The complete annotated mitochondrial genome of Cerastoderma glaucum (Bruguière, 1789) from the Baltic Sea
Beata Śmietanka

TL;DR
This paper presents the full mitochondrial genome of the bivalve Cerastoderma glaucum from the Baltic Sea, offering insights into its genetic structure and reproductive traits.
Contribution
The study provides the first complete annotated mitochondrial genome of Cerastoderma glaucum and reveals its lack of mitochondrial heteroplasmy.
Findings
The mitochondrial genome is 14,953 bp long and encodes 13 protein-coding genes, 2 rRNA genes, and 22 tRNA genes.
Baltic and English C. glaucum samples show close genetic affinity with a p-distance of 0.027.
No mitochondrial heteroplasmy was detected, indicating it is not a doubly uniparental inheritance species.
Abstract
Cerastoderma glaucum is a key intertidal bivalve in the north-east Atlantic, serving as a trophic resource for fish and birds. Its mitochondrial genome comprises 14,953 bp, encoding 13 protein-coding genes, 2 rRNA genes, and 22 tRNA genes. Gene structure resembles Cerastoderma edule but lacks an additional methionine tRNA. Phylogenetic analysis revealed close affinity between Baltic and English samples (p-distance = 0.027). No mitochondrial heteroplasmy was detected in mature male gonads, suggesting C. glaucum is not a doubly uniparental inheritance (DUI) species. These findings contribute to understanding mitogenomic variation and reproductive biology in bivalves inhabiting dynamic coastal ecosystems .
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1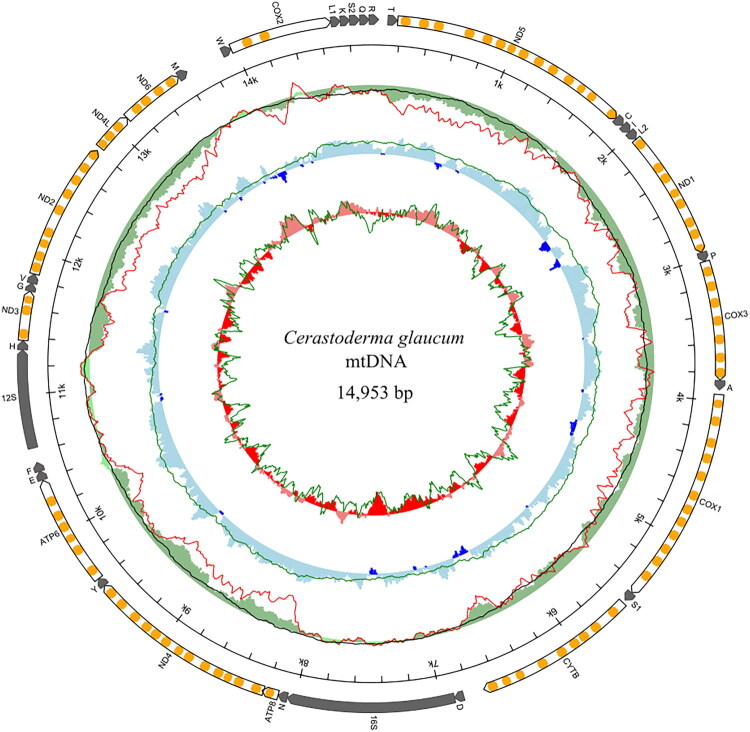 Figure 2
Figure 2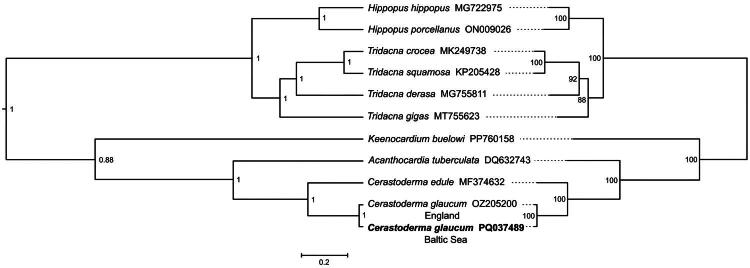 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Biology and Ecology Research · Marine Bivalve and Aquaculture Studies · Marine and coastal plant biology
Introduction
Cockles are considered a very important component of the coastal marine ecosystem and a major food source for many species, especially fish and birds exploring the intertidal environment. Euryhaline bivalve mollusc C. glaucum (Bruguière, 1789) is one of the two main cockles occurring in the coastal areas of the north-east Atlantic from Norway to Senegal, through the Baltic, Mediterranean, Black Sea, the Caspian Sea, and even the Aral Lake, in salinities from 4 to 100 ppt (Russell and Petersen 1973; Hayward and Ryland 1995; Nikula and Väinölä 2003; Malham et al. 2012). C. glaucum prefers non-tidal areas such as lagoons or salt marshes unlike the exploited edible C. edule often found on open coasts and estuaries (Reise 2003). The patterns of inter as well as intra-species genetic diversity are essential in the context of exact identification, management, and protection of natural resources. Cockles are considered important ecosystem engineers that contribute to a range of ecological functions (Carss et al. 2020). The reported declines of cockle populations noticed for many years due to climate events (Rowley et al. 2014), mass mortalities produced by pollution, predation, disease, parasite (De Montaudouin et al. 2009; Longshaw and Malham 2013), failed recruitment and probably most importantly over-fishing (Ducrotoy et al. 1989; Burdon et al. 2014; Villalba et al. 2014) underscores and justifies the critical importance of expanding the genetic resource base for these species (Mahony et al. 2020; Pampín et al. 2023).
