A New Neotropical Species of Gnathostoma (Nematoda: Gnathostomatidae) from the Northern Four-Eyed Opossum Philander vossi (Marsupialia)
M. Á. Mosqueda-Cabrera, E. Sánchez-Miranda, L. D. Castillo-Loeza, G. Torres-Carrera, L. García-Prieto

TL;DR
A new species of Gnathostoma nematode was discovered in a four-eyed opossum in Mexico, distinguished by its morphology and DNA.
Contribution
The discovery and formal description of a new Gnathostoma species from Mexico, supported by morphological and molecular data.
Findings
Gnathostoma mexicanum n. sp. is smaller and has fewer teeth on cuticular spines compared to G. turgidum.
The new species infects the pyloric region, while G. turgidum infects the stomach layers.
Molecular data confirm the genetic divergence of the new species from related Gnathostoma species.
Abstract
This work provides a detailed morphological description of a previously identified but unnamed lineage inside Gnathostoma spp. from Mexico. Specimens of the four-eyed opossum Philader vossi were collected in Tlacotalpan, Veracruz, Mexico. Specimens were examined morphologically using both light microscopy and scanning electron microscopy. Additionally, we generate DNA sequences for the following loci: mitochondrial cox1 and nuclear 5.8S rRNA and ITS-2 and 28S rDNA. The new species, Gnathostoma mexicanum n. sp., differs from G. turgidum Stossich, 1902, the common gnathostomid species infecting Didelphis spp. from the Americas, in its smaller body size, fewer teeth on the cuticular spines at anterior half of body, as well as site of infection (pyloric region vs. stomach layers). Host specificity further distinguishes the two species. Additionally, molecular data show that the new…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
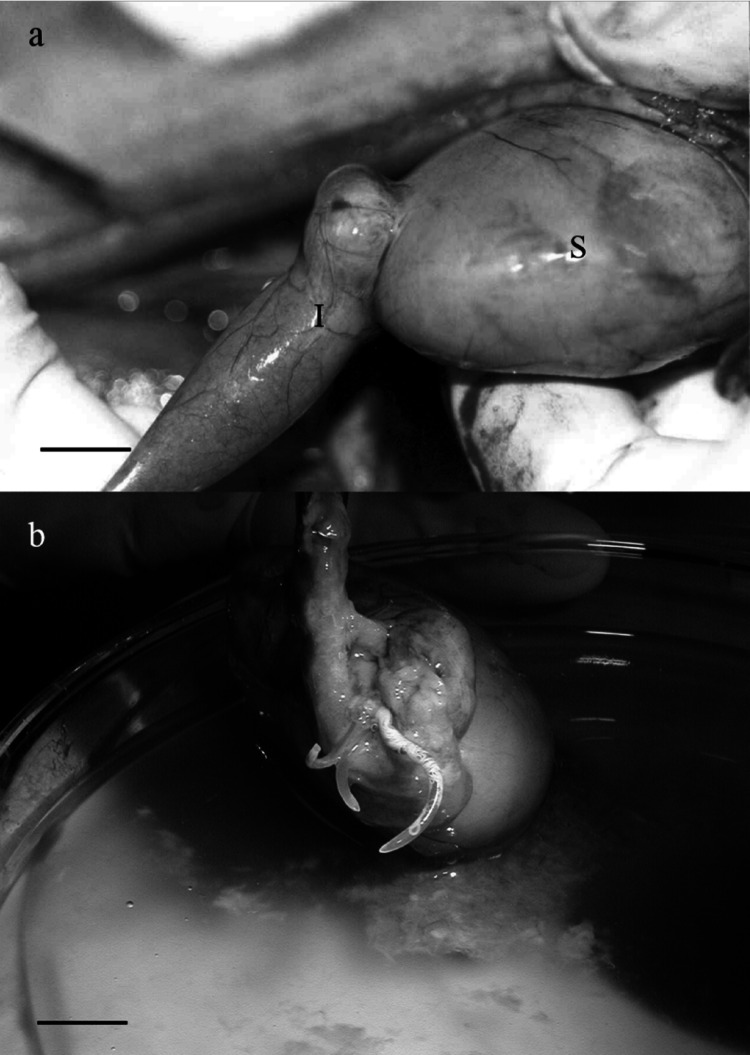 Figure 1
Figure 1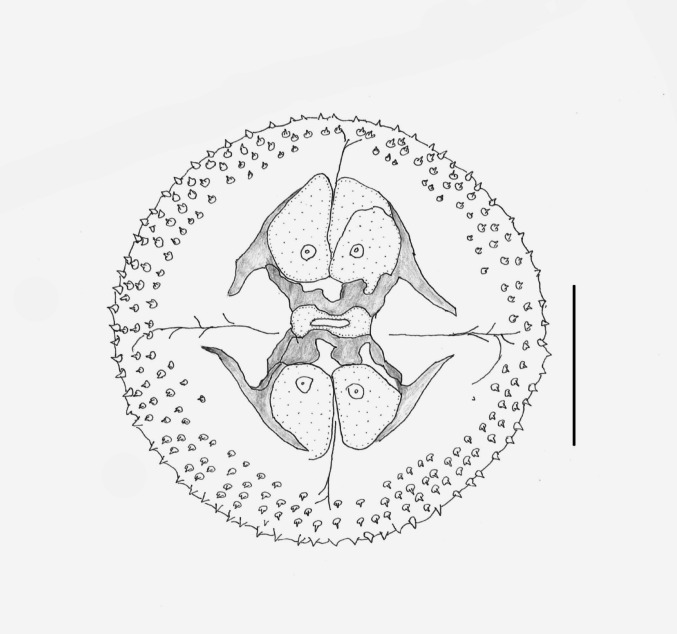 Figure 2
