Parental age selection in C. elegans influences progeny stress resistance capacity
Bennett T Van Camp, Sean P Curran

TL;DR
Selecting late-reproducing C. elegans parents leads to offspring with reduced stress resistance and shorter lifespan.
Contribution
The study reveals a novel homeostatic mechanism linking parental reproductive timing to offspring health outcomes.
Findings
Late-selected C. elegans progeny show reduced oxidative stress resistance and lifespan.
Late selection improves starvation resistance but does not affect developmental timing or reproduction.
Phenotypes resemble those with SKN-1 activation, suggesting transcriptional remodeling.
Abstract
With parental age rising around the globe, an increased understanding of the impact on health and longevity is needed. Here, we report how the continuous selection of the last progeny during the Caenorhabditis elegans reproductive span results in a diminishment of multiple age-related health measures. After more than 50 generations of late selection, progeny displayed diminished resistance to acute oxidative stress, disrupted partitioning of stored lipids, reduced movement capacity, and an overall shortening of lifespan (36.84% reduction). In contrast, starvation resistance was improved and late selection had negligible effects on developmental timing and total reproductive output that suggests a reduction in lifespan health to preserve reproductive capacity. The phenotypes of late selection are reminiscent of animals with activation of the cytoprotective transcription factor SKN-1 that…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
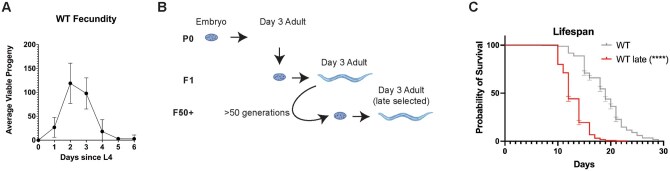 Figure 1
Figure 1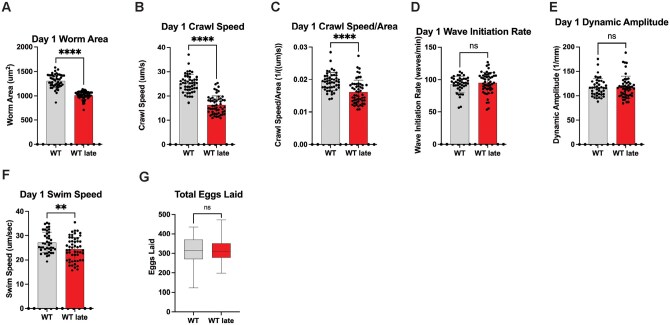 Figure 2
