An unknown apocarotenoid signal alters plant development by modulating shoot and root apical meristem activities
James M Bradley

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
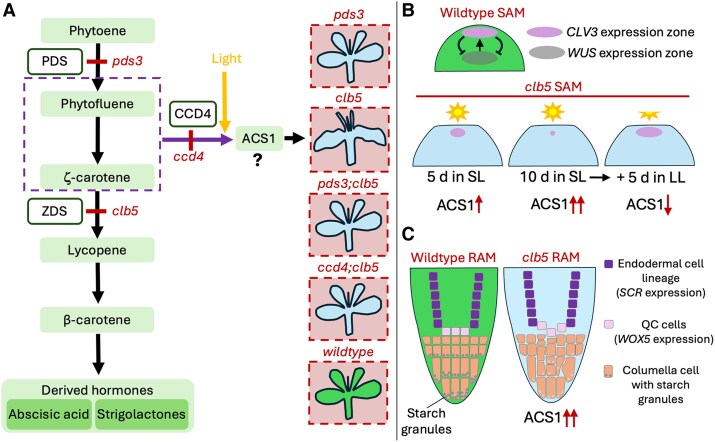 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhotosynthetic Processes and Mechanisms
The plant body plan is directed via organogenesis from stem cell niches in the shoot and root apical meristems. The process of organogenesis is plastic and can be modulated by abiotic cues and endogenous signals to allow the plant to produce new organs that are appropriately acclimated to their environment (Bar and Ori 2014). Endogenous signals are also vital to coordinate the differentiation of specialized cell types and their plastids (Sierra et al. 2023). For example, during the process of leaf development, proplastids must commit to differentiate into chloroplasts to conduct photosynthesis. In the columella cells of the root, proplastids differentiate into starch-accumulating amyloplasts to contribute to gravitropic responses (Sierra et al. 2023). Yet many plastid-resident proteins are encoded in the nuclear genome and must be imported during the process of plastid differentiation (Woodson and Chory 2008). There must therefore be signals that originate from the plastids and transmit information to the nucleus to coordinate plastid developmental status with nuclear gene expression (Woodson and Chory 2008; Sierra et al. 2022, 2023).
Carotenoids are plastid-derived orange, yellow, or red pigments present in all photosynthetic organisms that primarily serve photoprotective roles in chloroplasts (Nisar et al. 2015). These molecules are also responsible for the colors of flowers and fruits and are derived into an array of secondary metabolites, collectively called apocarotenoids (Nisar et al. 2015; Hou et al. 2016). Since carotenoids are such an important constituent of chloroplasts, it is perhaps unsurprising that their apocarotenoid derivatives have been proposed as signals to communicate information between the plastids and the nucleus during leaf development (Sierra et al. 2023). Indeed, some apocarotenoids are already well known to regulate plant growth and development. For example, the hormones abscisic acid and strigolactone are apocarotenoid derivatives regulating diverse aspects of plant development from shoot branching to germination (Umehara et al. 2008; Toh et al. 2012; Nisar et al. 2015). Besides these well-characterized examples, there are predicted to be many uncharacterized apocarotenoid small molecules that play roles in plant development and signaling with plant-associated organisms (Moreno et al. 2021).
In the last decade, genetic evidence has emerged for an unknown apocarotenoid signal that coordinates leaf development with chloroplast biogenesis (Hou et al. 2016; Fig. A). In 2014, Avendaño-Vázquez et al. became interested in a mutant called chloroplast biogenesis 5 (clb5) that arrests proplastid-to-chloroplast development early during differentiation. The clb5 homozygote has an albino phenotype that maps to a mutation in the ZDS gene involved in carotenoid biosynthesis. This mutant also has a striking leaf defect in which radially symmetric needle-like leaves form in lieu of wild type laminar leaves. This developmental defect is specific to clb5, as other loss-of-function carotenoid biosynthetic albino mutants, such as pds3, display laminar leaves. Neither abscisic acid nor strigolactones, both derived from carotenoids, rescue the clb5 leaf defect, indicating that the defect is not due to deficiencies of these hormones (Avendaño-Vázquez et al. 2014). However, genetic ablation of PDS, encoding the biosynthetic enzyme upstream of ZDS, could rescue the leaf defect, suggesting that the unknown apocarotenoid signal (which they called ACS1) is derived from cis-carotenoids that accumulate upstream of ZDS and below PDS (Avendaño-Vázquez et al. 2014; Hou et al. 2016). Further epistasis analysis between clb5 and plastid-localized carotenoid cleavage dioxygenase (CCD) genes identified the clb5 ccd4 double mutants that could partially rescue leaf shape, indicating that CCD4 promotes ACS1 synthesis (Avendaño-Vázquez et al. 2014). The leaf defect could also be rescued by reducing light fluency, suggesting that light promotes ACS1 accumulation (Escobar-Tovar et al. 2021). Indeed, light has been implicated in the nonenzymatic production of other apocarotenoids, such as β-ionone (Hou et al. 2016). The clb5 mutant also shows strong downregulation of nuclear-encoded genes required for correct chloroplast biogenesis (Hou et al. 2016), suggesting that ACS1 acts as a retrograde signal communicating information from the plastid to the nuclear genome and coordinating plastid-to-chloroplast differentiation with proper leaf development.
