Mangiferin as a Novel In Vitro Polyphenolic Inhibitor of Amyloid Aggregation
Daniele Florio, Enrico Gallo, Anella Saviano, Anna Schettino, Noemi Marigliano, Ilaria Leone, Francesco Maione, Daniela Marasco

TL;DR
Mangiferin, a natural compound, was found to inhibit amyloid aggregation in lab tests, showing potential for treating neurodegenerative diseases like Alzheimer's.
Contribution
Mangiferin is identified as a novel polyphenolic inhibitor of amyloid aggregation with multitargeted effects.
Findings
Mangiferin significantly inhibited Aβ1–42 and Cterm_mutA aggregation in a concentration-dependent manner.
Mangiferin altered aggregation mechanisms by forming larger oligomers in Aβ1–42 and reducing oligomer size in Cterm_mutA.
Mangiferin showed no cytotoxicity and reduced amyloid-induced toxicity in SH-SY5Y cells.
Abstract
Amyloid aggregation is a pathological hallmark of several neurodegenerative disorders, including Alzheimer’s disease. Polyphenolic compounds are emerging as promising candidates for therapeutic intervention due to their capacity to interfere with multiple stages of amyloidogenesis. In this study, we investigated, in vitro, the antiamyloidogenic potential of mangiferin (MGF), a xanthonoid polyphenol with established pharmacological activity but previously unexplored in the context of amyloid modulation. Using a combination of biophysical, spectroscopic, and microscopic techniques, we assessed the effects of MGF on the aggregation behavior of two distinct amyloidogenic peptides: Aβ1–42 and Cterm_mutA. Thioflavin T (ThT) assays revealed that MGF significantly inhibited aggregation in a concentration-dependent manner, with maximal inhibition at a 1:5 peptide:MGF ratio. Nanoparticle tracking…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 1
1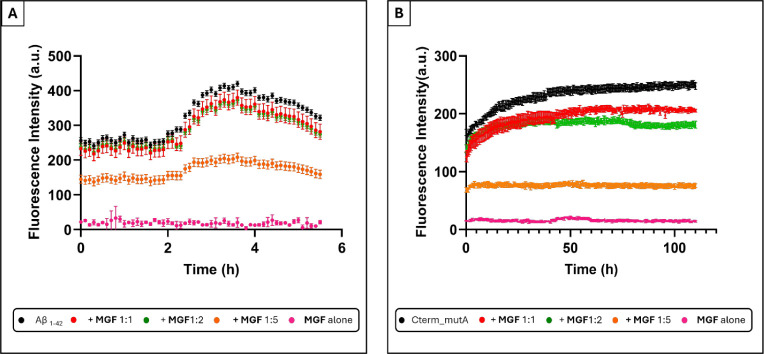 2
2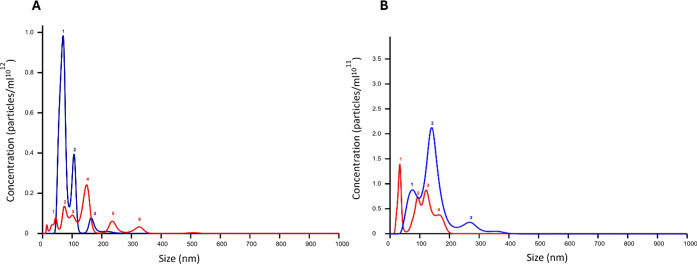 3
3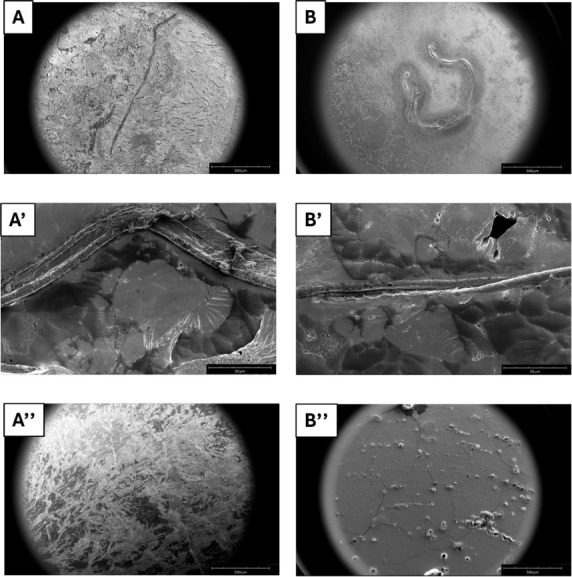 4
4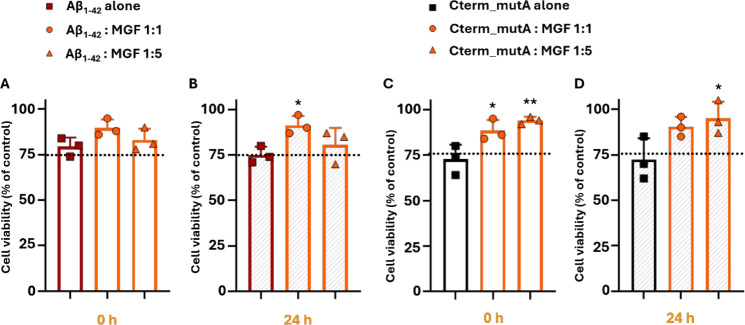 5
5| SEM
Analysis | NTA
Analysis Average Diameters (nm) (Peaks) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Samples | t1/2 (h) | Maximum Intensity (a.u.) | % Inhibition | Diameter (μm) | Length (μm) | 1st | 2nd | 3rd | 4th | 5th | 6th |
| Aβ1–42 | |||||||||||
| 2.5 | 414 | / | 23.2 ± 0.8 | 1231 ± 5 | 70 | 107 | 165 | ||||
| : | 2.6 | 373 | 10 | n.e. | n.e. | ||||||
| : | 2.7 | 361 | 13 | n.e. | n.e. | ||||||
| : | n.e. | 204 | 51 | no fiber | 47 | 72 | 102 | 150 | 237 | 328 | |
| Cterm_mutA | |||||||||||
| 14 | 257 | / | 13 ± 2 | 1404 ± 3 | 75 | 165 | 260 | ||||
| : | 10 | 205 | 20 | ||||||||
| : | 6.5 | 182 | 29 | ||||||||
| : | n.e. | 109 | 58 | no fiber | 34 | 96 | 122 | 165 | |||
- —AORN Foundation10.13039/100002617
- —Universit? degli Studi di Napoli Federico II10.13039/100007195
- —NextGenerationEU10.13039/100031478
- —Ministero dell'Universit? e della Ricerca10.13039/501100021856
- —Ministero dell'Universit? e della Ricerca10.13039/501100021856
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMangiferin and Mango Extracts · Natural Compound Pharmacology Studies · Ginger and Zingiberaceae research
Introduction