Figure 2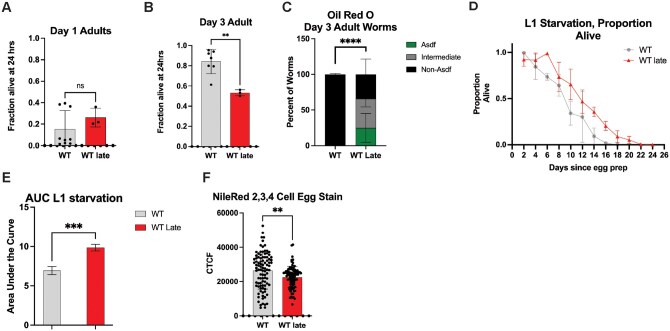 Figure 3
Figure 3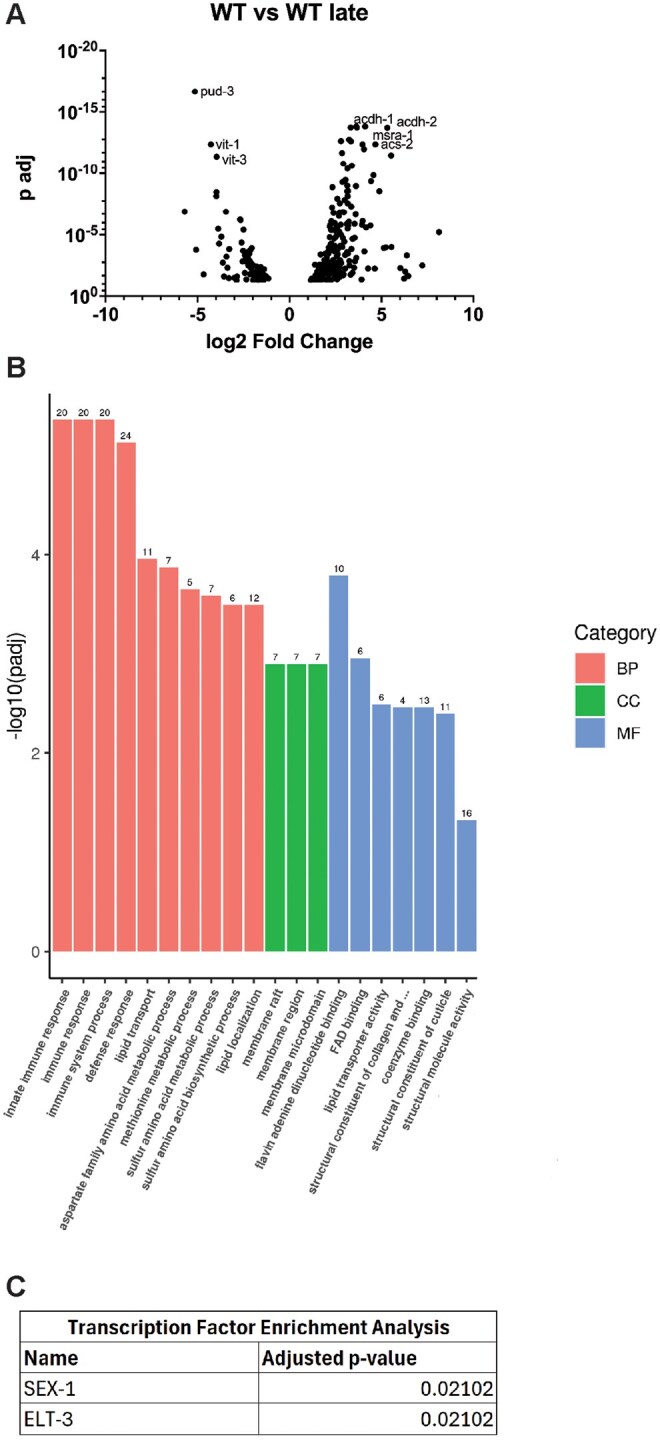 Figure 4
Figure 4- —SPC Hevolution Foundation
- —NIH10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · Reproductive Biology and Fertility · Evolution and Genetic Dynamics
Introduction
The underlying mechanisms of aging have been a topic of intense focus by the research community for their impact on multiple issues of societal importance (e.g., health, lifespan, population dynamics, and evolutionary history.1–4 However, the primary focus of aging research tends to examine the deterioration of a monogenerational cohort of research subjects despite the knowledge that transgenerational effects can be potent drivers of natural aging.4 Reproductive success is an important life history trait, most often considered in the context of parental fitness and behavior.5^,^6 However, age-related reproductive decline also influences offspring health and fitness3^,^7–9 and parental age has been shown to impact a wide variety of offspring phenotypes, including reproductive output, environmental stress resistance, early life survivability, and lifespan.3^,^7–12 In contrast, over the past 55 years, human lifespan has been on the rise in many countries, with a recent decrease observed in the United States13 and a global decrease due to the COVID-19 pandemic.13^,^14 Alongside this trend, the mean parental age has also risen, with the percentage of people becoming first-time fathers over forty doubling.15 It is imperative to understand the effects of late reproduction have on offspring health to understand how these population trends could broadly impact societal health.