Figure 2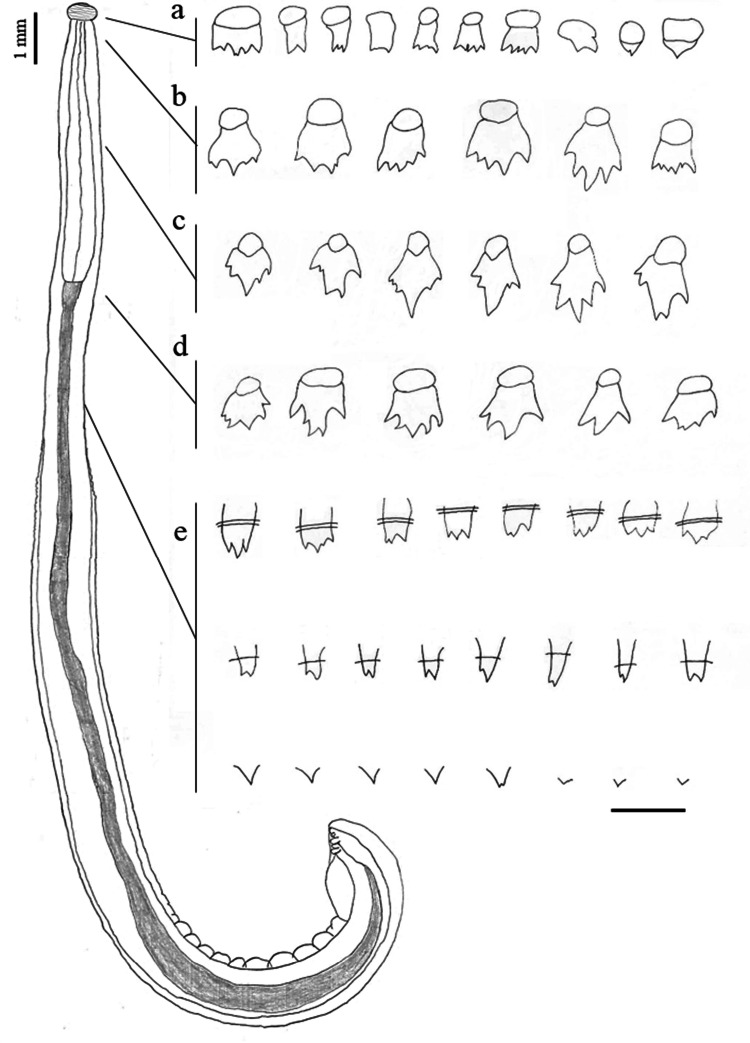 Figure 3
Figure 3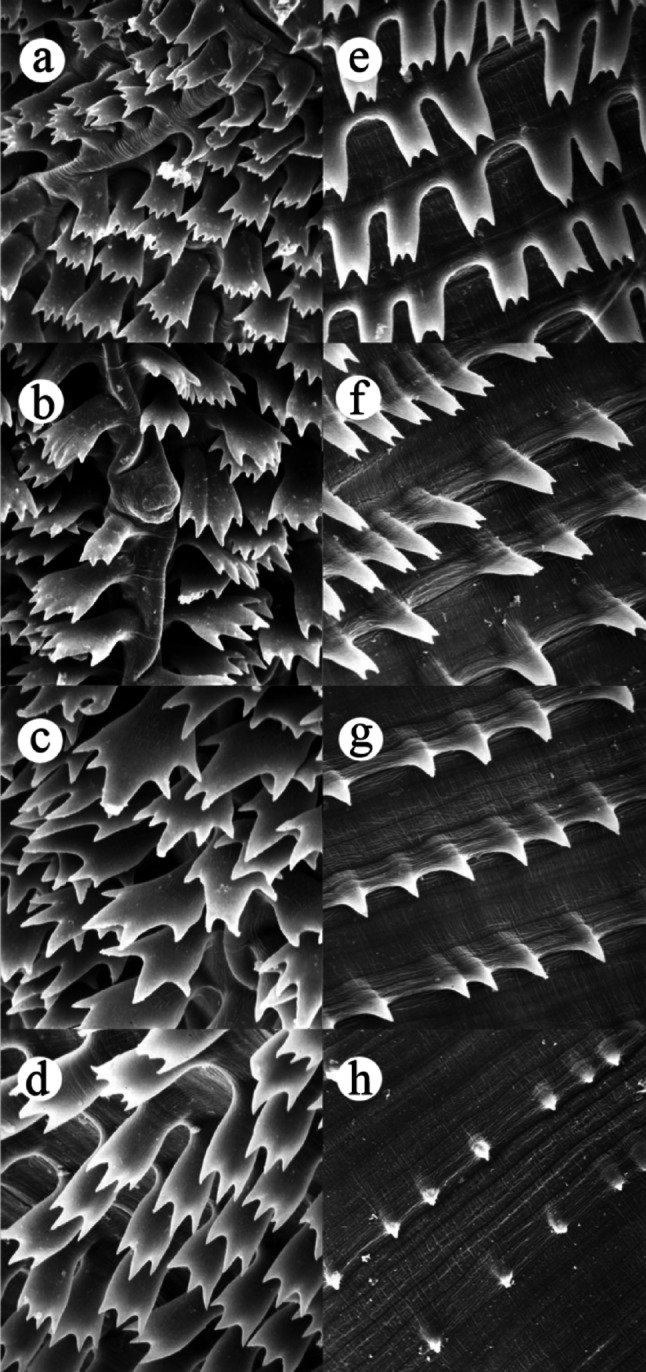 Figure 4
Figure 4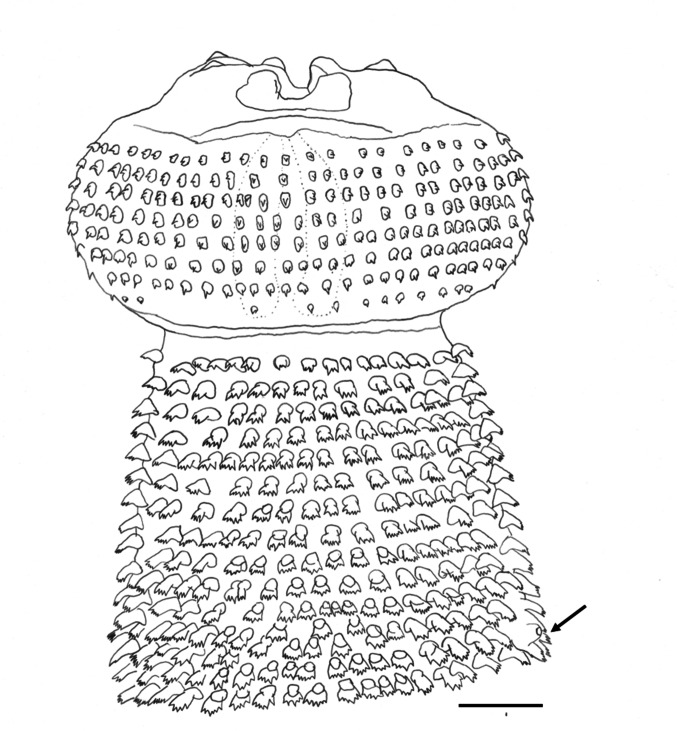 Figure 5
Figure 5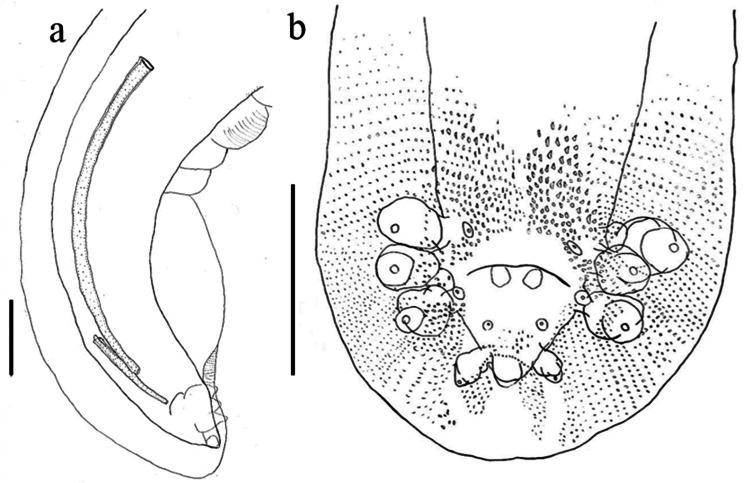 Figure 6
Figure 6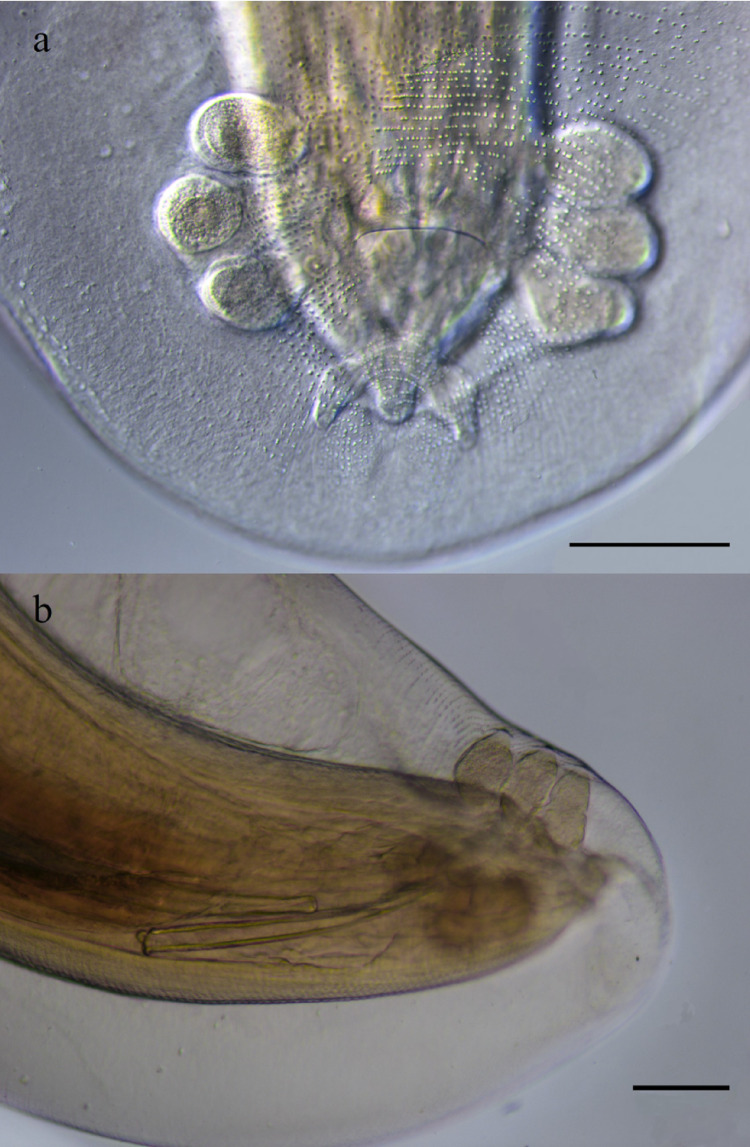 Figure 7
Figure 7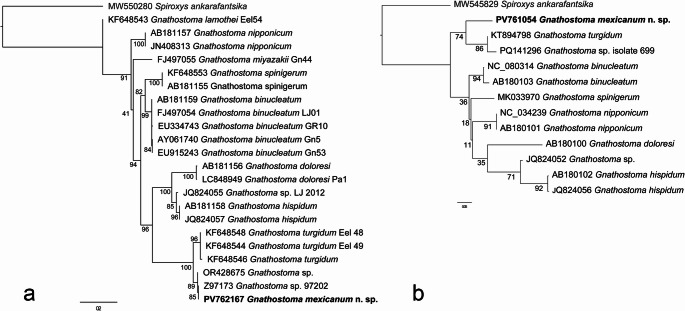 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMollusks and Parasites Studies · Tardigrade Biology and Ecology · Parasite Biology and Host Interactions
Introduction
Nematodes belonging to the genus Gnathostoma Owen, 1837 have been extensively investigated, in part because some species can cause zoonotic disease when third-stage larvae are ingested through undercooked intermediate hosts such as fish or frogs [1, 2]. Naturally, Gnathostoma species infect mammals, primarily carnivores or marsupials, though they have also been recorded in pigs [3, 4].
