A species-specific tale of quantitative disease resistance (QDR) in tomato
Pei Qin Ng

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
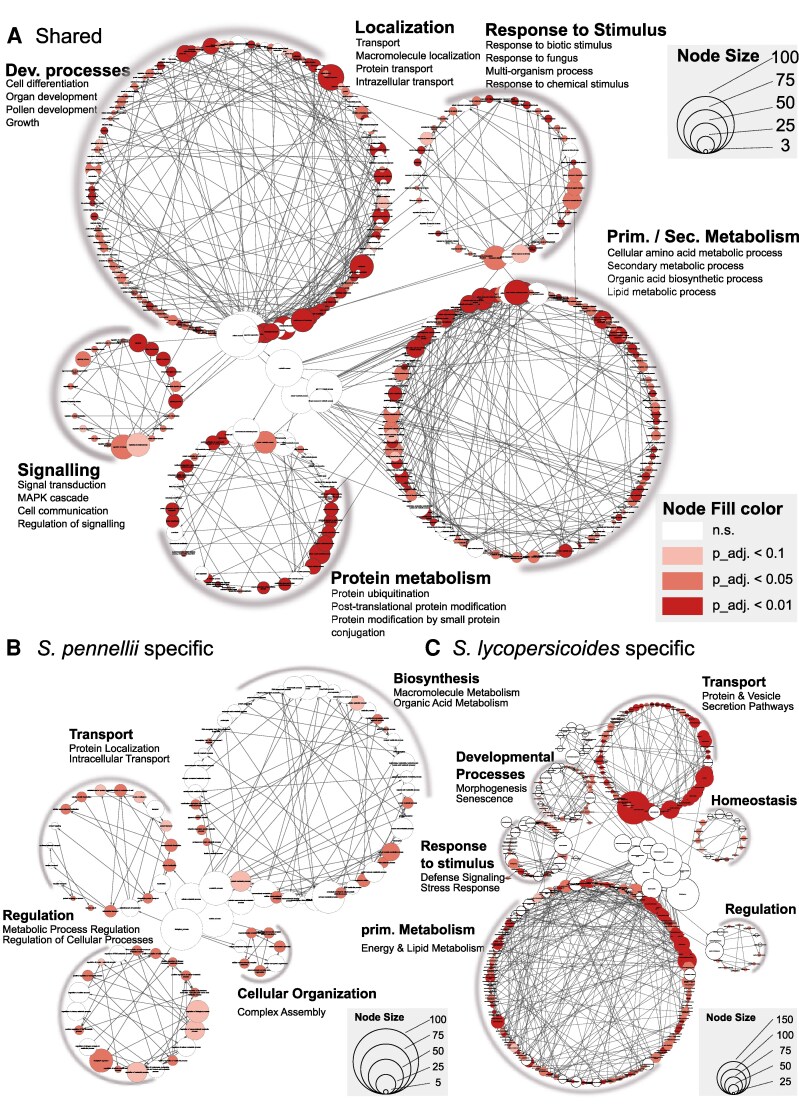 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Resistance · Fungal Plant Pathogen Control · Plant-Microbe Interactions and Immunity
Due to the molecular complexity and inefficacy of the gene-for-gene immune response, it is a considerable challenge to engineer disease-resistant crops against necrotrophic pathogens (Mengiste and Liao 2025). Quantitative disease resistance (QDR) is an alternative to the “all or nothing” gene-for-gene model in plants, in which QDR confers plants partial resistance to a broad spectrum of pathogens. Therefore, enhancing QDR in crop plants is critical to provide resistance against necrotrophic pathogens. Recent work by Delplace et al. (2025; highlighted by Ng [2025]) explored QDR diversity among Arabidopsis accessions, identifying core QDR genes involved in response to infection by the necrotrophic fungus Sclerotinia sclerotiorum.
In their new work, Severin Einspanier and colleagues (Einspanier et al. 2025) inspected the transcriptome of 5 tomato species to characterize the evolution of QDR regulation in response to S. sclerotiorum. Previously, the authors used a detached leaf assay to describe a diversity of resistance phenotypes among 4 wild tomato species (also used in the current study), with none showing 100% resistance to S. sclerotiorum (Einspanier et al. 2024). Before proceeding into in-depth transcriptomic analysis, the authors profiled the phylogenetic relationship of the 5 tomato species using protein sequence data. Lesion growth rate analysis revealed vastly different susceptibilities of respective cultivars. Through differential gene expression analysis, the authors concluded that the number of differentially expressed genes (DEGs) does not explain the QDR variability, as there were fewer DEGs for resistant versus susceptible S. lycopersicoides and vice versa for S. pennelli. The authors reported a total of 239 core DEG orthologues between the resistant and susceptible genotypes of all 5 tomato species, which followed diverse expression patterns upon S. sclerotium infection.
