023, Jian-Ping Zhong et Dong-Hao Wu LD23080501 (ZM); • Huangnidan Village, purple glutenite in the Danxia landform area, on bushes amid steep slopes near the highway, alt. 98 m a.s.l., 28°27'37.23"N, 119°57'47.73"E, August 5, 2023, Jian-Ping Zhong et Dong-Hao Wu LD23080502 (ZM); • the same locality, August 10, 2023, Wen-Yuan Xie, Jun-Feng Wang, Zheng-Hai Chen et al. LD23081003 (ZM, HZU); • the same locality, October 10, 2023, Jun-Feng Wang et Zheng-Hai Chen LD23101001 (ZM); • Shuilinggen Village, purple glutenite in the Danxia landform area, on bushes near the highway, alt. 333 m a.s.l., August 5, 2023, Jian-Ping Zhong et Dong-Hao LD23080503 (ZM); • Fuling Subdistrict, Jinshan, purple glutenite in the Danxia landform area, beside a hiking trail on the hillside, alt. 154 m a.s.l., October 10, 2023, Jun-Feng Wang et Zheng-Hai Chen LD23101002 (ZM); • the same locality and date, on tree crown by the hillside road, alt. 160 m a.s.l., Jun-Feng Wang et Zheng-Hai Chen LD23101003 (ZM); • the same locality and date, on tree crown by the hillside road, alt. 165 m a.s.l., Jun-Feng Wang et Zheng-Hai Chen LD23101005 (ZM); • the same locality and date, on bushes by the hillside road, alt. 163 m a.s.l., Jun-Feng Wang et Zheng-Hai Chen LD23101006 (ZM); • the same locality and date, on bushes by the hillside path, alt. 162 m a.s.l., Jun-Feng Wang et Zheng-Hai Chen LD23101007 (ZM); • Huangcun Town, Huangcun Village, purple glutenite in the Danxia landform area, in bushes by the highroad near a small temple, alt. 129 m a.s.l., 28°30'40.02"N, 120°2'48.74"E, July 30, 2023, Dong-Hao Wu LD23073001 (ZM); • Yanquan Subdistrict, introduced from Xiayinkeng Village and cultivated by Yong-Jun Chen, alt. 50 m a.s.l., July 30, 2023, Dong-Hao Wu LD23073002 (ZM); • the same locality, Xiayinkeng Village, Changkeng, purple glutenite in the Danxia landform area, in bushes by the roadside, alt. 235 m a.s.l., August 4, 2023, Dong-Hao Wu et Yong-Jun Chen LD23080401 (ZM); • Taiping Town, on steep shrubs, 119°53'19.8"E, 28°32'34.0"N, alt. 453 m a.s.l., July 27, 2024, Lian-Hai Wu et Zheng-Hai Chen LD24072701 (ZM); • the same locality and date, 119°53'19.87"E, 28°32'34.09"N, alt. 410 m a.s.l., Jun-Feng Wang et Dong-Hao Wu 24072702 (ZM). • Jinyun County, Huzheng Town, on steep shrubs, 120°21'12.12"E, 28°48'18.63"N, alt. 550 m a.s.l., July 7, 2024, Shang-Jiao Ying, Jun-Ping Li, Liang-Dong> et Zheng-Hai Chen JY24070701 (ZM). • Qingtian County, Lakou Town, near Kengkou Village, on shrubs and on rock walls next to the highway, 28°20'31.55"N, 119°56'53.19"E, alt. 53 m a.s.l., August 17, 2024, Jun-Feng Wang, Dong-Hao Wu et Jian-Ping Zhong QT24081701 (KUN, PE, ZM); • the same locality and date, near Yaojun Village, 28°24'31.26"N, 119°59'57.36"E, alt. 102 m a.s.l., Dong-Hao Wu et Jian-Ping Zhong QT24081702 (ZM); • Shuqiao Town, near Shakeng Village, on a tree crown and on a rock wall by the roadside, 28°24'54.20"N, 120°2'51.26"E, alt. 277 m a.s.l., Jun-Feng Wang et Jian-Ping Zhong QT24081703 (ZM).