Mitochondrial DNA is a useful marker for species identification, phylogeographic and phylogenetic analyses. However, in many bivalve species, a unique doubly uniparental inheritance (DUI) of mtDNA is observed (Skibinski et al. 1994; Zouros et al. 1994). Under this model, heteroplasmic males possess an additional divergent male mitogenome located in gametes and transmitted exclusively to the sons, homoplasmic females possess only female mtDNA transmitted to both offspring. The existence of such a marker seriously complicates the interpretation of mitochondrial DNA-based research; hence, it is crucial to positively identify DUI species.
The complete mitogenome of C. edule is available in GenBank (MF374632) but for the second cockle species C. glaucum, there is published online non-annotated sequence (OZ205200) from England. It is also unclear if these cockles are DUI species.
Here, we report the analysis of the next generation sequencing (NGS) data coming from mature male gonads of C. glaucum sampled in the Baltic Sea, filling this gap.
Materials and methods
The sample of adult C. glaucum (Figure 1) was collected in July 2020 from the Baltic Sea (54.668 N 18.673 E) as the only cockle of Cerastoderma in this sea (Piechocki and Wawrzyniak-Wydrowska 2016). Cockles were sexed by gonads analysis under the light microscope (presence of sperm cells or eggs) and stored at −70 °C. To verify the potential transmission of mtDNA to the offspring according to the DUI, the male specimen was selected for further analysis. Preserved gonadal tissue was deposited at the Institute of Oceanology PAN, Sopot, Poland; contact person and email: Beata Śmietanka [email protected] under the voucher number CG072020. Total DNA extraction from gonads was performed according to the CTAB method (Hoarau et al. 2002). DNA was quality checked by spectrophotometric assessment of concentration, then sent to Macrogen Inc. (Seoul, South Korea) for NGS approach. They prepared the TruSeq NGS library and sequenced it on the Illumina NovaSeq 6000 platform (San Diego, CA), yielding 45,484,508 raw, PE, 150 bp long reads.
Image of the species of Cerastoderma glaucum from the Baltic Sea. The photo was taken by Beata Śmietanka.
A preliminary assembly of raw reads was performed in MEGAHIT (Li et al. 2015), with a k-mer size of 100 bp. The obtained contigs were searched for mitochondrial protein signatures by Wise2 (Birney et al. 2004) and each of them was used as a bait in NOVOPlasty (Dierckxsens et al. 2017). Each NOVOPlasty assembly yielded the same, circularized mitochondrial contig, which was validated by mapping raw reads back on the assembly to the mitogenomes in CLC Genomics Workbench 9.5.5 (QIAGEN, Hilden, Germany). The depth of coverage obtained by mapping raw sequencing reads onto the C. glaucum mitochondrial genome assembly is presented in Supplementary Figure S1. For gene prediction and annotation, the MITOCONSTRICTOR (Lubośny et al. 2022) python script with default parameters running and combining results from the following bioinformatic tools: CRITICA (Badger and Olsen 1999), Wise2 (Birney et al. 2004), GLIMMER (Delcher et al. 1999), ARWEN (Laslett and Canbäck 2008), and Infernal (Nawrocki and Eddy 2013) was used.
