Silver‐selenium nanoparticles and selected chemical compounds significantly inhibit grapevine trunk disease pathogens
Kateřina Štůsková, Tomáš Kiss, Zuzana Bytešníková, Lukáš Richtera, David Gramaje, Ales Eichmeier

TL;DR
A 2-year study found that silver-selenium nanoparticles and certain chemicals can inhibit grapevine trunk disease pathogens without harming the plants.
Contribution
Silver-selenium nanoparticles show strong antifungal activity against specific GTD pathogens with no phytotoxic effects.
Findings
Silver-selenium nanoparticles inhibited D. eres and E. lata with 55.0% to 86.9% effectiveness.
Chemical treatments showed inhibition rates ranging from 33.6% to 93.7% against GTD pathogens.
No phytotoxic effects were observed at the tested nanoparticle concentrations.
Abstract
Grapevine is among the most economically important cultivated crops worldwide, yet it is increasingly threatened by the grapevine trunk disease (GTD) complex. Due to the lack of effective curative treatments for GTDs and the growing need to reduce chemical pesticide use, alternative strategies, such as the application of nanoparticles, are being investigated. In a 2‐year in planta study, the inhibitory effects of four chemical compounds and one nanoparticle formulation were evaluated against three serious pathogens associated with GTD complex: Diaporthe eres Nitschke, Diplodia seriata De Not., and Eutypa lata (Pers.) Tul. & C. Tul. Selection of the chemical compounds and nanoparticles was based on their inhibitory effects observed under in vitro conditions, as previously reported. All chemical treatments demonstrated antifungal activity, with inhibition rates ranging from 33.6% to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
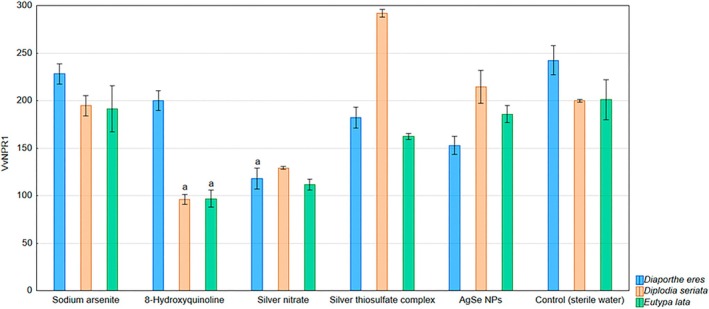 Figure 1
Figure 1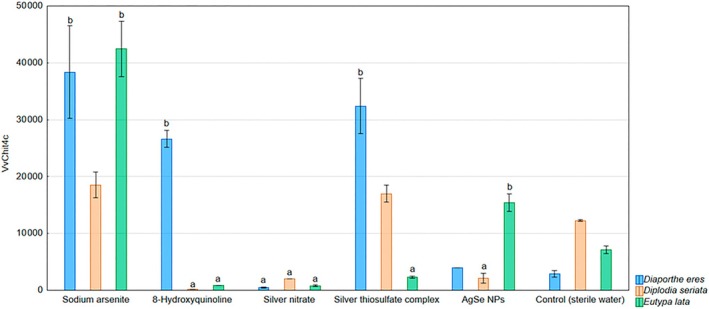 Figure 2
Figure 2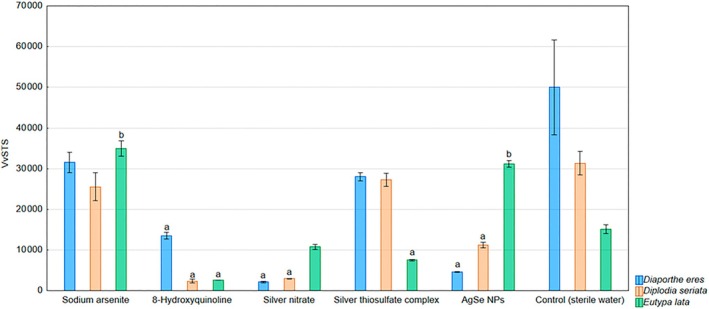 Figure 3
Figure 3| Name | Active ingredient | Concentration | Producer |
|---|---|---|---|
| Sodium arsenite | Sodium arsenite | 6.50 g l−1 | Merck KGaA, Darmstadt, Germany |
| 8‐Hydroxyquinoline | 8‐hydroxyquinoline | 0.05 g l−1 | Carl Roth GmbH & Co. KG, Karlsruhe, Germany |
| Silver nitrate | Ag+ | 1.00 g l−1 | Merck KGaA, Darmstadt, Germany |
| Silver thiosulfate complex | Ag+ |
5.15 g l−1Ag, 105.20 g l−1AgN, 217.50 g l−1AgS | Merck KGaA, Darmstadt, Germany |
| AgSe NPs | Ag, Se | 2.59 g l−1 Ag, 0.90 g l−1 Se | Department of Chemistry and Biochemistry, Mendel University, Brno, Czech Republic |
| Pathogen | Name of isolate | Host plant | Accession number | Source |
|---|---|---|---|---|
|
| Grapevine | Guarnaccia | ||
|
| OCR120 | Walnut | Eichmeier | |
|
| CS_12_27_4_KO | Grapevine | Baránek |
| Gene | Accession number | Forward primer 5′ → 3' | Reverse primer 3′ → 5' |
|---|---|---|---|
| VvACT | TGCTATCCTTCGTCTTGACCTTG | GGACTTCTGGACAACGGAATCTC | |
| VvNPR1 | GSVIVT00016536001 | GGAATTCGATGTTGGGTACG | GCAACCTTGTCAAGAATGTCC |
| VvSTS | CATCAAGGGTGCTATGCAGGT | TCAGAGCACACCACAAGAACTCG | |
| VvChit4c | TCGAATGCGATGGTGGAAA | TCCCCTGTCGAAACACCAAG |
| Sodium arsenite | 8‐Hydroxy‐quinoline | Silver nitrate | Silver thiosulfate complex | AgSe NPs | Control (sterile water) | |||
|---|---|---|---|---|---|---|---|---|
|
| Grapevine height (cm) | 2021 | 65.67 | 82.25 | 83.83 | 84.33 | 74.25 | 61.67 |
| 2022 | 110.25 | 125.88 | 102.25 | 138.00 | 110.75 | 105.75 | ||
| Root length (cm) | 2021 | 32.67 | 34.25 | 40.83 | 33.17 | 32.63 | 19.67 | |
| 2022 | 35.75 | 34.00 | 29.75 | 31.50 | 29.75 | 26.50 | ||
| Necrosis (%) | 2021 | 22.40 | 30.71 | 7.71 | 8.81 | 7.65 | 58.21 | |
| 2022 | 52.34 | 26.88 | 41.69 | 19.80 | 31.97 | 98.39 | ||
|
| Grapevine height (cm) | 2021 | 55.25 | 63.00 | 64.83 | 64.75 | 91.17 | 60.63 |
| 2022 | 106.00 | 110.00 | 105.75 | 105.00 | 118.50 | 110.50 | ||
| Root length (cm) | 2021 | 30.00 | 29.00 | 37.17 | 31.00 | 37.67 | 26.88 | |
| 2022 | 34.50 | 29.75 | 25.00 | 39.25 | 33.25 | 24.75 | ||
| Necrosis (%) | 2021 | 27.46 | 3.51 | 10.98 | 8.67 | 27.90 | 55.26 | |