Protein aggregation is a hallmark of several misfolding disorders, including Alzheimer’s (AD),? Parkinson’s (PD),? and Huntington’s diseases (HD).? Consequently, developing effective therapeutic strategies to prevent amyloid aggregation is crucial. Over time, numerous inhibitors of amyloid aggregation as antibodies,? peptides,? organic molecules,? metal complexes,? and nanoparticles? have been designed to selectively delay and/or suppress the formation of toxic amyloid species at different stages of aggregation, including intermediate oligomeric forms and mature fibrils.? In this context, natural bioactive compounds derived from food sources have attracted considerable attention in recent years,? particularly polyphenols, due to their distinctive physicochemical properties. ?−? ? Polyphenols are molecules containing one or more phenolic aromatic rings and exhibit a broad spectrum of bioactive properties, including antioxidant, antimicrobial, anticancer, antidiabetic, anti-inflammatory and neuroprotective activities. ?−? ? In several cases, they demonstrated to be able to cross the blood–brain barrier (BBB),? making them attractive as therapeutic agents. ?−? ? Among their bioactive properties, polyphenols are particularly recognized for having a marked capacity for redox homeostasis modulation.? They act as free radical scavengers,? chelators of transition metal ions and binders of cell membranes or other biomolecules.? Furthermore, they modulate enzymatic metabolism by inducing, activating, inhibiting, or protecting oxidase enzymes such as lipoxygenase (LO), cyclooxygenase (COX), myeloperoxidase (MPO), nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) and xanthine oxidase (XO).? Polyphenols protect cellular components by stabilizing radical intermediates through resonance delocalization, thereby preventing oxidative damage and lipid peroxidation. ?,?
In vitro studies showed the potential of various polyphenolic compounds to act as modulators of amyloid aggregation for their ability to directly interact with amino acid and peptide backbone: ?−? ? hydroxyl groups and aromatic rings of polyphenols can interfere with protein self-association by establishing both H-bond and aromatic interactions.? For instance, brazilin-7–2-butenoate (B-7–2-B), a derivative of the natural compound brazilin, exhibited strong inhibitory effects on Aβ_1–42_ and Aβ_1–40_ aggregation? preventing oligomer formation in a concentration-dependent manner, reducing oxidative stress and cytotoxicity induced by Aβ aggregates in neuronal cells, as well as alleviating behavioral and sensory deficits caused by Aβ aggregation in AD Caenorhabditis elegans models.? Similarly, bisdemethoxycurcumin (BDMC), a curcuminoid, decreased the development of β-sheet structures and efficiently suppressed aggregation linked to the ALS-related SOD1 mutant (L38R).? In addition, polyphenols such as forsitoside B (FTS·B) and echinacoside (ECH) have recently demonstrated an attractive antiaggregating ability:? both demonstrated to prevent α-synuclein aggregation by modulating liquid–liquid and liquid–solid phase separation pathways and to reduce its toxicity in neuronal cells.?