These effects have been observed across taxa, ranging from humans, elephants, roundworms, and flies.3^,^7–12 Several potential mechanisms have been described that may contribute to these effects, including a decline in overall gamete quality, a loss of parental capacity to provide somatic support to eggs and sperm,16 increased mutational load in the germline,17–21 and decreased nutrient provisioning to eggs, as observed in older Eupelmus vuilleti mothers.22 Conversely, in Sancassania berlesei, young mothers lay many eggs in nutrient-rich environments but swap in old age to producing fewer, but larger progeny that are able to compete with their older siblings in a more competitive nutrient environment.23 However, there is some evidence that this accelerated growth could be detrimental to the younger offspring in the long term.24^,^25
Although the mechanisms underlying the impact parental age have on offspring fitness have been hypothesized, the effects of multi-generational selection of advanced age requires additional exploration. As stated previously, there is some evidence in Drosophila that poor genetic quality as a result of advanced parental age could lead to a snowball effect in future generations.19^,^20^,^26 However, other studies have shown that selecting for older parental age for several generations in Drosophila produces long-lived offspring.27 Interestingly, studies in C. elegans and D. melanogaster have shown that short-term late selection (3 generations) shortens lifespan and decreased healthspan. However, these effects were significantly modulated with just one generation of swapping to a selection for young parents.28 The mechanism for these effects, their accumulation, and their durability remain poorly understood, as there appears to be a high variability in the observed effects based on the age of selection mating behaviors.16 Additionally, in shorter selections (under 10 generations) significant variability has been observed based on the duration of the selection, with the animals initially showing a decrease in reproduction followed by a normalization to baseline.11 Collectively, we need additional models to better understand how parental age selection impacts health.
To this end, we performed a long-term age selection using Caenorhabditis elegans as a model. To avoid potential short-term effects and short-term intergenerational variability, we performed this selection for 50 generations before collecting data. Below, we detail the healthspan and lifespan effects resulting from this generational selection, as well as a transcriptomic analyses of the selection cohorts. Here, we examine at the physiological outcomes resulting from long-term parental age selection and providing a new experimental model connecting multigenerational selection of late parenthood on offspring health. Overall, we hypothesize that this selection leads to a significant remodeling of homeostatic and stress response pathways, with tradeoffs being made to adapt to the selection process.
Materials and methods
Maintenance of C. elegans strains
Strains were grown at 20°C on nematode growth media (NGM) + streptomycin plates with OP50 food. WT, N2 Bristol strain was used and is the strain WT Late was derived from. All strains were unstarved for at least 3 generations before use.
WT Late worms were grown in conditions identical to unselected WT worms. On day 1-2 of adulthood, adults were moved to a new plate to prevent day 1-2 progeny contamination. On day 3 of adulthood, day 3 progeny were collected and used to propagate the next generation of WT Late worms. This process was repeated 50+ times prior to performing any other assays.
RNAseq analysis
Worms were synchronized overnight as L1s and then dropped on NGM+streptomycin plates with 25x OP50 until L4 and then collected. They were then washed 3x with M9 buffer and frozen at −80°C in TRI reagent until use. Worms were then homogenized and had their RNA extracted using the Zymo Direct-zol RNA Miniprep Kit (Cat. #R2052). Samples were then sequenced and read counts, Differential expression analysis and GO term analysis were reported by Novogene. Adjusted p-value of p < .05 was used as the threshold for significance
Genome-wide sequencing
Genome-wide sequencing was performed as previously described.29 Worms were synchronized overnight as L1s, and then, dropped on NGM + streptomycin plates with 25x OP50 until L4 and then collected. They were then washed 3x with M9 buffer, homogenized, and their DNA was extracted using Zymo Quick-DNA Miniprep Kit (Cat. #D3025). Samples were then sequenced and analyzed by Novogene.
Oil red O staining
Staining was performed as previously described.30^,^31 In brief, worms were synchronized overnight as L1s and then dropped on NGM + streptomycin plates with OP50 for 120hrs and then collected. They were then washed with PBS + triton, then rocked in 40% isopropyl alcohol for 3 min. Worms were then pelleted and treated with ORO in H2O for 2 h. They were then washed in PBS + triton for 30 min and imaged at 20× using LAS X software and Leica Thunder Imager Flexacam C3 color camera. The stained worms were categorized as Asdf (complete somatic lipid depletion), intermediate (incomplete somatic lipid depletion), and non-Asdf (no somatic lipid depletion).