Currently, there are 13 valid species of Gnathostoma, seven of which ranging in the American continent [5]: Gnathostoma sociale Leidy, 1858 from Mustela vison of USA [6]; Gnathostoma procyonis Chandler, 1942 infecting Procyon lotor of south USA [7]; Gnathostoma lamothei Bertoni-Ruiz et al. [3] (= Gnathostoma sp. I sensu [8], and = *G. procyonis *sensu [9]) in P. lotor hernandezii of Mexico); Gnathostoma binucleatum Almeyda-Artigas, 1991 in Leopardus pardalis and Felis catus from Mexico and Ecuador [8–11]; Gnathostoma americanum Travassos, 1925, a parasite of Leopardus tigrinus from Brazil; and Gnathostoma turgidum Stossich, 1902 (= G. didelphis Chandler, 1932), [5, 8, 12]. All of these species parasitize the stomach of their host, except for Gnathostoma miyazakii Anderson, 1964, which was found in the kidney tissue of Lontra canadensis in Canada and the USA [13, 14].
Following a detailed morphological and molecular study of Gnathostoma specimens collected in Mexico in the Northern four-eyed opossum, Philander vossi, we identified an undescribed species of this genus. This work aims to describe this new species, differentiating it from the seven known American Gnathostoma species, based on morphological and molecular evidence.
Materials and Methods
Specimen Collection
Eight specimens of P. vossi collected between April 2022 and March 2023 under collection permit SCPA/DGVS/03184/22 issued by the Secretaría de Medio Ambiente y Recursos Naturales, Mexico, were necropsied for helminths. Briefly, Tomahawk traps baited with a mixture of tuna and canned bananas were placed near freshwater bodies around Tlacotalpan city (18° 38′ 07.06″ N, 95°38′ 40.68″ W), Veracruz, Mexico. The marsupials were immediately taken to the laboratory to be sedated through premedication with xylazine hydrochloride (0.2 mg/kg), induction with ketamine (0.5 mg/kg), and euthanized with intraperitoneal overdose of sodium pentobarbital. All marsupial specimens were subjected to necropsy.
Morphological Analyses
The majority of collected worms were rinsed in saline (NaCl 0.85%), relaxed in chloral hydrate, fixed in hot 70% ethanol, stored in 70% ethanol, and examined in temporary whole mounts after clearing in Amman’s lactophenol. Measurements are given in millimeters unless specified otherwise and presented individually for holotype, allotype, and 2 paratypes (adult male and juvenile female). Line drawing was made using a drawing tube mounted on a stereoscope. Type specimens were deposited in the Colección Nacional de Helmintos (CNHE) Instituto de Biología (IB), Universidad Nacional Autónoma de México (IBUNAM), Mexico City, Mexico. For comparative purposes, the following Gnathostoma species from the United Sate National Museum (USNM), Smithsonian Institution, Washington, D.C, USA were examined: G. oligomucronatum nomen nudum (accession numbers: 88146, 88147 and 88148 deposited by Javier Almeyda Artigas); and G. turgidum from D. virginiana from CNHE, accession numbers: 4736, 4261, 4740, 4261.
One specimen (female), used for scanning electronic microscopy (SEM) studies, was processed according to [15] i.e., the specimen was fixed in 5% alkaline paraformaldehyde, and postfixed in osmium tetroxide; subsequently dehydrated with graded alcohols to acetone, dried at critical point (CO_2_), coated with a mix of gold–palladium, mounted on a metal stub and examined using a Hitachi SU1510 (Hitachi, Tokyo, Japan) scanning electron microscope at 10 kV at the Laboratorio Nacional de Biodiversidad (LANABIO), IB-UNAM. Ecological parameters follow [16].
Molecular and Phylogenetic Analysis
Specimens for molecular analysis were preserved in 100% ethanol. DNA extraction was performed using the DNeasy Blood & Tissue (QUIAGEN) kit following the manufacturer’s protocol. A fragment of internal spacers transcribed ITS-1, complete 5.8S rDNA gene and a fragment of ITS-2 (hereinafter referred to as ITS for practical purposes), with the ITS4/ITS5 primers [17], with the following thermal profile: 1 cycle of 94 °C for 5 min; 35 cycles of 94 °C for 1 min, 52 °C for 1 min and 72 °C for 1 min; final elongation at 72 °C for 10 min. We also amplified partial Cytochrome C oxidase (cox1) and partial sequence of large ribosomal subunit 28S rDNA, with the following primers LCO/HCO [18] and 28SA/28SBout [19, 20], respectively, with the same thermal profile used for ITS, only interchanging to 48 °C annealing temperature for cox1 amplification.