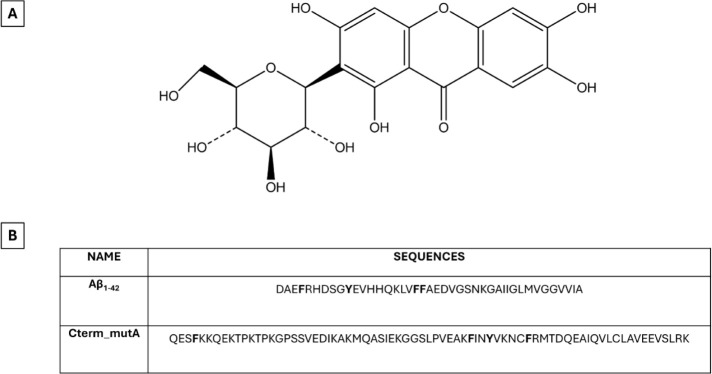
Mangiferin (2-β-d-glucopyranosyl-1,3,6,7-tetrahydroxy-9H-xanthen-9-one, MGF) (FigureA) is a naturally occurring C-glucosylxanthone polyphenol, primarily found in Mangifera indica. Structurally, it consists of a xanthone core substituted with four hydroxyl groups and a β-d-glucopyranosyl moiety at the C-2 position, making it a C-glycoside rather than an O-glycoside. This feature confers it greater stability against enzymatic hydrolysis.? MGF exhibits a broad spectrum of pharmacological properties ?−? ? that are largely attributed to its ability to modulate key intracellular signaling pathways, such as mitogen-activated protein kinase (MAPK), nuclear factor erythroid 2-related factor 2 (Nrf2), nuclear factor-kappa B (NF-κB), adenosine monophosphate-activated protein kinase (AMPK) and the mammalian target of rapamycin (mTOR). ?,?
(A) Chemical structure of MGF compound and (B) primary sequences of Aβ1–42 and Cterm_mutA polypeptides analyzed in this work. Aromatic amino acids are highlighted in bold.
Several studies have demonstrated that MGF exerts potent neuroprotective effects, primarily through the modulation of key signaling pathways involved in cellular survival,? oxidative stress response? and inflammation.? Indeed, MGF regulates the phosphoinositide 3-kinase/protein kinase B (PI3K/Akt), Nrf2/heme oxygenase-1 (HO-1) and extracellular signal-regulated kinase 1/2 (ERK1/2) pathways, which play crucial roles in neuronal survival and the cellular antioxidant defense system.? MGF significantly inhibits the activation of NF-κB and prevents the degradation of its inhibitory protein, IκB. By doing so, it regulates the transcription of a wide array of genes, including those encoding pro-inflammatory cytokines and mediators of neuroinflammation.? Moreover, MGF effectively suppresses the expression of interleukin (IL)-6 and IL-1β, two major pro-inflammatory cytokines known to enhance NF-κB signaling and contribute to neurodegenerative processes.? It also attenuates the activation of the nucleotide-binding oligomerization domain (NOD)-like receptor family pyrin domain containing 3 (NLRP3) inflammasome, a key component in the innate immune response implicated in chronic neuroinflammation in AD and PD.? Importantly, MGF was demonstrated to cross the BBB by exerting a direct neuroprotective action within the central nervous system (CNS). This includes protection against dopaminergic neuronal cell death, a hallmark feature of PD, highlighting its potential as a therapeutic agent in neurodegenerative disorder treatment.?