Starvation resistance assay
Worms were egg prepped and synchronized overnight as L1s and then rotated slowly in M9 at 20°C. Ten microliters were removed every 2 days and the larvae bodies were counted on a NGM + streptomycin plate with OP50. They were then allowed to grow for 72 h and recounted to get the proportion alive. The experiment ended once a 0% survival was reached for all conditions.
Egg Nile red staining
Staining was performed as previously described.30 Worms were egg prepped and then the eggs were washed in PBS + triton. Eggs were then incubated in 40% isopropyl alcohol overnight. The next day, the eggs were moved into Nile Red + DAPI staining solution and incubated for 2 h in the dark. They were then washed with PBS + triton and imaged at 63X using LAS X software and Leica Thunder Imager. Corrected total cell fluorescence (CTCF) was then measured in ImageJ and Microsoft Excel.
Oxidative stress assay
Assay was performed as previously described.31^,^32 Synchronous populations of day 3 adults were washed 3x in M9+triton. The worm pellet was aspirated down to 500 µL, and then, 500 µL of 20 mM hydrogen peroxide was then added. They were then incubated on a rotator at 20°C for 25 min. Worms were then washed 3x with M9+triton and dropped onto NGM + streptomycin plates with OP50. Worms were then counted and then counted again 24 h later to get their survival.
Movement/worm area measurements
For all assays, worms were egg prepped and synchronized overnight as L1s. Worms were then added to an OP50 plate and allowed to grow for and 72 h to day 1 adults. Worms were then washed with M9 + triton onto an unseeded NGM plate. The worms were allowed to acclimate for 30+ min to allow the liquid to evaporate. Crawling videos were then taken for 1 min at 7.5 fps. Worm Area was calculated using videos of crawling worms. Two milliliter of M9 was then added to the plate and worms were allowed to acclimate before taking swimming videos. Swimming videos were then taken for 30 s at 14 fps. All imaging was done with the MBF Bioscience WormLab microscope. Analysis was then performed by WormRACER.33
Lifespan assay
Assay was performed as previously described.29^,^34^,^35 Worms were egg prepped and synchronized as L1s overnight. They were then dropped onto NGM + streptomycin and OP50 plates. Worms were moved periodically to remove progeny as needed and kept at 20°C. Worms were scored via prodding with a platinum wire daily. Bagging, vulval bursting, and desiccation on the side of the plate led to censorship of that worm.
RNAi assay
gst4::gfp reporter worms were egg prepped and synchronized in M9 overnight at 20°C. L1s were then dropped onto NGM plates seeded with the appropriate L4440-based RNAi clone. At L4 stage, worms were assessed for changes in reporter expression and scored on a three-point scale (0 OFF, 1 normal, 2 increased, 3 HIGH) in a blinded manner.
Developmental timing assay
Worms were egg prepped and synchronized in M9 overnight at 20°C. Single L1s were then put onto NGM + streptomycin plates with OP50. Worms were then scored every 2 h until they lay their first egg.
Reproductive output assay
Worms were egg prepped and synchronized in M9 overnight at 20 °C. L1s were then put onto NGM + streptomycin plates with OP50. L4s were then singled. Each day, eggs were counted, and the adult worm was moved to a new plate. Assay continued until egg laying ceased.
Statistical analysis
All statistical analysis was done on GraphPad Prism version 9.5.0. Statistical analysis done via Prism were either an unpaired t-test or a Kaplan–Meier simple survival analysis. A two-way ANOVA was used to analyze the OilRedO staining results. p < .05 was the threshold for significance.
Results
Repetitive selection of late progeny reduces C. elegans lifespan
Advanced parental age is known to negatively impact a broad spectrum of healthspan metrics in offspring fitness (e.g., stress resistance, reproductive output, early life health/survival),3^,^7^,^8^,^16^,^26^,^36–40 however, much less is understood about the impact of advanced parental age in a multigenerational context.27^,^41 To further study this phenomenon, a late selection paradigm was applied to a population of wildtype (WT) C. elegans (Figure 1A and B). In this population (hereafter referred to as WT Late), only progeny produced during day 3 of parental adulthood were collected and used to create the next generation, representing the last quartile of reproductive output (Figure 1A). To assess the impact of repetitive and chronic selection of late progeny, the process was repeated for 50 generations before the changes in physiological and molecular markers were assessed.