PCR products were visualized by electrophoresis on agarose gel. Successful amplifications were purified using CentriSep 96 filter plates (ThermoFisher Scientific, Pittsburgh, Pennsylvania) with Sephadex G-50 (Cytiva, Marlborough, Massachusetts). Sequencing reactions included 0.4 µl BigDye Terminator v. 3.1 (Applied Biosystems, Waltham, Massachusetts), 2 µl Buffer 5x, 4 µl ddH2O, 1 µl of primer at 10 µM, and 3 µl purified PCR product (total volume 10 µl. Samples were purified using Sephadex G-50, then 25 µl de EDTA 0.5 mM was added to each sample and finally sequenced in an ABI-PRISM 3100 (Applied Biosystems® Waltham, Massachusetts) LANABIO, IB-UNAM.
The sequences generated here were aligned in MAFFT online version [21] with the sequences available in GenBank for cox1 and ITS independently. Spiroxys ankarafantsika, a member of Gnathostomatidae, was selected as outgroup. Phylogenetic relationships were constructed based on ITS sequences (644 bp and 26 terminals) as well as based on a cox1 matrix (1097 bp and 13 terminals); for more details of sequences used see Table 1. For the 28S rDNA region, a Blast search did not return any matches with Gnathostoma spp. due to the scarcity of available sequences for this genus in databases. Furthermore, the few sequences are notably short (e.g., G. spinigerum with 190 bp; acc. no. MT151891) making a reliable phylogenetic inference based on this region unfeasible. The inference criteria used was Maximum likelihood performed in IQtree [22] establishing 1000 pseudo replicates of ultrafast Bootstrap for nodal support value [23] and estimating automatically the substitution model that best fit to the data [24]. The best fit model for cox1 was TIM3 + F + I and for ITS was HKY + F + G4. Genetic distances were calculated with Mesquite [25] using the Kimura 2-parameters model according to [26].Table 1. Metadata associated to taxa used in phylogenetic analysis of Gnathostoma speciesTaxon namesLocalitiesAdult formLarvaCox1 GenBank acc noITS GenBank acc noReferencesGnathostoma binucleatumMexicoUnidentified fishNC_080314Díaz-Camacho et al. UnpublishedNayarit, MexicoHomo sapiensAY061740,[27]Colima, MexicoSciades guatemalensisFJ497054[28]Sinaloa, MexicoDormitator latifronsEU915243[29]Guayaquil, EcuadorRhamdia cinerascensAB180103AB181159[30]Gnathostoma doloresiJapanSus scrofaAB180100AB181156[30]Unidentified eelLC848949[31]Gnathostoma miyazakiiCanadaLontra canadensisFJ497055[28]Gnathostoma hispidumUnidentified eelJQ824056JQ824057Lee et al. UnpublishedChinaSus scrofaAB180102AB181158[30]Gnathostoma lamotheiVeracruz, MexicoProcyon lotor hernandeziiAY818004[3]Tabasco, MexicoGobiomorus dormitorEU334736[32]EUAMonopterus sp.KF648543[33]Gnathostoma mexicanum n. sp.“El Jobo” lagoon margins, Veracruz, MexicoPhilander vossiPV761054PV762167This WorkPhilander vossiOR428675[34]Gnathostoma nipponicumHeilongjiang, China,Unidentified fishJN408313Lee et al. UnpublishedJapanMustela sp.NC_034239Sun et al. UnpublishedMie, JapanMustela sibirica itatsiAB180101AB181157[30]G. spinigerumUSAMonopterus sp.KF648553[33]CambodiaMonopterus albusMK033970[35]Nakhon Nayok, ThailandMonopterus albusAB181155[30]Gnathostoma sp.IndonesiaUnidentified eelJQ824052JQ824055Lee et al. UnpublishedGnathostoma sp. IITlacotalpan, Veracruz, MexicoPhilander vossiZ97173[8]Gnathostoma turgidumUSAMonopterus sp.KF648544, KF648546, KF648548[33]BrazilDidelphis auritaKT894798[36]Gnathostoma sp. 1Oaxaca, MéxicoDormitator latifronsPQ141296[34]Spiroxys ankarafantsikaSouth AfricaPelusios castanoidesMW545829MW550280[37]
Results
Gnathostomatidae Railliet, 1895.
Gnathostomatinae Railliet, 1895.
Gnathostoma Owen, 1836.
Gnathostoma mexicanum** n. sp.** (Figs. 1, 2, 3, 4, 5, 6 and 7). Fig. 1. Infection site of Gnathostoma mexicanum n. sp. found in Philander vossi. a Tumor between stomach (S) and intestine (I), b Adult worms in the tumor. Scale bars, a = 20 mm, b = 5 mm
Taxonomic Summary
Type host: Philander vossi, Northern four-eyed opossum (Didelphimorphia: Didelphidae: Didelphinae).
Site of infection: in a single pyloric tumor, in which only the anterior third is buried; body in the lumen of the small intestine (Fig. 1).
Type locality: “El Jobo” lagoon margins (18° 37′ 58″ N, 95° 39′ 29″ W), proximities of Tlacotalpan City, Veracruz State, Mexico.
Specimens deposited: Holotype male CNHE: 12,849, allotype female CNHE: 12,850, Paratype male USNPC: 88,146, paratype female USNPC: 88,147, and paratype juvenile female CNHE: 12,287.