| 2022 | 43.39 | 28.52 | 54.61 | 46.53 | 65.03 | 84.69 | ||
|
| Grapevine height (cm) | 2021 | 91.00 | 63.67 | 72.00 | 56.67 | N/A | 73.00 |
| 2022 | 126.25 | 96.00 | 94.50 | 106.50 | 115.00 | 83.50 | ||
| Root length (cm) | 2021 | 38.00 | 25.33 | 35.00 | 28.00 | N/A | 32.83 | |
| 2022 | 30.50 | 25.75 | 36.25 | 27.25 | 32.00 | 23.50 | ||
| Necrosis (%) | 2021 | 19.76 | 4.74 | 8.97 | 10.72 | N/A | 34.11 | |
| 2022 | 55.80 | 12.45 | 48.41 | 29.33 | 37.83 | 84.08 |
| Number of plants | |||||||
|---|---|---|---|---|---|---|---|
|
|
|
| |||||
| Substance | Applied pathogen | 2021 | 2022 | 2021 | 2022 | 2021 | 2022 |
| Sodium arsenite |
| 3 | 4 | 0 | 0 | 0 | 0 |
|
| 0 | 1 | 3 | 3 | 1 | 0 | |
|
| 0 | 1 | 0 | 0 | 4 | 4 | |
| 8‐Hydroxyquinoline |
| 3 | 4 | 1 | 0 | 0 | 0 |
|
| 0 | 0 | 3 | 4 | 0 | 0 | |
|
| 0 | 1 | 0 | 0 | 3 | 3 | |
| Silver nitrate |
| 3 | 4 | 1 | 0 | 0 | 1 |
|
| 0 | 1 | 4 | 3 | 0 | 0 | |
|
| 0 | 0 | 1 | 0 | 3 | 4 | |
| Silver thiosulfate complex |
| 4 | 4 | 1 | 0 | 0 | 0 |
|
| 1 | 0 | 4 | 4 | 1 | 0 | |
|
| 0 | 0 | 1 | 0 | 4 | 4 | |
| AgSe NPs |
| 4 | 4 | 0 | 1 | 1 | 0 |
|
| 0 | 1 | 4 | 4 | 0 | 1 | |
|
| N/A | 0 | N/A | 0 | N/A | 4 | |
| Control (sterile water) |
| 4 | 3 | 1 | 1 | 0 | 0 |
|
| 0 | 1 | 4 | 4 | 1 | 0 | |
|
| 1 | 1 | 1 | 0 | 3 | 4 | |
- —European Cooperation in Science and Technology10.13039/501100000921
- —José Castillejo
- —Ministerstvo Zemědělství10.13039/501100006533
- —Ministry of Agriculture of the Czech Republic10.13039/501100006533
- —‘José Castillejo’ Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNematode management and characterization studies · Plant Pathogens and Fungal Diseases · Plant pathogens and resistance mechanisms
INTRODUCTION
1
Grapevine trunk diseases (GTDs) represent a complex of diseases primarily associated with fungal pathogens.1, 2 The GTD complex encompasses six main diseases: the Esca complex, Eutypa dieback, Botryosphaeria dieback, Phomopsis dieback, Petri disease, and black foot disease.2, 3 Diagnosing GTDs can be particularly challenging due to the sporadic appearance of symptoms and the overlap of both external and internal manifestations.4 The pathogens selected for the in planta experiment in this study were Diaporthe eres, Diplodia seriata, and Eutypa lata, each associated with a distinct form of grapevine trunk diseases. D. eres is typically associated with Phomopsis dieback (also known as cane and leaf spot or perennial cankers). This disease is characterized by the formation of black lesions on shoots, followed by progressive wilting and eventual necrosis.3, 5, 6 D. seriata is a pathogen responsible for Botryosphaeria dieback, which manifests as necrosis of woody tissues, bark cracking, and gradual dieback of branches.3 E. lata causes Eutypiosis, characterized by deformation and shortening of shoots, presence of necrotic tissues with distinctive dark streaking in the wood, and progressive death of affected plant parts.3, 4 GTDs pose significant health and economic threats to grapevines and are present in all major grape‐growing regions worldwide.7
The treatment of GTDs remains highly challenging, particularly following the ban of sodium arsenite in 2003 due to its environmental and human health risks.8 Complete eradication of GTDs using chemical or biocontrol agents is not possible; therefore, control strategies are primarily aimed at prevention and mitigation of symptoms.3, 9, 10 In the Czech Republic, the protection of grapevines against fungal diseases is strictly regulated and subject to limitations. Although plant protection products targeting GTDs are registered and in use in several regions worldwide, particularly in the Americas and Australia, no such formulation is currently approved for use in the Czech Republic.11 Overall, the number of chemical agents approved for GTD control within the European Union remains very limited. Additionally, there is a growing emphasis on the use of biological control agents, particularly the application of fungi from the genus Trichoderma 12, 13, 14, 15, 16, 17 or bacteria of the genera Bacillus 18, 19, 20 and Streptomyces.19, 21 As a result, chemical solutions are becoming increasingly restricted, and their registration and approval vary across member states.22