Chronic selection of late progeny shortens lifespan. (A) Reproductive curve of WT animals (B) Model of the late selection process (C) Lifespan assay of WT and WT Late animals (N = 3, n = minimum of 325 total). Comparisons were made using a Kaplan–Meier simple survival analysis. p < .05 was the threshold for significance. Figure 1B was created using BioRender.
We first measured the lifespan of synchronized populations of WT Late animals as compared to unselected controls and discovered that WT Late worms display a significant diminished life expectancy (Figure 1C and Table S1). In fact, the median lifespan of the WT Late population was 36% reduced compared to unselected controls and with a significant reduction of 31%, 36%, 33%, 20% at quartile 1, 2, 3, 4, respectively. These data reveal that generational selection of the latest progeny in C. elegans drives accelerated aging.
Repetitive selection of late progeny impacts C. elegans healthspan
To understand how the multigenerational selection of the last progeny influences healthspan, we subjected WT Late worms in a battery of physiological assays to define health status and the relationship, if any, to the decrease in lifespan observed. Movement (e.g., speed) is a powerful surrogate assay for organismal health that declines with age.31^,^33^,^35^,^42 As such, we measured movement capacity of age match WT Late animals as compared to unselected WT controls. It should be noted that WT late worms are slightly smaller than their WT counterparts (22% smaller) but controlling for this did not impact the movement comparisons (Figure 2A-C). WT Late worms performed significantly worse than their WT counterparts; displaying a significant reduction in crawl speed (35% slower) and swim speed (10% slower) with no significant reduction in wave initiation rate and dynamic amplitude. (Figure 2B-F). In light of the reproductive selection performed, we also measured the reproductive output of WT Late worms. Surprisingly, we did not observe a significant change in the total number of progeny in WT Late worms as compared to WT animals (Figure 2G). We did note a modest (<5%) delay in the timing of first egg being laid (Figure S1). This suggests that at least one aspect of fitness, specifically reproduction capacity, is preserved despite significant changes in organismal lifespan.
Progeny of late selection maintain reproductive output. (A) Day 1 Worm Area of WT and WT Late animals (N = 3, n = minimum of 49 total) (B) Day 1 Crawl Speed of WT and WT Late animals (N = 3, n = minimum of 49 total) (C) Day 1 Crawl Speed normalized to Area (N = 3, n = minimum of 49 total) (D) Day 1 Wave Initiation Rate of WT and WT Late animals (N = 3, n = minimum of 42 total) (E) Day 1 Dynamic Amplitude of WT and WT Late animals (N = 3, n = minimum of 42 total) (F) Day 1 Swim Speed of WT and WT Late animals (N = 3, n = minimum of 42 total) (G) Total Eggs Laid for WT and WT Late worms (N = 3, n = 36 total)
Generational selection of late progeny alters oxidative and metabolic resistance capacity
A common hallmark of aging4 is the impairment of stress resistance capacity and dysregulation of metabolic homeostasis. To further elucidate the impact of long-term late selection on C. elegans healthspan, the worms were subjected to oxidative stress via hydrogen peroxide exposure as previously described.43 At day 1 of adulthood, WT Late worms performed the same as unselected WT worms (Figure 3A). However, when day 3 adult worms were exposed, WT Late worms performed significantly worse than their unselected counterparts (36% reduced survival) (Figure 3B). This data reveals that repetitive selection of the last progeny leads to a loss of the ability to tolerate acute stress with age.