Representative DNA sequences, GenBank accession numbers: cox1: PV761054, ITS: PV762167 and 28S rDNA PV762168.
Etymology: This species is named after the country from which the parasite was collected.
Prevalence: 50.0% (8 hosts collected, 4 infected).
Mean intensity: 0.85 (4 infected host/5 worms).
ZooBank registration: urn:lsid:zoobank.org:pub:83C98784-3567-4E81-AD89-2DC27570AE30.
Description based on 5 adult specimens, complete sets of measurements from the male holotype, the female allotype, 2 paratypes (male and female), and one juvenile paratype (female).
Shared structures somewhat larger in females than in males. Delicate and slender worms; anterior fourth of body slightly thinner than posterior three fourths. Cephalic bulb armed with 8–9 rows of uncinated hooklets and one pair of broad protruded lips with four small, submedian, double papillae, and lateral amphids not seen (Fig. 2). Ballonets 4 in number. Esophagus divided into short, anterior, muscular part and long, posterior, glandular part. The anterior half of body covered with transverse rows of cuticular spines of several shapes and different number of points (Figs. 3 and 4), as well as a pair of lateral cervical papillae between rows 13–16 (Fig. 5).Fig. 2. Apical view of the cephalic bulb of female of Gnathostoma mexicanum n. sp. collected in Philander vossi. Scale bar, 200 µmFig. 3Morphology of body spines of male of Gnathostoma mexicanum n. sp. collected in Philander vossi. a first rows of body, b at level of the cervical papillae, c middle region of the esophagus, d esophagus-intestine junction, e subsequent levels of the spinated surface. Scale bar, 50 µmFig. 4Scanning electron microscopy of adult (male) of Gnathostoma mexicanum n. sp. collected in Philander vossi a at neck level, b at cervical papilla level, c at the mid-oesophageal level, d at oesophagus-intestinal junction level, e–h subsequent levels of the spine surfaceFig. 5Anterior end of the male Gnathostoma mexicanum n. sp., collected from Philander vossi, showing arrangement of cuticular spines in cephalic bulb up to level of the cervical papilla (arrow). Scale bar = 100 µm
Holotype: total length 27.99; body width at anterior fourth 0.740; maximum body width at posterior half 1.170. Esophagus length 5.190 (18.540% respect to the total length of body), maximum width 0.510; four cervical sacs, 1.480 length (28.5%, respect to the total length of esophagus). Two cervical papillae between the transverse rows of cuticular spines, 14 (right) and 15 (left); excretory pore in row 33. Cephalic bulb 0.240 length by 0.540 width armed with eight complete transverse rows of hooks. Cephalic hooks 12–20 μm (15 ± 0.3, n = 10) long, 7–12 μm (9 ± 2, n = 10) wide at base; hooks of the last row smaller 5–7 μm (6 ± 1, n = 10) long, 5–5 μm (5 ± 0, n = 10) wide at base. Lips 29 μm long by 74 μm wide.
Body spines arrangement as follows: first rows posterior to neck multidentate (2 to 4 teeth, commonly 3) [lateral teeth thicker and slightly longer than central, 10–27 μm (19 ± 5, n = 10) long by 10–17 μm (13 ± 3, n = 10) wide in the region of the lateral teeth]; spines at cervical papillae level (4 to 6 points, often 5,) central teeth slightly longer than lateral, 29–39 μm (35 ± 3, n = 10) long by 17–27 μm (23 ± 4, n = 10) wide at region of the lateral teeth; large spines at mid-esophageal region, multidentate (4 to 6 teeth, generally 5; central tooth longer and thicker, occasionally forked), 54–61 μm (58 ± 2, n = 10) long by 39–49 μm (46 ± 3, n = 10) wide at lateral teeth level (Figs. 3 and 4). Spinous area extension 12.515 (44.70%). At the esophagous-intestinal junction, multi-tipped cuticular teeth (3–5 teeth, commonly 5) [if 3, the central teeth are longer and thicker than the lateral ones; frequently central teeth with 1 or 2 lateral teeth, on each side; some spines with the first pair of teeth slightly shorter than the 3 central ones; 61–69 μm (66 ± 3, n = 10) long, 29–42 μm (34 ± 4, n = 10) wide at the level of the lateral teeth]; the remaining spinous surface with the following subdivisions: (a) at the end of first third, with 2 and 3 equal teeth, sometimes central shorter; (b) at the middle of the second third, with 2 teeth of equal/unequal size and, (c) at the end of the spinous surface with single-tooth spine (Figs. 3 and 4).
Posterior half of body smooth, except in anal region, where transverse rows of single-tooth spine directed forward are observed. Caudal extremity with broad alae (Fig. 6) and eight pairs of caudal papillae, arranged as follows: four pairs of large, pedunculate, lateral papillae, one pair (1) precloacal, one pair (2) adcloacal and two (3, 4) postcloacal; the posterior pair much shorter than the others; the first three pairs lying close together, the fourth markedly separated from them, very close to the tip (Figs. 6 and 7). Four pairs of small, sessile, ventral papillae: one pair (A) preacloacal, one pair (B) adcloacal, and two (C, D) postcloacal (Figs. 3 and 6B). Tail 0.27 in length. Spicules delicate, unequal, with rounded apices; right spicule 0.64, much shorter than left (2.29); ratio between spicules 1.00:3.58 (Figs. 6 and 7).Fig. 6. Posterior end of male of Gnathostoma mexicanum n. sp., parasite of Philander vossi. a Right lateral view and spicules, b Caudal bursa spination and genital papillae pattern. Scale bars = 100 µmFig. 7Caudal bursa of Gnathostoma mexicanum n. sp. parasite of Philander vossi. a Ventral view, b Right side view. Scale bar = 200 µm
Allotype: Total length: 36.29; body width: 8.4 in the forequarters; maximum width in the posterior half: 14. Esophagus length: 42 (11.61% of the total length) by 0.60 maximum width. Four cervical sacs, 1.52 (36.0%, respect to esophageal length). Cervical papillae between transverse rows 15 (rigth) and 16 (left) of cuticular spines. Excretory pore in row 37. Cephalic bulb 0.29 length by 0.55 width armed with nine complete transverse rows of hooklets. Body spines similar to those in male in the number of teeth, but with greater density in all regions. Extension of spinous area 47.60%. Vulva slightly postequatorial at 20.29 from anterior end (55.92%). Tail 0.21 in length. Embryonated eggs colourless. Eggshell with fine pits on entire surface and provided with cap-like thickenings in both poles. Eggs 62–70 μm (67 ± 5, n = 30) × 35–40 μm (38 ± 5, n = 30).