For comparative analysis, 11 Cardiidae family mitogenomes were selected based on finding regions of similarity between sequences in Nucleotide BLAST+ (Camacho et al. 2009). The pairwise genetic distance between mitogenomes was calculated as nucleotide p-distance in MEGA11 (Tamura et al. 2021). Reconstructions of phylogenetic relationships, based on 13 protein-coding and two rRNA genes alignment were performed using two programs: BEAST2 (Bouckaert et al. 2014) and Iqtree2 (Minh et al. 2020).
Results
Analysis of the NGS dataset obtained from C. glaucum specimen collected in the Baltic Sea revealed a single putative mitochondrial contig, supporting the presence of a solitary mitogenome (GenBank accession number: PQ037489) with a total length of 14,953 base pairs (bp). The mitogenome of C. glaucum (Figure 2) encodes 13 protein-coding genes: cox1-cox3, cytb, atp6, and atp8, seven NADH subunit genes, two rRNA genes, and 22 tRNA genes: trnT, trnC, trnI, trnL(tag), trnP, trnA, trnS(tga), trnD, trnN, trnY, trnE, trnF, trnH, trnG, trnV, trnM, trnW, trnL(taa), trnK, trnS(tct), trnQ, and trnR. The 22 tRNA genes varied in length from 57 to 69 bp. The nucleotide composition in C. glaucum mtDNA is: 35.4% of T, 24% of G, 23.8% of A, and 16.8% of C with G–C content of 40.8%. The phylogenetic reconstructions (Figure 3) placed the reported Baltic Sea C. glaucum on one clade with C. glaucum (OZ205200) from England with 0.027 nucleotide p-distance for the whole mitogenome sequence, additionally, the genetic variation in the protein-coding region was estimated separately for synonymous and non-synonymous sites, resulting in πS= 0.121 and πa = 0.006, respectively.
Genetic map of Cerastoderma glaucum mitochondrial genome. The white arrows with orange bands represent 13 protein-coding genes with predicted transmembrane domains. The dark arrows represent two rRNA and 22 tRNA genes. The green outer circle reflects the AT-skew and the red line on this circle represents filtered AT-skew, calculated at non-coding regions and the second codon position. A black line represents a filtered AT-skew, calculated at neutral and non-coding positions only. The middle blue circle presenting GC skew is connected with the green line for GC skew at neutral positions. The inner red circle shows local GC content, and the green line reflects GC content at neutral sites. The map and compositional indices were generated with MITOCONSTRICTOR (https://github.com/aburzynski/mitoconstrictor). Transmembrane domains in amino acid sequences of the proteins were predicted with TMHMM (Krogh et al. 2001).
Phylogenetic tree based on comparative analysis of the announced mitogenome of Cerastoderma glaucum from the Baltic Sea (in bold) and 10 mitogenome sequences chosen by Nucleotide BLAST+: Acanthocardia tuberculata (DQ632743, Dreyer and Steiner 2006), Cerastoderma edule (MF374632, unpublished), Cerastoderma glaucum (OZ205200, Wellcome Sanger Tree of Life Programme 2024), Hippopus hippopus (MG722975, Ma et al. 2019), Hippopus porcellanus (ON009026, Ma et al. 2022), Keenocardium buelowi (PP760158, Choi et al. 2024), Tridacna crocea (MK249738, Cai et al. 2019), Tridacna derasa (MG755811, Ma et al. 2018), Tridacna gigas (MT755623, Ma et al. 2020), and Tridacna squamosa (KP205428, Gan et al. 2016). The left phylogram presents the results of Bayesian analysis with node support values equal to 1, whereas the right tree reflects the maximum likelihood analysis with 100% bootstrap values. For BEAST2 reconstruction, the MCMC chain was run in four replicates for 107 generations with sampling at every 10,000th step, relaxed log-normal clock, nucleotide substitution model GTR + I + 4G chosen as the best fitting by the bModelTest package in the BEAST2 software, and the Yule prior for the common tree. For the Iqtree2 method, default parameters, as well as the best-fitting GTR + F + I substitution model and ultrafast bootstrap approximation parameter set to 100,000 replicates were used. For the visualization of obtained relationships, the FigTree v1.4.4 (Rambaut 2018) graphical viewer was applied.