Progeny of late parental selection display altered stress adaptation capacity. (A) Day 1 oxidative stress assays of WT and WT Late worms (N = 3, n = minimum of 718 total) (B) Day 3 oxidative stress assays of WT and WT Late worms (N = 3, n = minimum of 1113 total) (C) Oil Red O staining of Day 3 worms (N = 3, n = minimum of 328 total)(D) L1 starvation assay of WT and WT Late worms (N = 3, n = minimum of 1183 total) (E) Area under the Curve of the L1 starvation data (N = 3, n = minimum of 1183 total) (F) Nile Red staining of 2, 3, and 4 cell eggs for WT and WT Late animals (N = 3, n = minimum of 92 total).
We next assessed lipid distribution across C. elegans fat storage tissues by staining with the neutral lipid dye Oil Red O (ORO);30 looking specifically for the age-dependent somatic depletion of fat (Asdf), which has been previously described as an intersection between lipid homeostasis and immune response and predictive of lifespan potential.30^,^32^,^44 Aligned with our previous observation documenting the intriguing relationship with lipid homeostasis and stress resistance capacity we observed a significant decrease in the number of worms that do not display the Asdf phenotype in the WT Late population as compared to unselected controls (65% reduction) (Figure 3C and Figure S2A-D); this observation was driven most significantly in worms exhibiting an intermediate phenotype (an incomplete depletion of somatic lipids; Figure S2C). One hypothesis for why somatic lipids are redistributed to the germline is to ensure the survival of the next generation by providing additional resources. To test this idea, we stressed newly hatched larval stage 1 (L1) animals to starvation and measures survival as assessed by the ability to resume development when food is reintroduced.45 Progeny from WT Late animals that are derived from germ cells with extra lipid availability, exhibit a significant increase in their resistance to L1 starvation (42% increase in survival; Figure 3D and E). Specifically, unselected WT worms quickly lose the ability to tolerate starvation and resume development when food is reintroduced while WT Late animals are able to survive the complete absence of food for six additional days without a significant decrease in survival. Although developing oocytes have increased lipid bioavailability, we could not detect a significant increase in total lipids in newly fertilized embryos as measured by quantifying Nile Red stained lipids by fluorescent microscopy. In fact, quantification of two, three, and four cell embryos reveals that WT Late animals harbor less lipids than unselected WT counterparts (14% reduction; Figure 3F).30 Additionally, no significant change was measurable in the distribution of lipids between the somatic AB cell precursor or the germ cell precursor P1 cell (Figure S2E and F).
Repetitive selection of late progeny modifies the steady-state transcriptional landscape
Several of the phenotypes observed in WT Late animals resemble responses influenced by SKN-1 transcriptional activation.31 To investigate whether WT Late animals accumulated a genetic variation that activates SKN-1 we performed genome-wide sequencing (GWS). We identified 78 variants in coding regions; none in known regulators of SKN-1 activity. Nevertheless, we performed RNAi targeting these genes in a strain harboring a gst-4p::gfp reporter that is sensitive to SKN-1 activity, but none induced GFP expression (Table S2). Collectively, these data suggest SKN-1 is not activated to the level observed in response to other classical genetic mutations.31
Despite our inability to clearly define a SKN-1-activated state, to better determine the molecular basis of the phenotypes observed in WT Late animals we next examined the transcriptional landscape by RNAseq. We compared in WT Late animals to age-matched unselected WT animals which revealed significant transcriptional remodeling in L4 stage animals (Figure 4A and B; Table S3); 199 transcripts with increased expression and 93 transcripts with decreased expression. In GO term analyses, lipid homeostasis and innate immune response were among the classes of genes that display the most significant change after late selection (Figure 4B). Specifically, genes involved in innate immune response (eg, acdh-1), lipid transport (eg, vit-1, vit-3), and lipid utilization (eg, acs-2) were affected in late selected progeny (Figure 4A). Genes involved in oxidative stress response (eg, msra-1) were also found to be affected in late selected progeny. (Figure 4A). Although SKN-1 was not demonstrably activated and the expression of common SKN-1 targets were not significantly induced, transcription factor enrichment analysis (TFEA) revealed targets of SEX-1 and ELT-3 were commonly impacted in late selected progeny (Figure 4C) and may contribute to the modified transcriptional landscape that resembles phenotypes previously observed that are a result of increased in SKN-1 cytoprotection.31^,^32^,^44
Late progeny selection alters the transcriptional landscape. RNAseq data of L4 WT and WT Late worms (N = 3) (A) DEG analysis reveals significant changes in both immune response and lipid homeostasis. (B) GO term analysis reveals significant changes in immune response and lipid homeostasis (C) Transcription Factor Enrichment Analysis reveals significant changes in the targets of SEX-1 and ELT-3.