Nanoparticles (NPs) and nanomaterials (NMs) represent a highly promising and extensively studied field. NMs represent a suitable approach to reducing the use of conventional chemical protection. Within the context of modern agriculture, NPs offer considerable potential to enhance plant resistance to biotic stressors, including pests and pathogens, while simultaneously improving water‐use efficiency, promoting vegetative and reproductive growth, and facilitating plant adaptation to the impacts of climate change. Moreover, their application may significantly reduce the ecological footprint of agricultural systems and contribute to sustainable solutions in the face of the global food security crisis.23, 24, 25, 26 Nanomaterials have increasingly attracted attention in the field of agriculture for their potential application in the sustainable remediation of soil degradation. Due to their high surface area‐to‐volume ratio, adjustable physicochemical properties, and enhanced reactivity, they have recently been identified as promising candidates for nano‐enabled fertilizers. These nanofertilizers exhibit superior bioavailability and improved nutrient delivery efficiency, allowing for more effective plant uptake compared to conventional bulk formulations. Their application may thus contribute not only to soil restoration but also to increased nutrient use efficiency and reduced environmental impact.27, 28, 29
In a previously published study, an in vitro assay was conducted to evaluate the antifungal efficacy of newly synthesized silver selenium (AgSe) and copper selenium (CuSe) NPs, as well as mono‐elemental NPs of silver (Ag NPs), copper (Cu NPs), and selenium (Se NPs).1 In addition to selected chemical agents including sodium arsenite, 8‐hydroxyquinoline, silver nitrate, colloidal silver, Altron Silver fertilizer, and the silver thiosulfate complex (NH_4_)3/Ag(S_2_O3)2/. These compounds were tested against three prevalent grapevine trunk pathogens (GTPs) – D. eres, E. lata, and D. seriata. Among the tested agents, silver nitrate and AgSe NPs exhibited the most pronounced antifungal activity. These results highlight the potential of Ag‐based nanomaterials, particularly AgSe NPs, as effective antifungal agents against GTPs.1
Based on the results of the previous in vitro study,1 the current in planta experiment included a selection of treatments: sodium arsenite, hydroxyquinoline sulfate (8‐HCH), silver nitrate, silver thiosulfate complex, and AgSe NPs. Each compound was applied at the concentrations that exhibited the highest inhibitory effect in the earlier in vitro experiment.1 We hypothesized that (i) the selected chemical compounds and NPs would exhibit antifungal activity against GTD pathogens, and (ii) their application would alter the gene expression profile of grapevine.
MATERIALS AND METHODS
2
Chemical substances and nanomaterial for inhibition experiment
2.1
The chemical compounds and NPs used in this study are listed in Table 1. Sodium arsenite, silver nitrate, and silver thiosulfate complex were purchased from Merck KGaA (Darmstadt, Germany), while 8‐HCH was obtained from Carl Roth GmbH & Co. KG (Karlsruhe, Germany). Sodium arsenite was tested at a concentration of 6.50 g l^−1^, and 8‐HCH at 0.05 g l^−1^. Silver nitrate was applied at a concentration of 1.00 g l^−1^. The silver thiosulfate complex was tested at 100% concentration, containing 5.15 g l^−1^ Ag, 105.20 g l^−1^ AgN, and 217.50 g l^−1^ AgS. The AgSe NPs were newly synthesized by the Department of Chemistry and Biochemistry at Mendel University in Brno. Details of the synthesis are described by Štůsková et al. (2022).1 The AgSe NPs, tested at 100% concentration, contained 2.59 g l^−1^ Ag and 0.90 g l^−1^ Se.