With the aim of investigating the ability of MGF to directly modulate amyloid aggregation, two amyloid models were selected: (i) Aβ_1–42_ and (ii) Cterm_mutA. The Aβ_1–42_ polypeptide (FigureB) is a cleavage product of the amyloid precursor protein (APP), generated by β- and γ-secretases cleavage. ?,? It is one of the main components of amyloid deposits found in the brains of patients with AD. ?,? Conversely, the Cterm_mutA is a “not neurodegenerative” polypeptide (FigureB) since it is the C-terminal domain (CTD) of nucleophosmin 1 (NPM1) protein in its type A mutation. This mutation is the most common in Acute Myeloid Leukemia (AML) patients. ?,? NPM1 is not a “neurodegenerative protein” as traditionally defined, but many studies ?−? ? ? ? ? ? ? demonstrated that AML mutations determine a great propensity to amyloid aggregation. Since Aβ_1–42_ and Cterm_mutA have demonstrated differences in both the kinetics and morphology of the fibers ?−? ? they were employed as amyloid models in this study. Herein, a wide range of spectroscopic, biophysical, and microscopic techniques, as well as cellular assays, were employed to demonstrate the potential ability of MGF to act as an inhibitor of amyloid aggregation.
Results and Discussion
Kinetic Effects of MGF on Aβ1–42 and
Cterm_mutA Aggregation: ThT Assay
The effects of MGF on the self-aggregation process of Aβ_1–42_ and Cterm_mutA polypeptides were investigated by the Thioflavin T (ThT) assay. ?,? The time course profiles of ThT fluorescence for Aβ_1–42_ and Cterm_mutA, alone and in the presence of MGF, at the indicated molar ratios, are shown in Figure. The nonzero ThT fluorescence values observed at t = 0 for both polypeptides may indicate the presence of a partially preaggregated fraction within the initial samples. The corresponding t 1/2 values (the time at which ThT fluorescence reaches half of its maximum value), maxima of ThT intensity and percentages of inhibition are reported in Table. The results indicated that the presence of MGF significantly alters the kinetic profiles of the two polypeptides, with distinct variations depending on the molar ratio peptide: MGF. Aβ_1–42_ alone exhibited a typical sigmoidal time course of fluorescence, with a t 1/2 of 2.5 h and a maximum intensity of 414 au. The addition of MGF induced a concentration-dependent reduction of signals, with the most significant effect observed at a 1:5 molar ratio, which provided a maximum fluorescence value of 204 au with 51% of inhibition. Similarly, Cterm_mutA showed clear self-aggregation kinetics with a maximum intensity of 257 au with a t 1/2 of 10 h. The addition of MGF caused a decrease in fluorescence intensity at all three analyzed ratios. The reduced ThT fluorescence observed at t = 0 for 1:5 ratio suggests that the interaction between MGF and the amyloid species (already partially aggregated) may alter ThT binding properties even prior to the progression of aggregation and that MGF exerts an “almost immediate” inhibitory effect. Additionally, a reduction in t 1/2 was detected for 1:1 and 1:2 ratios (Table). At the 1:5 ratio, an inhibition of 58% was observed, while control experiments confirmed negligible interference of MGF with ThT fluorescence.
Overlay of time courses of ThT fluorescence emission intensity of: A) Aβ 1–42 in the absence and in the presence of MGF at 1:1, 1:2, and 1:5 peptide-to-compound molar ratio; (B) Cterm_mutA in the absence and presence of MGF at 1:1, 1:2 and 1:5 peptide-to-compound molar ratios. MGF alone was also assessed at 250 μM and 400 μM, corresponding to the highest concentrations used with Aβ1–42 and Cterm_mutA, respectively. Data points represent mean values and error bars indicate standard deviations from three independent experiments.
1: Experimental Values of t 1/2, Maximum Fluorescence Intensity, % of Inhibition, Sizes of Aggregates from SEM and NTA Analyses of Aβ1–42 and Cterm_mutA Polypeptides in the Absence and in the Presence of MGF
Effects of MGF on the Oligomeric States of the Aβ1–42 and the Cterm_mutA: Nanoparticle Tracking Analysis
(NTA)
The effects of MGF on the aggregation of the polypeptides were analyzed using NTA analysis.? This method relies on the detection of light scattering from individual particles and tracking of their trajectories over a brief period (from 30 s to 5 min). Compared to conventional Dynamic Light Scattering (DLS), NTA offers distinct advantages: it enables the quantification of particle number concentration, discriminates among small and weak scatterers in the presence of larger scatterers and provides a size distribution by analyzing the motion of individual particles. NTA experiments were carried out at a 1:5 peptide:MGF molar ratio, after 2 h of stirring, the size distribution of oligomers is reported in Table.
The NTA analysis of Aβ_1–42_ alone (FigureA) showed two peaks centered at 70 and 110 nm in accordance with previous analyses already reported;? the presence of MGF determined: (i) a marked reduction in the total concentration (particles mL^–1^) of aggregates; (ii) a shift of the predominant peak toward larger diameters with respect to Aβ_1–42_ alone; (iii) a more heterogeneous population of oligomers. This heterogeneity is reflected in the appearance of multiple distinct peaks of different sizes (Table). Similar inhibitory effects on Aβ_1–42_ aggregation have also been reported for other natural polyphenols, such as epigallocatechin gallate (EGCG), which significantly reduced aggregate concentration and altered the distribution of oligomeric species.?