Discussion and conclusions
The reported complete mitochondrial genome of C. glaucum from the Baltic Sea exhibits a length comparable to that of C. glaucum (OZ205200) from England (14,950 bp) and C. edule (MF374632) (14,947 bp), which has a similar gene arrangement. The sequence divergence between the Baltic and English C. glaucum mitogenomes is predominantly associated with differences at synonymous sites, with extremely weak signal of non-synonymous substitutions. This pattern suggests the presence of natural intra-species polymorphism rather than adaptive divergence. No signs of heteroplasmy was detected in the gonadal tissue of male C. glaucum, contrasting with findings in other bivalve species where the NGS approach has revealed the coexistence of two highly divergent mitogenomes in mature male gonads (Śmietanka et al. 2018; Lubośny et al. 2020). These results support the hypothesis that mitochondrial DNA in Cerastoderma is transmitted exclusively through the maternal lineage (standard maternal inheritance – SMI model). The annotated mitogenome of C. glaucum includes 22 tRNA genes. In contrast, the mitogenome of C. edule (MF374632, unpublished) contains an additional tRNA gene, trnM2. However, none of the bioinformatic tools employed in this study identified trnM2 as a functional tRNA gene, suggesting that the annotation of 23 tRNAs in C. edule may include a nonfunctional or pseudogenic artifact. Specifically, the ARWEN tool failed to recognize the trnM-like sequence as a valid tRNA, reinforcing the likelihood of its nonfunctional status. A similar annotation discrepancy has been observed in other Cardiidae species. For instance, Acanthocardia tuberculata (DQ632743) includes a manually predicted trnM2 gene (Dreyer and Steiner 2006), lacking computational validation. Notably, the anticodon of this putative trnM2 gene matches that of the canonical trnM(cat), unlike the trnM(tat) anticodon found in the trnM2 of Mytilus (Hoffman et al. 1992). The analytical pipeline applied in this study did not detect any spurious tRNA genes in DQ632743. Similarly, the mitogenome of Keenocardium buelowi includes only 22 tRNA genes, with a single trnM gene (Choi et al. 2024). The G + C content of the C. glaucum mitogenome is slightly lower (40.8%) than that of C. edule, which reaches 41.7%. These newly obtained mitogenomic data provide a valuable foundation for future studies focused on species identification, population genetics, and phylogeographic analyses within the genus Cerastoderma.
Supplementary Material
Supplementary material for review.docx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Badger JH, Olsen GJ. 1999. CRITICA: coding region identification tool invoking comparative analysis. Mol Biol Evol. 16(4):512–524. 10.1093/oxfordjournals.molbev.a 02613310331277 · doi ↗ · pubmed ↗
- 2Birney E, Clamp M, Durbin R. 2004. Gene Wise and Genomewise. Genome Res. 14(5):988–995. 10.1101/gr.186550415123596 PMC 479130 · doi ↗ · pubmed ↗
- 3Bouckaert R et al. 2014. BEAST 2: a software platform for Bayesian evolutionary analysis. P Lo S Comput Biol. 10(4):e 1003537. 10.1371/journal.pcbi.100353724722319 PMC 3985171 · doi ↗ · pubmed ↗
- 4Burdon D, Callaway R, Elliott M, Smith T, Wither A. 2014. Mass mortalities in bivalve populations: a review of the edible cockle Cerastoderma edule (L.). Estuarine Coast Shelf Sci. 150:271–280. 10.1016/j.ecss.2014.04.011 · doi ↗
- 5Cai S, Mu W, Wang H, Chen J, Zhang H. 2019. Sequence and phylogenetic analysis of the mitochondrial genome of giant clam, Tridacna crocea (Tridacninae: Tridacna). Mitochondrial DNA Part B. 4(1):1032–1033. 10.1080/23802359.2019.1579071 · doi ↗
- 6Camacho C et al. 2009. BLAST+: architecture and applications. BMC Bioinformatics. 10(1):421. 10.1186/1471-2105-10-42120003500 PMC 2803857 · doi ↗ · pubmed ↗
- 7Carss DN et al. 2020. Ecosystem services provided by a non-cultured shellfish species: the common cockle Cerastoderma edule. Mar Environ Res. 158:104931. 10.1016/j.marenvres.2020.10493132501263 · doi ↗ · pubmed ↗
- 8Choi H, Gwon Y, An YK, Eyun S. 2024. Positive selection of mitochondrial cytochrome b gene in the marine bivalve Keenocardium buelowi (Bivalvia, Cardiidae). Animals. 14(19):2812. 10.3390/ani 1419281239409760 PMC 11475489 · doi ↗ · pubmed ↗