Fungal isolates
2.2
The selected fungal pathogens are listed in Table 2. All strains are maintained in culture collections at Mendeleum – Institute of Genetics, Mendel University in Brno (Lednice, Czech Republic) or CBS‐KNAW (Utrecht, Netherlands). Three GTD pathogens were selected: D. eres CPC 28220,6 D. seriata OCR120,30 and E. lata CS_12_27_4_KO.31 D. eres and E. lata were isolated from grapevine wood, while D. seriata was isolated from walnut wood. All three pathogens are polyphagous and known to infect a wide range of plant hosts, including grapevines, pears, walnuts, blackberries, and mulberries.30, 32, 33
Inhibitory activity
2.3
The in planta experiment was conducted over two consecutive years (2021 and 2022) in a net house at Mendeleum – Institute of Genetics, Mendel University in Brno, Lednice, Czech Republic. The grapevine cultivar Sauvignon Blanc, grafted onto SO4 rootstock, was selected for the experiment due to its known high susceptibility to GTD pathogens.34, 35, 36 To simulate infection and evaluate the inhibitory effect of the treatments, the stem of each grafted vine was carefully peeled in the upper part, approximately 5–10 cm below the apex, using a sterile scalpel to expose the underlying tissue. At this site, a 3 mm diameter hole was created with a cork borer for the application of mycelial plugs.37 Chemicals or NPs were applied directly into the wound using a sterile brush. Sterile water served as the positive control. Each treatment was applied to four plants (replicates). Fungal discs (3 mm in diameter) were excised from active pathogen cultures and aseptically placed onto the treated wound sites. The inoculation site was covered with moistened cotton wool and sealed with Parafilm to maintain humidity. The grapevines were planted in containers and maintained under net house conditions, with regular monitoring and irrigation as needed. After 6 months, grapevines were removed from the containers and roots were thoroughly washed with tap water. Measurement included total grapevine height (stem height + length of the longest cane) and root length. To assess internal disease progression, bark at the inoculation site was removed and wood necrosis was measured. Necrosis was expressed as a percentage of stem height using the formula:
where x = percentage of necrosis, n = necrosis size (cm), and s = stem height (cm). Pathogens were re‐isolated from the edge of necrotic tissues to confirm infection. Additionally, RNA was subsequently isolated from the wood for subsequent gene expression analysis. The percentage of inhibition was calculated using the formula:
where I is the percentage of inhibition, r c is the average percentage of necrosis in the control group, and r t is the average percentage of necrosis in the treated group.1
Statistical analysis
2.4
Statistical analysis was performed using Statistica 14 CZ (StatSoft, Prague, Czech Republic). For each treatment, four plants were planted resulting in four numerical measurements per parameter: plant height (cm), root length (cm), and necrosis size (cm). A one‐way analysis of variance (ANOVA) was conducted separately for each parameter at a significance level of P = 0.05. When significant differences were detected, a post hoc least significant difference (LSD) test was applied at the same significance threshold (P = 0.05). The LSD test was used to assess the significance of differences between each treatment and the control.
Pathogen re‐isolation and identification
2.5
For pathogen re‐isolation, necrotic wood was surface‐sterilized by soaking in 1% sodium hypochlorite for 3 min, rinsed three times with distilled water, and then cut into small pieces using a sterile scalpel. Wood fragments were aseptically transferred onto potato dextrose agar (PDA; HiMedia, Čaderský‐Envitek, Brno, Czech Republic) supplemented with streptomycin (HiMedia, Čaderský‐Envitek, Brno, Czech Republic) at a concentration of 100 mg/L to inhibit bacterial growth. Emerging fungal colonies were subcultured onto fresh PDA plates to obtain pure cultures. These pure cultures were then grouped into three categories based on morphological characteristics: the first group included cultures morphologically consistent with Diaporthe, the second with Diplodia, and the third with Eutypa. One representative culture from each group was selected, and mycelium was collected for DNA extraction and subsequent sequencing to confirm the genus. To identify the reisolated fungi at the genetic level, genomic DNA was extracted from selected pure cultures using the NucleoSpin Tissue Kit (Macherey‐Nagel, Düren, Germany). Amplification of the internal transcribed spacer (ITS) region was performed using the universal fungal primers ITS1 (5’‐TCC GTA GGT GAA CCT GCG G‐3′) and ITS4 (5’‐TCC TCC GCT TAT TGA TAT GC‐3′).38 Diaporthe was identified to the species level based on beta‐tubulin (tub2) and translation elongation factor 1‐alpha (tef1) sequences, while Diplodia and Eutypa were identified based on the tub2 sequence, following Eichmeier et al. (2020).30 Sample preparation for sequencing was carried out using the Mix2Seq Kit (Eurofins Genomics, Ebersberg, Germany), and sequencing was performed by Eurofins Genomics. Resulting sequences were compared to reference sequences using the BLAST algorithm39 using GenBank NCBI to determine fungal identity.
Gene expression analysis
2.6
Gene expression analysis was performed only in the second year of the experiment, following the significant inhibition effects observed during the first year. To gain deeper insights into the interaction between the pathogen, the plant, and the applied substance, the second‐year analysis focused on the plant's molecular response.
Four marker genes were selected for gene expression analysis: actin (VvACT), non‐expresser of pathogenesis‐related gene 1 (VvNPR1), stilbene synthase (VvSTS), and acidic class IV chitinase (VvChit4c). The VvACT gene served as a housekeeping gene to normalize the expression levels of the target genes. Gene expression was evaluated using real‐time PCR (qPCR). Primer sequences are listed in Table 3. Total RNA was extracted using the Spectrum™ Plant Total RNA Kit (Sigma‐Aldrich, St. Louis, Missouri, USA). RNA concentrations were measured using a SpectrostarNano (BMG Labtech, Ortenberg, Germany) and adjusted to a uniform concentration of 90 ng/μL using sterile nuclease‐free water (Ambion, Pleasanton, California, USA). Reverse transcription was performed following the protocol described by Eichmeier et al. (2010),40 using the RevertAid RT Reverse Transcription Kit (Thermo Fisher Scientific, Waltham, Massachusetts, USA), resulting in complementary DNA (cDNA) samples. qPCR was carried out using Genaxxon mastermix (Genaxxon Bioscience GmbH, Ulm, Germany) on a qTower3 real‐time PCR cycler (Analytik Jena, Jena, Germany), with gene‐specific thermal cycling protocols outlines in Table S1. Fluorescence signals and data analysis were processed using qPCRsoft 3.4 software (Analytik Jena). Statistical analysis of normalized gene expression levels was conducted using Statistica 14 CZ. A gene was considered upregulated when its normalized expression exceeded a 2‐fold increase, and downregulated when it dropped below 0.5‐fold, relative to the untreated controls.