Discussion
The new species C. danxiacola exhibits close phylogenetic and morphological relationships to C. terniflora and C. chinensis. In the reconstructed phylogenetic tree, C. danxiacola is positioned within sect. Clematis (Wang and Li 2005), which is characterized by opposite phyllotaxy, spreading calyx, and glabrous filaments. This study corroborates previous phylogenetic analyses of Clematis that grouped C. terniflora, C. chinensis, C. armandii Franch. (1885: 184), C. crassifolia Benth. (1861: 7), C. hexapetala Pall. (1776: 735), and C. brachyura Maxim. (1877: 221) as a monophyletic group (Xie et al. 2011; Xiao et al. 2022). The phylogenetic placement of C. danxiacola within the clade demonstrates slight variations between plastome and ITS sequences. In the plastome phylogenetic tree, C. danxiacola specimens form a clade sister to that containing C. terniflora and C. chinensis, while in the ITS phylogenetic tree, C. danxiacola formed a clade with C. armandii, and this combined clade was sister to a clade comprising C. terniflora, C. mandshurica Rupr. (1857: 258), C. chinensis, and C. hexapetala. This cyto-nuclear discordance aligns with previous research findings, suggesting widespread interspecific hybridization events in Clematis (Xiao et al. 2022). Morphologically, C. danxiacola shares the strongest similarities with C. terniflora; it bears some resemblance to C. chinensis and C. chinensis var. vestita (Rehder & E.H.Wilson) W.T. Wang (1998: 158) [≡ C. chinensis f. vestita Rehder & E.H.Wilson (1913: 330)], but maintains its natural color when dried, and exhibits distinct characteristics in its bracts and sepals (Table 3).
Table 3.: Morphological comparison with Clematis danxiacola sp. nov. and its allied taxa.
Conclusion
This study presents a new species of Clematis based on morphological and molecular evidence. The documented plastome provides valuable data for future research on the systematics, evolution, and conservation of the genus.
Supplementary Material
XML Treatment for Clematis danxiacola
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bankevich A Nurk S Antipov D Gurevich AA Dvorkin M Kulikov AS Lesin VM Nikolenko SI Pham S Prjibelski AD (2012) SP Ades: A new genome assembly algorithm and its applications to single-cell sequencing.Journal of Computational Biology 19: 455–477. 10.1089/cmb.2012.002122506599 PMC 3342519 · doi ↗ · pubmed ↗
- 2Bentham G (1861) Flora Hongkongensis: A Description of the Flowering Plants and Ferns of the Island of Hongkong.Lovell Reeve, London, 7 pp. 10.5962/bhl.title.21052 · doi ↗
- 3Brandenburg WA (2000) Meclatis in Clematis: Yellow flowering Clematis species. Systematic studies in Clematis I. (Ranunculaceae), inclusive of cultonomic aspects. Wageningen Universiteit, Wageningen.
- 4Chen LY Song MS Zha HG Li ZM (2014) A modified protocol for plant genome DNA extraction.Plant Diversity and Resources 36: 375–380.
- 5de Candolle AP (1818) Regni vegetabilis systema naturale I.Treuttel & Wurtz, Paris, 137 pp. 10.5962/bhl.title.59874 · doi ↗
- 6Franchet A (1885) Plantae Davidianae ex Sinarum Imperio: Plantes du Thibet Oriental, Province de Moupine. Nouvelles Archives du Muséum d’Histoire Naturelle, sér. 2, 8: 184.
- 7Greiner S Lehwark P Bock R (2019) Organellar Genome DRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Research 47: W 59–W 64. 10.1093/nar/gkz 238PMC 660250230949694 · doi ↗ · pubmed ↗
- 8Greuter W (1965) Beiträge zur Flora der Südägäis 1–7.Candollea 20: 212–213. 10.5169/seals-880361 · doi ↗