Size distribution of particles using NTA of: (A) Aβ1–42 and (B) Cterm_mutA in the absence (blue) and in the presence of MGF (red). Numbers refer to the peaks whose sizes are given in Table .
Cterm_mutA was analyzed, for the first time, by means of NTA and exhibited three main peaks with diameters of 76, 165 and 260 nm after 2 h of aggregation, which are in reasonable agreement with those detected with DLS after 17 h of aggregation.? Also, in this case, the presence of MGF (FigureB) determined a reduction in the aggregate concentrations but caused a shift toward lower diameters with respect to the polypeptide alone. These results indicate that, in solution, MGF alters the aggregation of amyloid models to two different extents: for Aβ_1–42_ stabilized larger aggregates, while for Cterm_mutA smaller species.
MGF Suppresses Fibril Formation: Microscopy Experiments
To get insights into the effects of MGF on the morphology of the fibers derived from Aβ_1–42_ and Cterm_mutA, Scanning Electron Microscopy (SEM) images were recorded at two different times based on the t 1/2 values of polypeptides (Table): 24 h for Aβ_1–42_ and 48 h of aggregation for Cterm_mutA, at 1:5 peptide:MGF molar ratio. As reported in Figure, well-defined fibers were observed for both Aβ_1–42_ (FigureA,A’) and Cterm_mutA (FigureB,B’) alone as already reported. ?,? Specifically, Aβ_1–42_ peptide provided a fiber with an average length of ∼1 mm and a diameter of ∼20 μm (FigureA,A’ and Table) while in the case of Cterm_mutA, the fibers had an average length of ∼1.4 mm and a diameter of ∼13 μm.
*SEM micrographs of: (A-A’) Aβ1–42 alone and (A’’) Aβ1–42 + MGF after 24 h of aggregation; (B–B’) Cterm_mutA alone and (B’’) Cterm_mutA
- MGF after 48 h. Surface overviews at 500 μm (A, A’’, B, B’’) and 50 μm (A’’ and B’’).*
The presence of MGF resulted in the complete suppression of fibrillization, with no aggregates observed (FigureA’’,B’’) like MGF alone (Figure S1A,B). To further corroborate the inhibitory effects of MGF on amyloid fibers, label-free microscopy was employed? and the images were captured both in the blue-emission region (Figure S2A,B)? and in the bright field (Figure S3A-C) after 24 h of aggregation for Aβ_1–42_ and 48 h for Cterm_mutA, in the presence or absence of MGF. Aggregates derived from both polypeptides displayed blue light emission, indicating the presence of amyloid fibers (Figure S2A,B). In contrast, the presence of MGF completely suppressed this emission (Figure S2A,B). Bright-field images (Figure S3A,C) further confirmed these observations.
Cellular Effects of MGF on the Amyloid Cytotoxicity Driven by
Aβ1–42 and Cterm_mutA
The cytotoxicity of MGF was already investigated in murine macrophage J774A.1 cell line.? Since SH-SY5Y is a well-established human neuroblastomal cell line sensitive to amyloid toxicity, to preliminary evaluate the effects of MGF, a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) cell viability assay was performed on SH-SY5Y cells treated with different concentrations of MGF (up to 400 μM at 0 and 24 h). MGF exhibited no significant cytotoxicity under the tested conditions (Figure S4), indicating that MGF is well-tolerated by SH-SY5Y cells within the concentration range evaluated, thereby supporting its suitability for further investigation as a neuroprotective agent. The neuroprotective potential of MGF against amyloid-induced cytotoxicity of Aβ_1–42_ and Cterm_MutA polypeptides at two different time points (0 and 24 h) and two different molar ratios (1:1 and 1:5 peptides:MGF) was assessed. As reported in Figure, both polypeptides alone and untreated showed cytotoxic effects: in the case of Aβ_1–42_, cell viability was significantly reduced compared to untreated controls, with the effect being more pronounced after 24 h of preaggregation with a cell viability <75%, consistent with already reported studies ?,? where prolonged aggregation increased the toxicity of the peptide (FigureA,B).