Discussion
Reproductive aging is typically discussed in terms of the health of the parent.5^,^6 However, it has been shown that parental age can also significantly alter the healthspan and lifespan of their progeny.3^,^7^,^8^,^16^,^26^,^36–40 We characterized the impacts of a multigenerational change in parental age by 50 generations of selecting for day 3 progeny, that result in significant alterations to the transcriptomic landscape, diminished overall healthspan, and reduced lifespan. Many of these changes were reminiscent of changes observed in a constitutively active SKN-1 system, with similar timings in the onset of age-dependent phenotypes.31^,^32^,^44
One of the major changes notices is a vast transcriptomic remodeling of lipid homeostasis genes (Figure 4A and B; Table S3). Lipid changes were some of the most substantial changes in the GO term analysis as well as the downregulation of vit-1 and vit-3 and an upregulation of acs-2 (Figure 4A and B). This is consistent with the finding that WT Late worms have a lower egg lipid content that their WT counterparts (Figure 3F) as a knockdown of acs-2 has been shown to increase fatty deposits46 and vitellogenins are well known yolk protein precursors.47 This is consistent with the hypothesis that long-term parental age effects could occur via decreased nutrient provisioning,16 as has been seen previously in Eupelmus vuilleti.22 However, this result was somewhat surprising given that WT Late worms have better L1 starvation resistance and adult WT Late worms Asdf more, a phenotype usually thought to be a form of terminal investment in the progeny44 (Figure 3C-E). This is also surprising, given that previous studies have shown that unselected day 2 and day 3 progeny have increased vitellogenin provisioning and starvation resistance compared to day 1 progeny.48 One potential explanation is that while WT Late eggs may not have more lipids, they may be supplied with an increased amount of other nutrients from the parents. Another potential explanation is that WT Late worms develop slightly slower which could potentially increase L1 starvation resistance as the animals use their energy stores slower (Figure S1A). Interestingly, previous studies have implicated that downregulation of vit-1 and vit-5 (as seen in WT Late animals) can increase starvation resistance in hlh-30 mutant animals.49 This indicates that decreased lipid provisioning can increase L1 starvation resistance in some conditions.49
Separately, Transcription Factor Enrichment Analysis (TFEA) reveals that WT Late worms displayed an enrichment for the targets of SEX-1 and ELT-3 (Figure 4C). This is interesting because SEX-1, a nuclear hormone receptor, is known to impact early development50 and ELT-3, an erythrocytelike transcription factor, is known to impact the expression of SKN-1 targets.51^,^52 It is possible that the affected targets of these transcription factors are at least partially responsible for the reproductive and the phenotypes related to SKN-1 activation observed in WT Late worms. However, none of the GWS hits individually were sufficient to drive a change in gst-4::gfp expression when knocked down via RNAi (Table S2) supporting the conclusion that a remodeling of the transcriptional landscape and not a single genetic variant is responsible for the phenotypes observed.
When looking at movement, WT Late worms exhibit a lower day 1 crawl speed and swim speed with no corresponding deficiencies in wave initiation rate or dynamic amplitude (Figure 2A-F). These phenotypes suggest a decrease in movement effectiveness, but not necessarily a change in rhythm.53 While the exact mechanism of this change is unclear, this could indicate a preserved neurological control of rhythm with deficiencies in the strength of the muscles those neurons signal to.53 As movement is an established predictor of animal health,54 these data reveal a general decline in healthspan in WT Late animals.