To understand the influence of gene network rewiring in QDR responses in susceptible versus resistant cultivars, the authors deployed weighted gene correlation network analysis on 2 species: S lycopersicoides and S. pennellii. This revealed clear transcriptional differences between susceptible and resistant genotypes, allowing the authors to describe a 2-tier QDR system comprised of a broad functioning defence network and species-specific responses (Figure). The analysis also showed that the shared genes from resistance modules possessed a lower-than-average transcript diversification index, indicating signs of purifying selection. The authors hypothesized that QDR regulation in S. lyocopersicoides and S. pennellii might therefore be driven by differential regulation (“rewiring”) of a relatively well-conserved, ancestral suite of genes.
The authors then performed an integrative analysis of weighted gene correlation network analysis, directed gene regulatory networks, and differential expression to identify central regulators of QDR in all tested species. These central regulators are often transcription factors forming the core of the transcription network (Sorrells and Johnson 2015). Interestingly, the NAC family TF NAC29 was identified as a key gene in mediating QDR responses toward S. sclerotiorum. The NAC transcription factors (TFs) are key players in plant developmental processes, such as embryogenesis, leaf senescence, and root development (Wen et al. 2022; Xiong et al. 2025). In this study, the role of NAC29 in QDR is unique to S. pennellii, while the same gene is not associated with QDR in the other species. In a highly susceptible genotype of S. pennellii, NAC29 contained a premature stop codon within the coding sequence, suggesting a new functional role of NAC29 in QDR resistance phenotype.
In summary, the work of Einspanier and colleagues on tomato presents early evidence of gene network rewiring in QDR against a necrotrophic pathogen, as well as the co-option of a TF in driving QDR through the species-specific case study of S. penelli. Future direction of this study will require generating a nac29 knockout in the S. penelli–resistant cultivar to confirm its role in S. sclerotiorum disease resistance. This demonstrates the complex gene regulation involved in plant disease resistance mechanisms via QDR and opens more avenues to our understanding of the evolution driving this defence mechanism.
Recent related articles in The Plant Cell:
Delplace et al. (2025) explored QDR toward Sclerotinia sclerotiorum in Arabidopsis thaliana accessions. Yue et al. (2024) used a genome-wide association study to identify a NAC TF variant associated with fruit ripening in apple.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Delplace F, Khafif M, Stam R, Barbacci A, Raffaele S. Neutral transcriptome rewiring promotes quantitative disease resistance evolvability at the species level. Plant Cell. 2025:37(6):koaf 105. 10.1093/plcell/koaf 10540341395 PMC 12159807 · doi ↗ · pubmed ↗
- 2Einspanier S, Tominello-Ramirez C, Delplace F, Stam R. Co-option of transcription factors drives evolution of quantitative disease resistance against a necrotrophic pathogen. Plant Cell. 2025:37(10):koaf 233. 10.1093/plcell/koaf 23341026897 PMC 12527349 · doi ↗ · pubmed ↗
- 3Einspanier S, Tominello-Ramirez C, Hasler M, Barbacci A, Raffaele S, Stam R. High-resolution disease phenotyping reveals distinct resistance mechanisms of tomato crop wild relatives against sclerotinia sclerotiorum. Plant Phenomics. 2024:6:0214. 10.34133/plantphenomics.021439105186 PMC 11298253 · doi ↗ · pubmed ↗
- 4Mengiste T, Liao C-J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens, 20 years later: what has changed? Annu Rev Phytopathol. 2025:63(1):279–308. 10.1146/annurev-phyto-121823-03113940465653 · doi ↗ · pubmed ↗
- 5Ng PQ . Quantitive disease resistance (QDR): the alternative to “all-or-nothing” strategy in plant immunity. Plant Cell. 2025:37(6):132. 10.1093/plcell/koaf 132PMC 1216458040397690 · doi ↗ · pubmed ↗
- 6Sorrells Trevor R, Johnson Alexander D. Making sense of transcription networks. Cell. 2015:161(4):714–723. 10.1016/j.cell.2015.04.01425957680 PMC 4531093 · doi ↗ · pubmed ↗
- 7Wen B, Gong X, Tan Q, Zhao W, Chen X, Li D, Li L, Xiao W. Md NAC 4 interacts with Md APRR 2 to regulate nitrogen deficiency-induced leaf senescence in apple (Malus domestica). Front Plant Sci. 2022:13:925035. 10.3389/fpls.2022.92503535845636 PMC 9280364 · doi ↗ · pubmed ↗
- 8Xiong H, He H, Chang Y, Miao B, Liu Z, Wang Q, Dong F, Xiong L. Multiple roles of NAC transcription factors in plant development and stress responses. J Integr Plant Biol. 2025:67(3):510–538. 10.1111/jipb.1385439950532 · doi ↗ · pubmed ↗