*Effect of MGF in cell viability of SH-SY5Y human neuroblastoma cells treated with Aβ1–42 alone (50 μM) and Aβ1–42:MGF at 1:1 and 1:5 peptide-to-compound molar ratios after stirring for 0 and 24 h was evaluated (A and B). Similarly, we assessed the protective effect of MGF in SH-SY5Y cells treated with Cterm_mutA peptide alone (80 μM) and Cterm_mutA:MGF at 1:1 and 1:5 peptide-to-compound molar ratios under the same conditions (C and D). The dotted lines indicate the threshold for 75% cell viability. Cell viability (% of control) is presented as the mean ± SD of three independent experiments. Statistical analysis was performed using one-way ANOVA with Bonferroni’s multiple comparisons test. *p ≤ 0.05, *p ≤ 0.01 vs Aβ1–42 alone or Cterm_mutA alone at the corresponding time points.
Similarly, Cterm_mutA induced cytotoxicity at 0 and 24 h, with cell viability falling <72% in both cases (FigureC-D). The incubation with MGF slightly mitigated the cytotoxic effects of both polypeptides. Specifically, for Aβ_1–42_, the condition 1:1 peptide:MGF molar ratio, after 24 h, led to a recovery of cell viability (mean cell viability 91%, p ≤ 0.05), indicating that MGF can counteract the toxicity of Aβ species (FigureA-B). The lack of a clear dose–response effect in the case of Aβ_1–42_, suggested the occurrence of other factors that can affect cell viability. Furthermore, for Cterm_mutA, MGF demonstrated an more pronounced effect since an increase in cell viability was observed at 1:1 molar ratio peptide:MGF after 0 h (mean cell viability 88%, p ≤ 0.05), with a further enhancement at the 1:5 molar ratio (mean cell viability 94%, p ≤ 0.01) (FigureC). Notably, this effect remained significant at the 1:5 ratio even at 24 h (mean cell viability 95%, p ≤ 0.05) (FigureD).
Experimental Section
Reagents
Dimethyl sulfoxide (DMSO), fetal bovine serum (FBS), (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) buffer solution and MGF were purchased from Sigma-Aldrich Co. (now under Merck, Darmstadt, Germany). Dulbecco’s modified Eagle’s medium (DMEM) was obtained from Corning. 3-(4,5-dimethyl-2-thiazolyl)2,5-diphenyl-2H-tetrazolium bromide (MTT) was purchased from BioBasic. Unless otherwise stated, all other reagents were obtained from BioCell (Milan, Italy).
Preparation of MGF for In Vitro Assays
MGF (Product No. M3547, CAS No. 4773–96–0, Merck, Darmstadt, Germany) was dissolved in DMSO to prepare concentrated stock solutions. ?,? To minimize the final concentration of DMSO in cell culture assays, mother stocks were prepared at concentrations four times higher than the desired working concentrations. Specifically, for experiments involving Aβ_1–42_, stock solutions of 200 and 1000 μM were used to achieve final concentrations of 50 and 250 μM, respectively. For Cterm_mutA experiments, stock solutions of 320 μM and 1600 μM were prepared to obtain final concentrations of 80 μM and 400 μM. All stock solutions were freshly prepared and diluted in culture medium immediately prior to use. The final DMSO concentration in each well was carefully controlled to remain below levels known to induce cytotoxicity or exert pharmacological activity per se. Vehicle controls were included in all experiments and prepared under identical conditions to account for any solvent-related effects.
Polypeptides
The Aβ_1–42_ and Cterm_mutA polypeptides (sequences are reported in FigureB) were purchased from NovoPro Bioscience Inc. (Shanghai, China). Both peptides were treated with 1,1,1,3,3,3-hexafluoro-2-propanol (HFIP) to guarantee a monomeric state, lyophilized and stored at −20 °C until use.
Fluorescence Assays
ThT emission assays of Aβ_1–42_ alone (50 μM) and in the presence of MGF at 1:5 peptide:compound molar ratio in 50 mM NaCl, 20 mM phosphate buffer (pH 7.4)/DMSO 2% (v/v), using a ThT final concentration of 5 μM, were carried out in black plates (96-well) under stirring on a fluorescence reader Envision 2105 (PerkinElmer). Measurements were collected every 7 min (λex= 440 nm and λem= 483 nm). Assays were performed in duplicate at 25 °C. For Cterm_mutA alone (80 μM) and in the presence of MGF at 1:5 peptide:compound molar ratio, the experiments were performed in 50 mM phosphate buffer (pH 7.4)/DMSO 2% and 50 μM of ThT and were analyzed using a CLARIOstar fluorescence microplate reader (BMG Labtech) at 25 °C in black, clear-bottomed 96-well half-area polystyrene plates with nonbonding surface (Corning #3881) covered with aluminum thermowell sealing tape (Corning #6570). The experiments were performed in 100 μL aliquots in triplicate.
