16S rRNA gene amplicon sequences from coastal sediments at the Sonoran Desert and Gulf of California interface
Cristal Ramos-Madrigal, Esperanza Martínez-Romero, Yunuen Tapia-Torres, Luis E. Servín-Garcidueñas

TL;DR
This paper presents the bacterial and archaeal community composition in coastal desert sediments using 16S rRNA gene sequencing.
Contribution
The study provides a detailed taxonomic analysis of microbial communities in a unique extreme environment at the Sonoran Desert and Gulf of California interface.
Findings
Halobacterota, Bacteroidota, and Proteobacteria were identified as dominant phyla in the sediments.
The 16S rRNA gene amplicon data serves as a resource for studying microbial diversity in extreme environments.
Abstract
We report the taxonomic composition of bacterial and archaeal communities from sand and salt-crust sediments in the coastal Sonoran Desert, Mexico. Analysis of 16S rRNA gene amplicon sequences revealed Halobacterota, Bacteroidota, and Proteobacteria as dominant phyla, providing a resource to explore microbial diversity in extreme environments.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
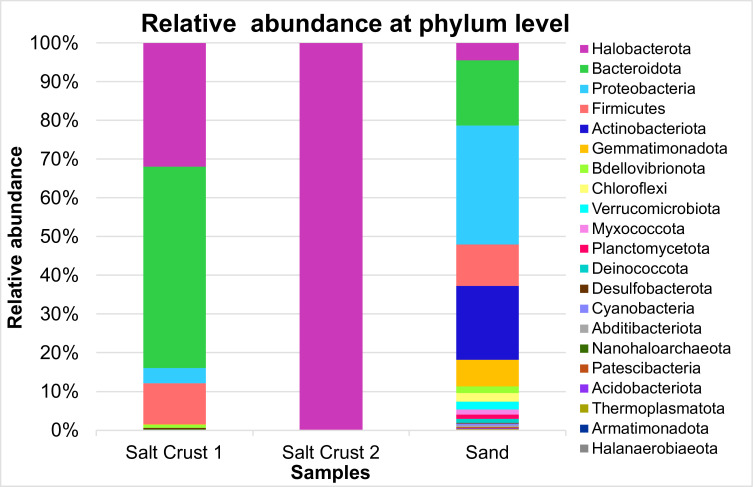 Fig 1
Fig 1| Primers | Sample | Relative abundance of Bacteria (%) | Relative abundance of Archaea (%) | Total paired-end reads | Reads after quality filtering (Q20) | Reads after denoising and chimera removal | No. of ASVs | Identified genera |
|---|---|---|---|---|---|---|---|---|
| 341F | Salt Crust 1 | 67.74 | 32.25 | 181,801 | 135,455 | 52,202 | 271 | 36 |
| Sand | 95.32 | 4.67 | 91,715 | 48,328 | 33,797 | 789 | 125 | |
| 787F | Salt Crust 2 | 0.07 | 99.92 | 87,922 | 66,228 | 20,883 | 110 | 17 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Community Ecology and Physiology · Genomics and Phylogenetic Studies · Methane Hydrates and Related Phenomena
ANNOUNCEMENT
In the Sonoran Desert along the Gulf of California, extensive salt crusts and sediments form under hypersaline conditions, temperature extremes, and intense solar irradiance. Here, we profiled bacterial and archaeal diversity in these coastal habitats using 16S rRNA amplicon sequencing to provide a high-resolution taxonomic profile of desert sediments.
Samples of salt crust and surface sand were collected in March 2024 at a coastal site (31°42′36″N, 113°58′07″W). Samples were stored in sterile bags at room temperature until DNA extraction. DNA was extracted with the DNeasy PowerSoil Pro kit (Qiagen, Germantown, MD, USA), following the manufacturer’s instructions. DNA was quantified with the QuantiFluor dsDNA System (Promega, Madison, WI, USA). The salt-crust and sand samples were analyzed with two primer sets targeting different 16S rRNA regions. The V3-V4 region was amplified using the primers 341F (5′-CCTACGGGNGGCWGCAG-3′) and 805R (5′-GACTACHVGGGTATCTAATCC-3′), hereafter “Salt Crust 1” and “Sand” (1). The V5-V6 region was amplified using the archaeal-biased primers 787F (5′-GGATTAGATACCCSBGTAGT-3′) and 1059R (5′-ACGGGCGGTGTGTACAAG-3′), hereafter “Salt Crust 2” (2). The sand sample amplified only with primers 341F/805R. PCR was performed with an initial denaturation at 95°C for 3 min; 25 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 30 s; a final extension at 72°C for 5 min. Libraries were prepared with the Nextera XT Index Kit (Illumina), following the manufacturer’s instructions. Libraries were sequenced on an Illumina MiSeq (2 × 301 bp; Macrogen Co., Seoul, South Korea). Read quality was assessed with FastQC v0.11.8 (with default settings) (3) and trimmed at Q20 using Trimmomatic v0.39 (4). Reads were further processed in QIIME 2 v2024.10 (5, 6) with q2-dada2 (7) for denoising and chimera removal. Taxonomy was assigned to amplicon sequence variants (ASVs) using the q2-feature-classifier (classify-consensus-vsearch method) (8) with the SILVA v138 SSURef Nr99 database (9).