RESULTS
3
Inhibitory activity
3.1
The measured values of grapevine height (cm), root length (cm), and necrosis size (%) are shown in Table 4, while inhibition percentages are presented in Table S2.
In grapevine inoculated with D. eres, a statistically significant increase in grapevine height was observed only in 2021 for the treatment with silver thiosulfate complex, which reached 84.33 cm compared to 61.67 cm in the sterile water control. However, in 2022, no treatment, including silver thiosulfate complex, significantly affected grapevine height. Conversely, root length was significantly influenced by all treatments in the first year, with values ranging from 32.63 to 40.83 cm, compared to 19.67 cm in the control. In the second year, only sodium arsenite significantly increased root length, with a value of 35.75 cm versus 26.50 cm in the control. All tested chemical substances and nanoparticles had a significant effect on necrosis size and inhibition percentage in both years. In 2021, necrosis sizes for sodium arsenite, 8_HCH, silver nitrate, silver thiosulfate complex, and AgSe NPs were 22.40%, 30.71%, 7.71%, 8.81%, and 7.65%, respectively, compared to 58.21% in the control. The corresponding inhibition percentages were 61.51% for sodium arsenite, 47.24% for 8‐HCH, 86.75% for silver nitrate, 84.87% for silver thiosulfate complex, and 86.85% for AgSe NPs. In 2022, necrosis sizes were 52.34%, 26.88%, 41.69%, 19.80%, and 31.97% for sodium arsenite, 8‐HCH, silver nitrate, silver thiosulfate complex, and AgSe NPs, respectively, while the control showed 98.39% necrosis. Corresponding inhibition percentages were 46.80% for sodium arsenite, 72.68% for 8‐HCH, 57.63% for silver nitrate, 79.87% for silver thiosulfate complex, and 67.51% for AgSe NPs (Fig. S1).
In grapevines inoculated with D. seriata, a statistically significant increase in grapevine height was observed only in 2021 for the treatment with AgSe NPs, where the average height reached 91.17 cm compared to 60.63 cm in the control group treated with sterile water. In 2022, none of the treatment groups had a significant effect on grapevine height. Root length was not significantly affected by any of the selected chemicals or NPs in the first year. However, in 2022, sodium arsenite (34.50 cm), silver thiosulfate complex (39.25 cm), and AgSe NPs (33.25 cm) significantly increased root length relative to the control (24.75 cm). Necrosis size and inhibition percentage were significantly affected by all tested chemical treatments in both years. However, neither of the NP treatments has a statistically significant effect on these parameters in either year. In 2021, necrosis sizes for sodium arsenite, 8‐HCH, silver nitrate, and silver thiosulfate complex were 27.46%, 3.51%, 10.98%, and 8.67%, respectively, compared to 55.26% in the control. Corresponding inhibition percentages were 50.30% for sodium arsenite, 93.65% for 8‐HCH, 80.13% for silver nitrate, and 84.31% for silver thiosulfate complex. In 2022, necrosis sizes for sodium arsenite, 8‐HCH, silver nitrate, and silver thiosulfate complex were 43.39%, 28.52%, 54.61%, and 46.53%, respectively, compared to 84.69% in the control. The corresponding inhibition percentages were 48.77% for sodium arsenite, 66.32% for 8‐HCH, 35.52% for silver nitrate, and 45.06% for silver thiosulfate complex (Fig. S2).
In grapevines inoculated with E. lata, the AgSe NPs treatment was not evaluated in the first year due to dieback of three out of four grapevines, which precluded statistical analysis. In 2021, none of the chemical treatments significantly affected grapevine height. However, in 2022, significant increases in grapevine height were observed with sodium arsenite (126.25 cm) and AgSe NPs (115.0 cm), compared to the control (83.50 cm). Root length was not significantly affected by any treatment in 2021. In contrast, in 2022, root length was significantly increased by sodium arsenite (30.50 cm), silver nitrate (36.25 cm), and AgSe NPs (32.00 cm), compared to the control (23.50 cm). Necrosis size and corresponding inhibition percentage were significantly affected by all tested chemicals in the first year. Necrosis sizes were 19.76% for sodium arsenite, 4.74% for 8‐HCH, 8.97% for silver nitrate, and 10.72% for silver thiosulfate complex, compared to 34.11% in the control. Corresponding inhibition percentages were 42.08% for sodium arsenite, 86.11% for 8‐HCH, 73.72% for silver nitrate, and 63.58% for silver thiosulfate complex. In 2022, necrosis sizes were 55.80% for sodium arsenite, 12.45% for 8‐HCH, 48.41% for silver nitrate, 29.33% for silver thiosulfate complex, and 37.83% for AgSe NPs, while the control recorded 84.08%. The inhibition percentages were 33.64% for sodium arsenite, 85.18% for 8‐HCH, 42.43% for silver nitrate, 65.11% for silver thiosulfate complex, and 55.01% for AgSe NPs (Fig. S3).