The WT Late population also exhibited several other healthspan-related changes, including a decrease in late-life stress resistance and a 36% decrease in median lifespan (Figures 1 and 3; Table S1). This was surprising as similar studies in Drosophila found that long-term parental age selection improved lifespan and stress resistance.27 One potential explanation is that the severity of the selection has a significant impact on the outcome of the selection. Here, the selection was much less severe, beginning just after the drop from peak reproduction while previous studies use a much more extreme cutoff.27 Another note of interest is that this decreased lifespan, healthspan, and stress resistance phenotypes all mimic patterns that have been previously observed in mutants with a constitutively active stress resistance pathway, perhaps indicating that the cumulative effect of late parental age selection is somehow stressful to the animals.31^,^32^,^44 We selected for a minimum of 50 generations to minimize possible epigenetic effects that typically last 3-5 generations that could confound analyses.55^,^56 Although our genome-wide sequencing identified possible mutations, none single-handedly recapitulated the skn-1gf-like phenotype, which suggests that the changes observed are not a result of a monogenic selection. However, it remains possible that the changes could be because of some combination of genetic changes. This lack of clear genetic driver could suggest that causal changes in the epigenome are likely and could drive the altered transcriptional landscape.
Collectively, this study reveals the repetitive and chronic selection of late progeny across generations can significantly alter healthspan and lifespan trajectories. One limitation of this study is the exclusive use of the N2-Bristol genetic background. Future studies using different species, including wild strains isolated from different environments,57 would shed light onto the potentially conserved effect of long-term late selection and may lend further evidence to a potential epigenetic mechanism of these effects. Moreover, additional clarity with regard to the “durability” of these effects when the selection is removed or reversed, will provide clarity if the long-term selection studied here shares molecular mechanisms with short-term selection previously reported.28
In summary, this study provides a new system for studying the impact of long-term late reproductive selection that can function as a powerful model for the current trend of increasing parental age in humans.15 Although further study is necessary to elucidate the exact mechanism of these effects, this could indicate a looming population health crisis should parental age continue to increase.
Supplementary Material
glaf221_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1López-Otín C , Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;1531194-217. 10.1016/j.cell.2013.05.03923746838 PMC 3836174 · doi ↗ · pubmed ↗
- 2Partridge L , Mangel M. Messages from mortality: the evolution of death rates in the old. Trends Ecol Evol. 1999;14:438-442. 10.1016/s 0169-5347(99)01646-810511720 · doi ↗ · pubmed ↗
- 3Hvide HK , Johnsen J, Salvanes KG. Parental age and birth defects: a sibling study. Eur J Epidemiol. 2021;36:849-860. 10.1007/s 10654-021-00734-833761052 PMC 8416834 · doi ↗ · pubmed ↗
- 4López-Otín C , Blasco MA, Partridge L, Serrano M, Kroemer G. Hallmarks of aging: an expanding universe. Cell. 2023;186:243-278. 10.1016/j.cell.2022.11.00136599349 · doi ↗ · pubmed ↗
- 5Lemaitre JF , Gaillard JM. Reproductive senescence: new perspectives in the wild. Biol Rev Camb Philos Soc. 2017;92:2182-2199. 10.1111/brv.1232828374548 · doi ↗ · pubmed ↗
- 6Spanoudakis E , Tavernarakis N. Age-associated anatomical and physiological alterations in Caenorhabditis elegans. Mech Ageing Dev. 2023;213:111827. 10.1016/j.mad.2023.11182737268279 · doi ↗ · pubmed ↗
- 7Lippens C , Faivre B, Lechenault C, Sorci G. Aging parasites produce offspring with poor fitness prospects. Biol Lett. 2017;13:20160888. 10.1098/rsbl.2016.088828202684 PMC 5326515 · doi ↗ · pubmed ↗
- 8Priest NK , Mackowiak B, Promislow DE. The role of parental age effects on the evolution of aging. Evolution. 2002;56:927-935. 10.1111/j.0014-3820.2002.tb 01405.x 12093028 · doi ↗ · pubmed ↗