Sand showed the highest number of unique ASVs, whereas salt crust yielded fewer but more specialized lineages, consistent with hypersaline conditions and primer specificity (Table 1). In the sand sample, the dominant phyla were Proteobacteria (30.70%), Actinobacteriota (19.08%), and Bacteroidota (16.83%). In Salt Crust 1, the most abundant phyla were Bacteroidota 51.95%, Halobacterota 31.96%, and Firmicutes 10.56%. In Salt Crust 2, 99.92% of sequences belonged to Halobacterota, whereas 0.07% corresponded to Actinobacteriota (Fig. 1). Thus, different primer sets revealed complementary diversity. Salt Crust 1 and Salt Crust 2 shared 13 archaeal genera; they shared seven and five genera, respectively, with the Sand sample. Two genera were exclusive to Salt Crust 1, and four were exclusive to Salt Crust 2, indicating complementary detection of archaeal groups.
Relative abundance of bacterial and archaeal phyla based on 16S rRNA amplicon sequencing. Sand and Salt Crust 1 were amplified with 341F/805R primers for the detection of Bacteria and Archaea. Salt crust 2 used archaeal-biased 787F/1059R primers to further investigate archaeal diversity.
Halophiles, including Natronorubrum tibetense (identified in all samples), Natronococcus occultus (Salt Crusts 1 and 2), Halalkalicoccus jeotgali (Salt Crust 2), Natrinema pellirubrum (Salt Crust 1), and marine halophilic bacteria of the order Oceanospirillales (Salt Crust 1 and Sand), have been reported in the Sonoran Desert and other hypersaline environments (10–16). Halophiles have biotechnological potential (17–19). These findings highlight the coastal zone of the Sonoran Desert as a microbial reservoir and a priority for further investigation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Herlemann DP, Labrenz M, Jürgens K, Bertilsson S, Waniek JJ, Andersson AF. 2011. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J 5:1571–1579. doi:10.1038/ismej.2011.4121472016 PMC 3176514 · doi ↗ · pubmed ↗
- 2Yu Y, Lee C, Kim J, Hwang S. 2005. Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction. Biotechnol Bioeng 89:670–679. doi:10.1002/bit.2034715696537 · doi ↗ · pubmed ↗
- 3Andrews S. 2010. Fast QC: a quality control tool for high throughput sequence data. Available from: https://www.bioinformatics.babraham.ac.uk/projects/fastqc
- 4Bolger AM, Lohse M, Usadel B. 2014. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30:2114–2120. doi:10.1093/bioinformatics/btu 17024695404 PMC 4103590 · doi ↗ · pubmed ↗
- 5Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, Fierer N, Peña AG, Goodrich JK, Gordon JI, et al.. 2010. QIIME allows analysis of high-throughput community sequencing data. Nat Methods 7:335–336. doi:10.1038/nmeth.f.30320383131 PMC 3156573 · doi ↗ · pubmed ↗
- 6Bolyen E, Rideout JR, Dillon MR, Bokulich NA, Abnet CC, Al-Ghalith GA, Alexander H, Alm EJ, Arumugam M, Asnicar F, et al.. 2019. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol 37:852–857. doi:10.1038/s 41587-019-0209-931341288 PMC 7015180 · doi ↗ · pubmed ↗
- 7Callahan BJ, Mc Murdie PJ, Rosen MJ, Han AW, Johnson AJA, Holmes SP. 2016. DADA 2: high-resolution sample inference from Illumina amplicon data. Nat Methods 13:581–583. doi:10.1038/nmeth.386927214047 PMC 4927377 · doi ↗ · pubmed ↗
- 8Rognes T, Flouri T, Nichols B, Quince C, Mahé F. 2016. VSEARCH: a versatile open source tool for metagenomics. Peer J 4:e 2584. doi:10.7717/peerj.258427781170 PMC 5075697 · doi ↗ · pubmed ↗