NTA Measurements
The NTA measurements were carried out using a nanosight NS300 instrument (Alfatest, Italy). Samples of Aβ_1–42_ and Cterm_mutA at a concentration of 100 μM, in the absence and in the presence of MGF (1:5 peptide:compound molar ratio), after 2 h of aggregation, were 1000-fold diluted in Milli-Q water to a final volume of 1 mL and were injected into the sample chamber using a syringe. The dilution was done in accordance with the ideal particle-per-frame value (20–100 particles/frame). The following settings were chosen according to the manufacturer’s software manual (NanoSight NS300 User Manual, MAN0541–01-EN-00, 2017).?
SEM Analysis
The two polypeptides, Aβ_1–42_ (50 μM) and Cterm_mutA (80 μM), alone and in the presence of MGF compound (1:5 peptide:compound molar ratio), were morphologically analyzed after 24 and 48 h of aggregation, respectively, using field-emission SEM (Phenom_XL, Alfatest, Milan, Italy). After this time, 50 μL of solution was drop-cast on an aluminum stub and dried under vacuum to prepare the samples. For 75 s, a thin layer of gold was sputtered at a current of 25 mA. Following the introduction of the sputter-coated samples into the specimen chamber, micrographs were taken using a secondary electron detector (SED) at an accelerating voltage of 10 kV. MGF compound alone (500 μM) was analyzed as a control.
Fluorescence Microscopy
Aβ_1–42_ and Cterm_mutA samples employed for the SEM experiments were drop-cast on clean coverslip glass, dried and imaged with fluorescence microscopy. Fluorescence images were captured with an automated upright microscope system (Leica DM5500 B) coupled with Leica Cytovision software.
Cell Culture
SH-SY5Y human neuroblastoma cell lines (CRL-2266, ATCC, Manassas, VA, USA) were cultured in 100 × 20 mm dishes (1 ×10^6^ cells/dish) in DMEM (Corning; Product No. 10–013-CV) supplemented with 10% FBS (Sial; Product No. YourSIAL-FBS-SA), 100 U mL^–1^ penicillin, 100 μg mL^–1^ streptomycin (Corning; Product No. 30–002-CI) and 25 mM HEPES (Sigma-Aldrich; Product No. H0887). Cells were maintained in a humidified atmosphere of 5% CO_2_ at 37 °C and passaged upon reaching 80% confluence. ?,?
MTT Assay
The ability of MGF to reduce the neurotoxicity of Aβ_1–42_ and Cterm_mutA polypeptides was evaluated in a human neuroblastoma cell line by using the MTT assay. As a preliminary step, the assay was first performed to assess whether MGF exerted any cytotoxic effects on this specific cell line. To this aim, SH-SY5Y cells were seeded at a density of 2.5 × 10^4^ cells per well in 96-well plates, allowed to adhere overnight, and subsequently treated with MGF at concentrations of 50, 80, 250 and 400 μM (after 0, 24 and 48 h of stirring), with cell viability assessed at 24 h time-point. Subsequently, cells were treated with Aβ_1–42_ peptide (50 μM), either alone or in combination with MGF at 1:1 and 1:5 peptide:compound molar ratios (after 0 and 24 h of stirring). Likewise, the Cterm_mutA peptide (80 μM) was tested under the same conditions. Control cells were incubated with DMSO diluted in the cell culture medium at the same % used for treatments. At the selected time point (24 h), 10 μL of MTT (BioBasic; Product No. T0793) solution (5 mg mL^–1^ in phosphate-buffered saline, PBS; pH 7.4) was added to each well and the plates were incubated for 3 h at 37 °C in the dark. Then, the medium was removed and the resulting formazan crystals were dissolved in 150 μL DMSO for 15 min. The spectrophotometric absorbance was measured using a microtiter enzyme-linked immunosorbent assay reader (Multiskan GO Microplate Spectrophotometer; Thermo Scientific) at 540 nm. The percentage of cell viability was determined by the following formula: OD of treated cells/OD of control × 100.? Each experiment was performed in biological triplicate.
Statistical Analysis
All statistical analyses were performed in accordance with established guidelines for experimental design, data analysis and transparent reporting. Data from three independent experiments (n = 3) are reported as the mean ± standard deviation (SD). For ThT fluorescence assays, error bars indicate the SD of three independent replicates. For cytotoxicity assays, statistical significance between groups was assessed by one-way ANOVA followed by Bonferroni’s post hoc test for multiple comparisons, with p ≤ 0.05 considered significant. Analyses were performed using GraphPad Prism 8.0 (GraphPad Software, San Diego, CA, USA).
