How tomatoes balance defense: Orchestration of gray mold immunity by the SlMYC2–SlLBD40/42–SlBPM4 module
Jiajun Wang, Yueyao Wang

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
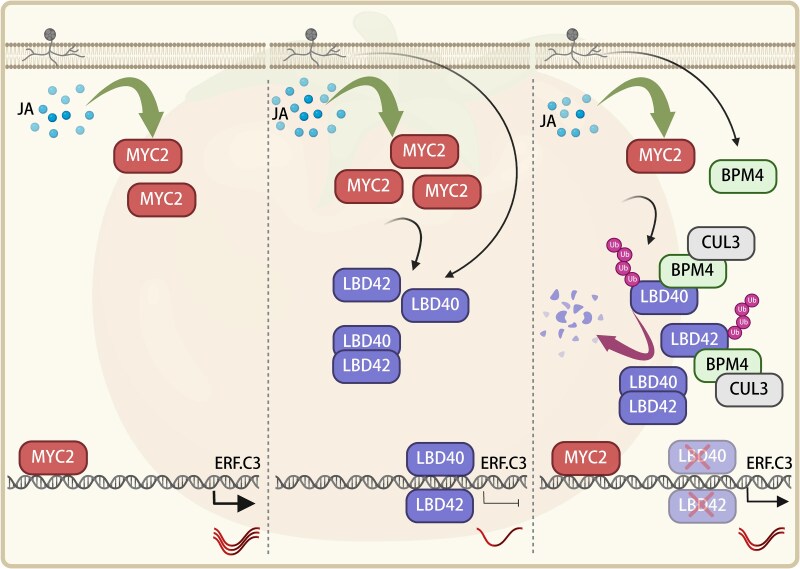 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Plant Pathogenic Bacteria Studies · Plant Virus Research Studies
Botrytis cinerea is a necrotrophic plant fungal pathogen, causing gray mold diseases on many economically important crops. It affects crops at both growing and post-harvest stages, and leading to severe economic losses (Bi et al. 2023). Infection by B. cinerea induces the accumulation of jasmonic acid–isoleucine (JA-Ile) in hosts. As an active signaling molecule, JA-Ile binds to the JA receptor COI1, thereby releasing the repression of bHLH transcription factors MYC2 and activating the JA defense signaling pathway (Macioszek et al. 2023). In tomato, SlMYC2 promotes the expression of numerous defense genes and the biosynthesis of secondary metabolites to resist B. cinerea, including the ethylene response factor (ERF) ERF.C3, which further activates the key defense gene PR-STH2 involved in plant defense responses (Du et al. 2017).
Lateral organ boundaries domain (LBD) proteins are a plant-specific transcription factor family. In tomato, SlLBD40 and SlMYC2 form a positive feedback regulatory loop to promote fruit enlargement. SlMYC2 activates SlLBD40 transcription, while SlLBD40 stabilizes SlMYC2 protein (Liu et al. 2024). In a new study, Jialong Zhang and colleagues (Zhang et al. 2025) report that SlLBD40/42 and SlMYC2 establish a negative feedback pathway that prevents tomato immunity from being overactivated during pathogen infection.
Similar to SlLBD40, the authors showed that SlMYC2 directly binds to the G-box (CACGTG) in the SlLBD42 promoter and activates its transcription. In contrast to the slmyc2 mutant, which is highly susceptible to B. cinerea, CRISPR/Cas9 knockout mutants of SlLBD40 and SlLBD42 exhibited resistant phenotypes in both leaves and fruits, whereas overexpression lines of SlLBD40 and SlLBD42 resulted in susceptibility. Moreover, sllbd40 slmyc2 and sllbd42 slmyc2 double mutants remained susceptible but to a lesser extent than slmyc2, indicating that SlLBD40/42 antagonize SlMYC2 in regulating tomato resistance to B. cinerea.