Re‐isolation of pathogens
3.2
The number of plants from which pathogens were successfully re‐isolated is presented in Table 5. The re‐isolation method confirmed the presence of all inoculated pathogens, verifying successful infection of the grapevines. In both years, the target pathogens were re‐isolated from 3 to 4 plants per treatment (75–100%), thereby fulfilling Koch's postulates.
Gene expression analysis
3.3
In the analysis of gene expression, three genes involved in the plant‐pathogen response were monitored. The VvNPR1 gene was downregulated in response to 8‐HCH and silver nitrate.
8‐HCH downregulated VvNPR1 in grapevines inoculated with the pathogens D. seriata and E. lata, while in the case of silver nitrate, downregulation was observed in D. eres. In other treatments, the expression of this gene was not affected (Fig. 1).
Expression levels of the gene VvNPR1 monitored in the in planta experiment evaluating the inhibitory effects of selected chemical compounds and nanoparticles against the grapevine trunk disease pathogens D. eres, D. seriata, and E. lata: a – the gene was downregulated. There was no upregulation of the VvNPR1 gene in any of the treated groups.
The VvChit4c gene was upregulated in grapevines treated with sodium arsenite and inoculated with D. eres and E. lata. In grapevines treated with 8‐HCH and inoculated with D. seriata and E. lata, downregulation was observed, whereas in the case of D. eres, VvChit4c was upregulated. The silver nitrate treatment exhibited downregulations for all tested pathogens. Silver thiosulfate complex increased expression in grapevines inoculated with D. eres but decreased expression in those inoculated with E. lata. AgSe NPs reduced VvChit4c expression in grapevines inoculated with D. seriata, whereas in those inoculated with E. lata, expression was increased (Fig. 2).
Expression levels of the gene VvChit4c monitored in the in planta experiment evaluating the inhibitory effects of selected chemical compounds and nanoparticles against the grapevine trunk disease pathogens D. eres, D. seriata, and E. lata: a – the gene was downregulated; b – the gene was upregulated.
The last evaluated gene expression was the stilbene synthesis gene VvSTS. The expression of this gene was significantly reduced by 8‐HCH, silver nitrate, silver thiosulfate complex, and AgSe NPs. 8‐HCH downregulated expression in all three tested pathogens, while silver nitrate did not affect expression in grapevines inoculated with E. lata. Silver thiosulfate complex downregulated VvSTS expression in grapevines inoculated with E. lata. AgSe NPs reduced expression in grapevines inoculated with D. eres and D. seriata, whereas in grapevines inoculated with E. lata, expression was significantly increased. In the case of E. lata, expression was also increased by sodium arsenite application (Fig. 3).
Expression levels of the gene VvSTS monitored in the in planta experiment evaluating the inhibitory effects of selected chemical compounds and nanoparticles against the grapevine trunk disease pathogens D. eres, D. seriata, and E. lata: a – the gene was downregulated; b – the gene was upregulated.
This study also confirmed that the nanoparticles used did not exhibit any phytotoxicity at the applied concentrations.
DISCUSSION
4
The application of nanotechnology in agriculture has presented a major revolution. The increasing pressure to intensify crop yield is focusing on the application of nano‐biotechnologies in agriculture to improve the food market in a sustainable manner. This study evaluated the in planta antifungal effects of four chemical compounds and one nanoparticle formulation over 2 years against key GTD pathogens: Diaporthe eres, Diplodia seriata, and Eutypa lata. All tested treatments showed inhibitory effects, with silver‐selenium nanoparticles proving particularly effective against D. eres and E. lata, and without phytotoxic effects. The findings support the potential of nanoparticles as an environmentally friendly alternative to conventional plant protection in viticulture.
In our study, sodium arsenite significantly inhibited all tested GTD pathogens. This inhibitory effect has also been reported in other studies. Trouvelot et al. (2023)41 observed that grapevines treated with sodium arsenite did not develop foliar symptoms of Esca. Moreover, their study confirmed the impact of sodium arsenite on the metabolome, including the production of defensive secondary metabolites and histological changes in the interaction zone where the pathogen challenge occurred. In addition to its direct antifungal activity, sodium arsenite has been shown to stimulate the production of plant secondary metabolites associated with plant defense, particularly within woody tissues. This stimulatory effect on host tolerance and metabolic responses was also reported by Songy et al. (2019).42 Moreover, Bruez et al. (2021)43 demonstrated the inhibitory effect of sodium arsenite on Fomitiporia mediterranea, basidiomycete species involved in the Esca complex. In their study on the grapevine wood mycobiome, F. mediterranea showed a relative abundance of 63–94% in the control group, compared to only 1–40% in the group treated with sodium arsenite. A reduction in D. seriata and E. lata was also observed in the treated plants; however, it was not as pronounced as in the case of F. mediterranea. For D. eres, our study was the first to confirm the inhibitory effect of sodium arsenite against this pathogen.