Conclusions
In this study, we aimed to evaluate the antiamyloidogenic activity of MGF using a multitiered approach that combined spectroscopic and microscopic methodologies with cellular assays. We investigated its effects on the aggregation behavior of two structurally distinct amyloidogenic peptides: Aβ_1–42_, a prototypical peptide implicated in AD and Cterm_mutA, a synthetic model peptide characterized by aggregation driven predominantly through π–π interactions.
ThT fluorescence assays revealed that MGF exerts an inhibitory effect on amyloid aggregation for both peptides, with maximal inhibition observed at a 1:5 peptide:MGF molar ratio. This ratio was subsequently employed in NTA and electron microscopy studies. Despite the distinct aggregation mechanisms of Aβ_1–42_ and Cterm_mutA-driven predominantly by hydrophobic/electrostatic interactions and aromatic stacking, respectively, MGF effectively inhibited aggregation in both cases. The data suggest that MGF could disrupt peptide self-association through multiple interaction modalities: its aromatic scaffold is likely critical for disrupting π-stacking in Cterm_mutA, while its polyhydroxylated structure facilitates hydrogen bonding and electrostatic interference in Aβ_1–42_ fibrillogenesis. ?,? This hypothesis, however, requires further, more detailed structural studies. These differences were further supported by NTA and microscopy data, which showed a stabilization of larger, nonfibrillar oligomeric species in the case of Aβ_1–42_, consistent with fibrillation arrest and a marked reduction in oligomer size for Cterm_mutA, consistent with disruption of aromatic interactions.
Importantly, MGF displayed no intrinsic cytotoxicity in SH-SY5Y neuroblastoma cells. Moreover, cotreatment with MGF slightly ameliorated the cytotoxic effects induced by both Aβ_1–42_ and Cterm_mutA aggregates, underscoring its neuroprotective potential.
In conclusion, our findings suggest MGF as a starting-point molecule to develop modulators of amyloid aggregation, with potential for applications in neuroprotection, neurodiagnostics and the development of novel antiamyloid therapies. To fully translate these findings, comprehensive preclinical and in vivo studies are now critically needed to elucidate the complete therapeutic potential and mechanisms of action.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Murphy M. P.Le Vine H.III Alzheimer’s disease and the amyloid-β peptide J. Alzheimers Dis.201019131110.3233/JAD-2010-122120061647 PMC 2813509 · doi ↗ · pubmed ↗
- 2Calo L.Wegrzynowicz M.Santivañez-Perez J.Grazia Spillantini M.Synaptic failure and α-synuclein Mov. Disord.201631216917710.1002/mds.2647926790375 · doi ↗ · pubmed ↗
- 3Arrasate M.Finkbeiner S.Protein aggregates in Huntington’s disease Exp. Neurol.2012238111110.1016/j.expneurol.2011.12.01322200539 PMC 3909772 · doi ↗ · pubmed ↗
- 4Haddad H. W.Malone G. W.Comardelle N. J.Degueure A. E.Poliwoda S.Kaye R. J.Murnane K. S.Kaye A. M.Kaye A. D.Aduhelm, a novel anti-amyloid monoclonal antibody, for the treatment of Alzheimer’s Disease: A comprehensive review Health Psychol. Res.20221023702310.52965/001c.3702335910244 PMC 9334157 · doi ↗ · pubmed ↗
- 5Mitra A.Sarkar N.Sequence and structure-based peptides as potent amyloid inhibitors: A review Arch. Biochem. Biophys.202069510861410.1016/j.abb.2020.10861433010227 · doi ↗ · pubmed ↗
- 6Ren B.Liu Y.Zhang Y.Cai Y.Gong X.Chang Y.Xu L.Zheng J.Genistein: A Dual Inhibitor of Both Amyloid beta and Human Islet Amylin Peptides ACS Chem. Neurosci.2018951215122410.1021/acschemneuro.8b 0003929432676 · doi ↗ · pubmed ↗
- 7Gomes L. M.Bataglioli J. C.Storr T.Metal complexes that bind to the amyloid-β peptide of relevance to Alzheimer’s disease Coord. Chem. Rev.202041221325510.1016/j.ccr.2020.213255 · doi ↗
- 8Lin H.-C.Ho M.-Y.Tsen C.-M.Huang C.-C.Wu C.-C.Huang Y.-J.Hsiao I.-L.Chuang C.-Y.From the cover: comparative proteomics reveals silver nanoparticles alter fatty acid metabolism and amyloid beta clearance for neuronal apoptosis in a triple cell coculture model of the blood–brain barrier Toxicol. Sci.2017158115116310.1093/toxsci/kfx 07928460142 · doi ↗ · pubmed ↗