Paratype (male): total length 23.04; body width at anterior fourth 0.71; maximum body width at posterior half 1.10; cervical papillae between transverse rows 14 and 15 of cuticular scales; cephalic bulb 0.29 length, 0.48 width, armed with eight complete and one incomplete transverse rows of hooklets; esophagus length 2.69, with a maximum width of 0.47; extension of spined area 55.08% of body length. Body spines arrangement similar in the number of teeth to that in holotype, but with lower density in all regions respect the allotype. Posterior half of body smooth, except in anal region, where transverse rows of one-tooth spines directed anteriorly are present. Caudal extremity with broad alae and eight pairs of caudal papillae, arranged as follows: four pairs of large, pedunculate, lateral papillae; one pair (1) precloacal, one pair (2) adcloacal and two (3, 4) postcloacal; the posterior pair much shorter than the others; the first three pairs are very close to each other, the fourth markedly separated from them, very close to the tip. Four pairs of small, sessile, ventral papillae: one pair (A) precloacal, one pair (B) adcloacal, and two (C, D) postcloacal. Tail 0.28 in length. Spicules delicate, uneven, with blunt apices; right 0.64, shorter than left (2.20); ratio 1.00:3.44.
Paratype (female): Total length 38.84; body width at anterior fourth 0.97; maximum width at posterior half 1.71. Cervical papillae between transverse rows 14 and 15 of cuticular scales. Cephalic bulb length 0.35 and 0.61 width, armed with eight complete and one incomplete transverse row of hooklets; spinous area occupies 49.51% of the body length. Body spines arrangement similar to those in allotype, with an apparently higher density respect the holotype. Vulva postequatorial, situated at 20.38 from anterior end (52.47%); fertilized uterine eggs colorless, ovum unsegmented. Eggshell with fine granulations on entire surface, provided with cap-like thickenings in both poles. Eggs 66.3 μm (60.4–70.0, n = 10) × 36.8 μm (34.0–40.0, n = 10).
Juvenil (female): Body length: 15.93 by 0.57 maximum width. Two cervical papillae on rows 13 (left) and 16 (right). The spiny surface of the body covers 8.73 of its length, reaching the level of the postequatorial vulva (54.7% of body length). Cervical sacs: 1.58 length on average. Esophagus: 2.53 (15.8% of body length) by 0.36 width. Body width at the esophagus-intestinal junction: 0.47. Head bulb: 0.20 × 0.34 length and width, respectively, with eight complete rows and one incomplete row of single-pronged hooks. Tail length: 0.24. Body spines similar to those of the allotype in the number of teeth, but with lower density in all regions.
Molecular and Phylogenetic Results
Both phylogenetic trees recover the new species as a sister taxon to G. turgidum with high support value (100%) in the ITS-based tree, but regular/low (74%) in the cox1-based tree. In general, internal relationships are not consistent between both trees, showing distinct topologies in early divergent branches (Fig. 8).Fig. 8. Maximum likelihood reconstruction of phylogenetic relationships between Gnathostoma species based on ITS (a) and cox1 (b) DNA sequences. The new species are denoted in bold
Comparative genetic divergences of ITS locus shows that the new species is nearly identical to unidentified larva Gnathostoma sp. II found parasitising P. vossi from Tlacotalpan, Veracruz (Table 2) and differs from the G. turgidum sequences by 6.4% on average, while intraspecific divergences do not exceed 0.5% in G. turgidum and 0.2% in the new species. Genetic divergence of cox1 between G. mexicanum and G. turgidum was 12.3% (Table 3).Table 2. Pairwise genetic distances (K2P) of ITS region between Gnathostoma speciesGnathostoma species(1)(2)(3)(4)(5)(6)(7)(8)(9)(10)(1) G. turgidum0.5%(2) Gnathostoma mexicanum n. sp.6.4%0.2%(3) Gnathostoma sp. II7.4%0.4%NA(4) G. hispidum30.6%31.1%51.5%1.7%(5) G. doloresi36.5%37.4%57.8%16.3%0.6%(6) G. binucleatum32.6%32.6%40.8%23.1%31.7%0.1%(7) G. spinigerum33.4%33.8%47.4%24.7%30.8%12.9%0%(8) G. nipponicum32.6%33.5%45.4%24.1%30.7%15.9%19.4%0%(9) G. miyazakii38.1%36.4%45.3%29%35%18%23.6%17.1%NA(10) G. lamothei28.3%32.2%43.6%27%34.3%20.8%24.1%19.5%19.7%0%Table 3. Pairwise genetic distances (K2P) of cox1 region between Gnathostoma speciesGnathostoma species12345678(1) G. binucleatum1.8%(2) G. nipponicum11.3%0%(3) G. spinigerum17.1%13.4%NA(4) G. doloresi14.9%13.8%NANA(5) G. hispidum12.3%15.1%NA13.8%0.5%(6) Gnathostoma sp.19.0%14.6%NA13.1%7.5%NA(7) G. turgidum24.3%15.5%15.4%NANANA3.1%(8) G. mexicanum n. sp.15.0%15.3%16.4%NANANA12.9%NA