The antifungal activity of 8‐HCH against GTD pathogens has also been confirmed in previous studies. Castillo‐Pando et al. (1997)44 reported that 8‐HCH, applied at a concentration of 1.5 g l^−1^ to one‐year‐old grapevine canes infected with Diaporthe ampelina, which causes Phomopsis dieback, reduced pycnidia viability to 0.1%, compared to 100% viability in the untreated control. Similarly, Fourie and Halleen (2006)45 observed a reduction in GTD pathogen incidence to 13.1% in grapevines treated with Chinosol (8‐HCH), in contrast to 26.4% in the untreated controls. In addition, Souza et al. (2021)46 demonstrated the efficacy of a synthetic derivative of 8‐HCH PH 151‐8‐hydroxyquinoline‐5‐(N‐4‐chlorophenyl) sulfonamide against Ilyonectria liriodendri in an in vivo experiment. At a concentration of 0.625 g l^−1^, PH 151 resulted in only 17% re‐isolation of the pathogen, whereas the commercial fungicide Mancozeb, used at the same concentration, showed a re‐isolation rate of 82%. The positive control showed 90% re‐isolation, highlighting the superior inhibitory effect of the 8‐HCH derivative.
Silver nitrate also demonstrated inhibitory activity against GTD pathogens in our study; however, its use on grapevine has not been previously reported. Nonetheless, several studies have confirmed its antimicrobial properties against other plant pathogens. For example, Ganiyu et al. (2024)47 reported that foliar application of silver nitrate at a concentration of 0.1 g l^−1^ on tomato plants significantly reduced the incidence of tomato wilt caused by Fusarium oxysporum f.sp. lycopersici to 10% compared to 89.90% in untreated controls. Similary, Oyelakin et al. (2023)48 observed that application of silver nitrate 16.99 g l^−1^ on tomato plants (cv. Roma VF) reduced the incidence of late blight caused by Phytophthora infestans from 53.33% in control plants to 33.30% in treated plants. In the 82B tomato cultivar, the reduction was even more substantial, from 60% in controls to 23.33% in treated plants. Additionally, Jo et al. (2009)49 demonstrated significant antifungal effects of silver nitrate against Bipolaris sorokiniana Shoemaker and Magnaporthe grisea in perennial ryegrass (Lolium perenne L.). When applied at a concentration of 0.05 g l^−1^, silver nitrate reduced leave damage to 7% compared to 70% in the sterile water control.
In our study, the in planta experiment demonstrated that the silver thiosulfate complex exhibited statistically significant inhibitory activity against all three tested GTD pathogens: D. eres, D. seriata, and E. lata. To our knowledge, no previous studies have investigated the antifungal activity of the silver thiosulfate complex against plant‐pathogenic fungi, making this one of the first reports highlighting its potential in plant disease management.
AgSe NPs demonstrated significant inhibitory activity against D. eres and E. lata, but not against D. seriata. Currently, no studies have directly assessed the efficacy of AgSe NPs against grapevine pathogens; however, several studies confirm the antifungal effects of individual Ag and Se NPs on fungal pathogens affecting other cultivated crops. Malandrakis et al. (2019)50 reported that Ag NPs at a concentration of 0.001 g l^−1^, achieved 100% inhibition of Botrytis cinerea on plum (Prunus domestica L.) fruits. Similarly, Jebril et al. (2020)51 evaluated the in vivo efficacy of Ag NPs at 0.02 g l^−1^, applied via soil irrigation after inoculation with Verticillium dahlia in eggplant (Solanum melongena L.) and observed an 87% reduction in diseases severity compared to untreated controls. The antifungal potential of Se NPs has also been reported. Hashem et al. (2021)52 found that Se NPs applied at 0.005 g l^−1^ reduced the disease index in faba bean (Vicia faba L.) plants infected with Rhizoctonia solani from 88% (control) to 20%. Likewise, Nandini et al. (2017)53 demonstrated that treating pearl millet (Cenchrus americanus) seeds with Se NPs (0.1 g l^−1^) significantly reduced the incidence of downy mildew caused by Sclerospora graminicola, from 93% in untreated plants to 49% in treated ones.
The expression patterns of three defense‐related genes (VvNPR1, VvChit4c, and VvSTS) revealed distinct responses depending on the applied treatment and the GTD pathogen involved, indicating a complex interaction between the grapevine defense system and the applied compounds.
The gene VvNPR1, a key regulator of systemic acquired resistance, was downregulated by 8‐HCH and silver nitrate, suggesting a possible suppression of salicylic acid‐mediated signaling. Notably, 8‐HCH suppressed VvNPR1 expression in plants inoculated with D. seriata and E. lata, while silver nitrate had a similar effect in plants inoculated with D. eres. In other treatments, VvNPR1 expression remained stable, indicating that these compounds selectively affect signaling pathways depending on the pathogen. The effect of chemical compounds on the upregulation of VvNPR1 expression has also been observed in other studies. An increase in VvNPR1 expression was recorded following the application of benzothiadiazole, both in grapevine cell cultures54 and in Vitis vinifera cv. Chardonnay plants.55 A transient increase in VvNPR1 expression was also observed following postharvest treatment of grape berries with β‐aminobutyric acid.56
The expression of VvChit4c, which encodes a class IV chitinase and is associated with fungal cell wall degradation, showed greater variability. Sodium arsenite significantly upregulated VvChit4c expression in grapevines inoculated with D. eres and E. lata, suggesting activation of defense mechanisms. In contrast, 8‐HCH led to downregulation in D. seriata and E. lata‐inoculated plants, but induced expression in D. eres, highlighting a possible pathogen‐specific modulation. Silver nitrate consistently suppressed VvChit4c expression across all pathogens, potentially indicating an overall suppressive effect on this defense pathway. Interestingly, silver thiosulfate complex and AgSe NPs produced mixed responses depending on the pathogen, further supporting the hypothesis that the grapevine response is highly specific and not solely dependent on the treatment itself. Differential modulation of chitinase gene expression following chitosan treatment and pathogen infection was also confirmed by the study of Martín et al. (2023).57 Application of chitosan to asymptomatic grapevine plants did not result in any significant changes in the expression of chitinase‐encoding genes. In contrast, in plants exhibiting symptoms of Esca disease complex, chitosan treatment led to a pronounced downregulation of these genes. This finding, consistent with the results of our study, suggests that the effect of chitosan is pathogen‐dependent and that, in symptomatic plants, it may modulate defense pathways toward suppression of genes associated with pathogen cell wall degradation.