The authors found that SlLBD40 and SlLBD42 act as transcriptional repressors of defense gene expression, as infection with B. cinerea triggered strong upregulation of defense-related genes, including SlERF.C3, SlPR-STH2, SlTD, and SlJA2L, in sllbd40 and sllbd42 mutants, but the expression of these genes was suppressed in SlLBD40 and SlLBD42 overexpression lines. Both SlLBD40 and SlLBD42 contain an EAR (Ethylene-responsive element-binding factor–Associated Amphiphilic Repression) motif at their C terminus, and mutation of this motif completely abolished their transcriptional repression activity. Overexpressing SlLBD40/42 fused to the VP16 transcriptional activation domain exhibited significant resistance to B. cinerea, accompanied by significant upregulation of the B. cinerea-responsive gene SlERF.C3. These results demonstrate that the negative regulation of tomato resistance to B. cinerea by SlLBD40/42 is due to their transcriptional repression activity.
To further explore how SlLBD40 and SlLBD42 antagonize SlMYC2 in regulating B. cinerea resistance, yeast 1-hybrid, electrophoretic mobility shift, and chromatin immunoprecipitation–qPCR assays revealed that SlLBD40/42 bind to a PBE-box (CACATG) near the start codon of the SlERF.C3 promoter, whereas SlMYC2 binds to a distal PBE-box. Dual-luciferase reporter assays showed that SlLBD40/42 significantly suppress SlMYC2-mediated activation of the SlERF.C3 promoter. Consistent with the observation that loss of SlMYC2 compromised the enhanced resistance of the sllbd40 and sllbd42 mutants, the upregulation of SlERF.C3 transcripts in these mutants was largely decreased in the slmyc2 background. These genetic and molecular experiments demonstrate that SlLBD40/42 act to repress SlMYC2-mediated activation of target genes, thereby preventing excessive immune activation. Together with the finding that SlMYC2 positively regulates SlLBD40/42 transcription, these data establish a SlMYC2–SlLBD40/42 negative feedback loop in the regulation of B. cinerea resistance.
Through yeast 2-hybrid screening, the BTB/POZ protein SlBPM4 was identified as a SlLBD40/42 interacting protein. The authors confirmed this interaction in the nucleus using bimolecular fluorescence complementation, co-immunoprecipitation, dual-luciferase complementation imaging, and yeast 2-hybrid assays. Protein degradation assays showed that SlLBD40 and SlLBD42 are mainly degraded via the 26S proteasome pathway. In cell-free degradation assays, SlLBD40 and SlLBD42 were degraded significantly more slowly in the slbpm4 mutant, and their ubiquitination levels were markedly reduced. Similarly, during B. cinerea infection, SlLBD40/42 protein levels accumulated to higher levels in slbpm4 mutants compared with wild type. Furthermore, when SlLBD40/42 were coexpressed with SlBPM4, their protein levels decreased in a SlBPM4 dose-dependent manner, demonstrating that SlBPM4 functions as an E3 ubiquitin ligase to promote their ubiquitination and degradation. Genetic analyses further showed that slbpm4 mutants were more susceptible to B. cinerea, whereas SlBPM4 overexpression enhanced resistance. slbpm4 sllbd40 and slbpm4 sllbd42 double mutants resembled sllbd40 and sllbd42 single mutants in showing increased resistance, further supporting the conclusion that SlLBD40/42 act downstream of SlBPM4.
Interestingly, the authors found that upon B. cinerea infection, SlMYC2 transcript levels peaked at 24 h, followed by SlLBD40 and SlLBD42 at 36 h, while SlBPM4 peaked later at around 60 h. Protein accumulation of SlMYC2, SlLBD40/42, and SlBPM4 mirrored this temporal pattern. Therefore, upon B. cinerea infection, SlMYC2 activates SlLBD40/42 transcription, after which SlLBD40/42 suppress SlMYC2-mediated activation of defense genes, acting as brakes to prevent excessive immune activation. As infection progresses, SlBPM4 induction promotes the ubiquitin-dependent degradation of SlLBD40/42 via the 26S proteasome, thereby relieving their repression of SlMYC2 and ensuring a balanced defense response (Fig.).