Remarks based on host specificity observed in the adult Gnathostoma spp., as well as the distribution, density and kind/shape of transversal rows of cuticular spines, the new species is readily distinguishable from the 13 valid species comprising the genus (Table 4). Comparison of the redescriptions by [5] of the American Gnathostoma species with both the morphological observations of our specimens and phylogenetic results indicates that the new species most closely resembles G. turgidum.Table 4. Distinctive characters between species of the genus Gnathostoma and Gnathostoma mexicanum n. spTaxonCharacterReferencesType hostCountryLength (mm) (Male, Female)Percentage of body surface covered by spinesMaximum number of teeth on the spinesPolar plugs in the eggGnathostoma spinigerumOwen (1836)Panthera tigris^1^Englad(12–31, 11–34)Entire4–51[38, 39]Gnathostoma socialeLeidy (1858)Mustela vison^1^USA(24, 30–35)345–61[6, 40]Gnathostoma hispidumFedtschenko (1872)Sus scrofa ferus, S. s. domesticus^1^Hungary(20, 26)Entire7–101[38, 41]Gnathostoma turgidumStossich (1902)Didelphis azarae^1^Argentina(40–42, 67–72), females up to 100 in length505–10^^2[5, 42, 43]Gnathostoma doloresiTubangui (1925)Sus scrofa domesticus^1^Philippines(11–12, 30–41)Entire6–72[38, 44]Gnathostoma americanumTravassos (1925)Leopardus tigrinus^1^Brazil?755–72[42]Gnathostoma nipponicumYamaguti (1941)Mustela sibirica itatsi^2^Japan(20–28, 22–34)504–71[38, 45]Gnathostoma procyonisChandler (1942)Procyon lotor lotor^1^USA(16–19, 20–26)Entire3–41[46, 47]Gnathostoma miyazakiiAnderson (1964)Lontra cannadensis^3^Canada(42, 41)Bosses in the posterior half, only in males6–71[6]Gnathostoma malaysiaeMiyazaki and Dunn (1965)Rattus surifer^1^Tioman Island(16, 22)Entire62[48]Gnathostoma vietnamicumLe-Van-Hoa (1965)Lutrogale perspicillata^3^Vietnam(28, 43)3391[49]Gnathostoma binucleatumAlmeyda-Artigas (1991)Leopardus pardalis^1^Mexico(21–25, 15–27)Entire3^^1[50]Gnathostoma lamotheiBertoni-Ruiz, García-Prieto, Osorio-Sarabia and León-Règagnon (2005)Procyon l. hermandezii^1^Mexico(13–19; 18–20)Bosses in the posterior half4–6^^1[3]Gnathostoma mexicanum n. sp.Philander vossi^4^Mexico(23, 39)45–484–6^^2Present study1 = Stomach, 2 = Esophagous, 3 = Kidney, 4 = Pylorus, *at level of cervical papillae
The main similarities are: (a) number of hooks from cephalic bulb; (b) spines present only at the anterior mid of body; (c) number and arrangement of caudal papillae; (d) position of the vulva, and (e) cap-like thickness at each end of the eggs. However, remarkable differences were also found: (1) fewer teeth on spines at level of cervical papillae (six vs. 10 teeth in G. turgidum); (2) spines from mid-esophagous level to intestine junction with maximum two lateral small teeth vs. maximum four lateral small teeth in G. turgidum; (3) minor body size (up to three times smaller than the Stossich’s species); (4) larger egg size (66.3 µm in the new species vs. 51.0 µm); (5) egg surface with fewer pits in G. mexicanum and (6) smaller cap-like length (3.48 µm vs. 4.35 µm in G. turgidum).
In addition, the new species produces tumors at the pyloric region of host with the anterior third of body embedded in the layers of the serosa. Gnathostoma turgidum forms individual tunnels penetrating the anterior part of body inside the mucus and muscularis of stomach, without specificity for some region. Larvae of four-stage and juvenils of G. turgidum have been reported infecting the liver of P. opossum [51, 52], which indicate that these larval stage nematode migrate through liver to reach the stomach where this mature and reproduce [53]. The finding of juvenile stages of G. mexicanum sp. n. exclusively at pyloric tumor, suggests that migration not encompass the liver. Notwithstanding, this require future investigations.
Discussion
After the examination of the specimens labeled as Gnathostoma oligomucronatum (= Gnathostoma sp. II sensu [8]) by Bertoni-Ruiz et al. [5], a parasite of the pyloric region of Philander vossi (= Philander opossum pallidum) from El Jobo Lagoon, Tlacotalpan, Veracruz, Mexico, and deposited at USNM (88148, and 58535.02 by Javier Almeyda Artigas), the authors concluded that they correspond to G. turgidum. However, Almeyda-Artigas et al. [8] reported that ITS-2 rDNA sequences from this material support the distinction of Gnathostoma sp. II from G. turgidum. Unsurprisingly, the percentage of identity of the larva of Gnathostoma sp. II from Veracruz matches with G. mexicanum.
To our knowledge, no formal description of G. oligomucronatum has been published. Nevertheless, morphometric data from this material (USNM 88146, 88147, and 88148) align precisely with the diagnosis of the new species described herein. Consequently, G. oligomucronatum is relegated to nomen nudum.
With the species described here, there are now eight recognized Gnathostoma species in the Americas.