Finally, the VvSTS gene, responsible for stilbene synthesis, was generally downregulated by most treatments, including 8‐HCH, silver nitrate, silver thiosulfate complex, and AgSe NPs. Stilbenes are known phytoalexins involved in pathogen resistance, and their reduced expression may suggest a possible trade‐off or redirection of defense signaling. However, in E. lata‐inoculated plants, AgSe NPs and sodium arsenite induced VvSTS expression, suggesting a potential enhancement of stilbene‐based defense under certain conditions. According to the study by Songy et al. (2019),42 one of the factors influencing the modulation of VvSTS gene expression may be the phenological stage of the grapevine. Following the application of sodium arsenite to grafted Vitis vinifera cv. Tempranillo plants, a more than twofold upregulation of the VvSTS gene was observed during the flowering stage. In contrast, no significant changes in gene expression were detected during the berry maturity stage. These results suggest that the effect of chemical treatment may be strongly dependent on the developmental stage of the plant.
Together, these results indicate that the tested compounds can differentially modulate grapevine defense‐related gene expression in a pathogen‐dependent manner. While some treatments stimulated specific defense pathways, others appeared to suppress them, underlining the importance of context in evaluating the potential of alternative disease management strategies. Further research should focus on linking these molecular responses to actual disease progression and field efficacy to better understand their protective potential.
CONCLUSION
5
The in planta experiment confirmed the inhibitory activity of selected chemical compounds against GTD pathogens, specifically D. eres, D. seriata, and E. lata. The newly synthesized AgSe NPs exhibited significant inhibitory effects against D. eres and E. lata. However, no significant inhibition was observed against D. seriata. Although AgSe NPs did not demonstrate efficacy against all the tested pathogens, the results suggest that these nanoparticles hold promise as a potential alternative for the plant protection against specific fungal pathogens. Further research is needed to optimize their formulation and broaden their spectrum of activity. The nanoparticles tested in this study showed no signs of phytotoxicity at the concentrations applied, nevertheless, further research using the method of treating inoculated pruning wounds will be necessary.
AUTHOR CONTRIBUTIONS
A. Eichmeier, D. Gramaje and K. Štůsková designed the scheme of the experiment. K. Štůsková established and evaluated the in‐planta experiment, performed the statistical analysis and wrote the main part of the text. T. Kiss and K. Štůsková designed the gene expression analysis procedure, performed real‐time PCR and evaluated the effect of treatments on the expression profile of grapevines. Z. Bytešníková and L. Richtera prepared the NPs used. A. Eichmeier and D. Gramaje approved the scheme of the experiment, supervised the course of the experiment, and edited the text.
Supporting information
Data S1. Supporting Information.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Štůsková K , Pečenka J , Tekielska DA , Špetík M , Bytešníková Z , Švec P et al., The in vitro effects of selected substances and nanomaterials against Diaporthe eres, Diplodia seriata and Eutypa lata . Ann Appl Biol 182:226–237 (2022).
- 2Bertsch C , Ramírez‐Suero M , Magnin‐Robert M , Larignon P , Chong J , Abou‐Mansour E et al., Grapevine trunk diseases: complex and still poorly understood. Plant Pathol 62:243–265 (2013).
- 3Gramaje D , Úrbez‐Torres JR and Sosnowski MR , Managing grapevine trunk diseases with respect to etiology and epidemiology: current strategies and future prospects. Plant Dis 102:12–39 (2018).30673457 10.1094/PDIS-04-17-0512-FE · doi ↗ · pubmed ↗
- 4Mugnai L , Graniti A and Surico G , Esca (black measles) and Brown wood‐streaking: two old and elusive diseases of grapevines. Plant Dis 83:404–418 (1999).30845530 10.1094/PDIS.1999.83.5.404 · doi ↗ · pubmed ↗
- 5Úrbez‐Torres JR , Peduto F , Smith RJ and Gubler WD , Phomopsis dieback: a grapevine trunk disease caused by Phomopsis viticola in California. Plant Dis 97:1571–1579 (2013).30716818 10.1094/PDIS-11-12-1072-RE · doi ↗ · pubmed ↗
- 6Guarnaccia V , Groenewald JZ , Woodhall J , Armengol J , Cinelli T , Eichmeier A et al., Diaporthe diversity and pathogenicity revealed from a broad survey of grapevine diseases in Europe. Persoonia 40:135–153 (2018).30504999 10.3767/persoonia.2018.40.06PMC 6146647 · doi ↗ · pubmed ↗
- 7Guerin‐Dubrana L , Fontaine F and Mugnai L , Grapevine trunk disease in European and Mediterranean vineyards: occurrence, distribution and associated disease‐affecting cultural factors. Phytopathol Mediterr 58:49–71 (2019).
- 8Mondello V , Songy A , Battiston E , Pinto C , Coppin C , Trotel‐Aziz P et al., Grapevine trunk diseases: a review of fifteen years of trials for their control with chemicals and biocontrol agents. Plant Dis 102:1189–1217 (2018).30673583 10.1094/PDIS-08-17-1181-FE · doi ↗ · pubmed ↗