Recent related articles in The Plant Cell:
Xu et al. (2023) revealed that auxin-induced WRKY23 activates PLT genes while LBD-mediated removal of the repressor bHLH041 derepresses them, and together these coordinated actions establish callus pluripotency in Arabidopsis regeneration. Zhang et al. (2025) demonstrated that jasmonate signaling regulates Arabidopsis seed size by coordinating JAZ/MYC-mediated repression with the SOD7-DPA4-KLU pathway, integrating hormonal and transcriptional control to modulate seed growth, especially under salinity stress. Wang et al. (2024) showed that jasmonate induces translation of the tRNA-binding protein YUELAO1 via its 3′ UTR, which activates MYC2 and thereby enhances jasmonate responses, revealing a translational layer of regulation in jasmonate signaling.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bi K, Liang Y, Mengiste T, Sharon A. Killing softly: a roadmap of Botrytis cinerea pathogenicity. Trends Plant Sci. 2023:28(2):211–222. 10.1016/j.tplants.2022.08.02436184487 · doi ↗ · pubmed ↗
- 2Du M, Zhao J, Tzeng DTW, Liu Y, Deng L, Yang T, Zhai Q, Wu F, Huang Z, Zhou M, et al MYC 2 orchestrates a hierarchical transcriptional cascade that regulates jasmonate-mediated plant immunity in tomato. Plant Cell. 2017:29(8):1883–1906. 10.1105/tpc.16.0095328733419 PMC 5590496 · doi ↗ · pubmed ↗
- 3Liu L, Zhang J, Xu J, Li Y, Lv H, Wang F, Guo J, Lin T, Zhao B, Li XX, et al Sl MYC 2 promotes Sl LBD 40-mediated cell expansion in tomato fruit development. Plant J. 2024:118(6):1872–1888. 10.1111/tpj.1671538481350 · doi ↗ · pubmed ↗
- 4Macioszek VK, Jecz T, Ciereszko I, Kononowicz AK. Jasmonic acid as a mediator in plant response to necrotrophic fungi. Cells. 2023:12(7):1027. 10.3390/cells 1207102737048100 PMC 10093439 · doi ↗ · pubmed ↗
- 5Wang J, Li Y, Hu Y, Zhu S. Jasmonate induces translation of the Arabidopsis transfer RNA-binding protein YUELAO 1, which activates MYC 2 in jasmonate signaling. Plant Cell. 2024:37(1):koae 294. 10.1093/plcell/koae 29439489485 PMC 11663558 · doi ↗ · pubmed ↗
- 6Xu C, Chang P, Guo S, Yang X, Liu X, Sui B, Yu D, Xin W, Hu Y. Transcriptional activation by WRKY 23 and derepression by removal of b HLH 041 coordinately establish callus pluripotency in Arabidopsis regeneration. Plant Cell. 2023:36(1):158–173. 10.1093/plcell/koad 25537804093 PMC 10734573 · doi ↗ · pubmed ↗
- 7Zhang J, Dong D, Jia Y, Li H, Liu L, Xu J, Cui H, Zhang N, Guo YD. Fine-tuning of MYC 2-mediated Botrytis defense response by LBD 40/42- CRL 3BPM 4 module in tomato. Plant Cell. 2025; 10.1093/plcell/koaf 25841134692 · doi ↗ · pubmed ↗
- 8Zhang J, Yao J, He K, Yu C, Du J, Du J, Fu Q, Yao R, Howe GA, Han X, et al Jasmonate signaling coordinates with the SOD 7-KLU pathway to regulate seed size in Arabidopsis thaliana. Plant Cell. 2025:37(8):koaf 178. 10.1093/plcell/koaf 17840679362 · doi ↗ · pubmed ↗