In this issue of Plant Physiology, Sierra et al. (2025) further interrogate the developmental defects induced by ACS1 in the clb5 mutant. Unlike previous studies with clb5, here the authors used light as a tool to modulate ACS1 accumulation. By shifting plants from standard light (SL; high ACS1) to low light (LL; low ACS1), the authors could infer the developmental window within which ACS1 acts to affect leaf development. They grew clb5 and pds3 (control) plants in SL conditions for increasing periods of time (4–10 d). If left in SL conditions, clb5 plants developed radially symmetric primary leaves. However, shifting plants to LL after 4 to 5 d in SL resulted in most plants developing laminar leaves, whereas shifting plants after 6 d in SL saw the development of radially symmetric primary leaves. Interestingly, these later-shifted plants developed a second pair of leaves that had acquired a laminar form. The authors concluded that there must be a defined developmental window during which ACS1 affects the fate of cells destined to become leaves, suggesting a potential role for ACS1 in the shoot apical meristem (SAM), where leaf organogenesis occurs.
Upon inspection, the authors indeed noted an irregular SAM in clb5, which lacked the typical dome shape of wild type. The SAM is maintained through a negative regulatory loop involving the small secreted peptide CLAVATA3 (CLV3) and the transcription factor WUSCHEL (WUS). WUS is expressed in the SAM organizing center, a region just below the stem cell niche at the shoot apex, and is a positive regulator of CLV3 expression, which itself negatively regulates WUS and stem cell accumulation (Clark et al. 1995; Laux et al. 1996; Fletcher 2018; Fig. B). To investigate SAM organization, Sierra et al. (2025) introduced pCLV3::CLV3-GUS and pWUS::GUS reporters into wild type and clb5 backgrounds. They did not observe WUS expression in the clb5 mutant, even after the emergence of the second leaf pair. However, CLV3 expression was clearly seen in 5-d-old seedlings and decreased over time under SL conditions so that by day 10 expression was very low. This suggested that the accumulation of ACS1 leads to a depletion of the stem cell population in clb5. The absence of WUS expression in clb5 is consistent with the finding that WUS is not required to establish the SAM stem cell population but is required to maintain meristem integrity during growth (Laux et al. 1996; Zhang et al. 2017). The authors then switched their SL-grown 10-d-old clb5 plants to LL for a further 5 d and found that CLV3 expression recovered, and this correlated with the development of laminar leaves. Given the dysregulation of the CLV3-WUS module in the SL-grown clb5 mutant, the authors thus concluded that ACS1 disrupts SAM integrity. However, since shifting to LL rescued CLV3 expression, the authors suggested that ACS1 causes a reversible developmental effect rather than a toxic by-product that irreversibly damages plant development.
The authors then turned their attention to below-ground effects and assessed the role that ACS1 might play in root apical meristem (RAM) organization and function. The RAM consists of distinct stem cell initials maintained adjacent to rarely dividing quiescent centre (QC) cells (Sarkar et al. 2007; Dubrovsky and Vissenberg 2021). The stem cell initials divide asymmetrically, with the daughter cells more distal from the QC cells going on to differentiate into various cell types of the root (Petricka et al. 2012; Dubrovsky and Vissenberg 2021). The stem cell initials just below the QC cells, for example, give rise to differentiated columella cells (Petricka et al. 2012). Just as in the SAM, small peptide ligands and transcription factors are involved in maintaining stem cell niches in the root (Petricka et al. 2012). One key transcription factor is WUSCHEL-related homeobox 5 (WOX5). This protein is expressed in QC cells and non–cell autonomously maintains the pluripotency of surrounding stem cells (Petricka et al. 2012). For example, in wox5 mutants, columella stem cell initials begin to differentiate and acquire starch granules (Sarkar et al. 2007). Another important regulator of the RAM is the GRAS transcription factor scarecrow (SCR), which is required for QC specification and asymmetric division of surrounding stem cells (Petricka et al. 2012).
Accordingly, Sierra et al. (2025) used proWOX5::GFP and proSCR::SCR-GFP reporters to characterize the RAM of clb5 plants. Analysis of 5- and 10-d-old plants grown in SL showed no clear differences in expression patterns from the WOX or SCR promoters, as compared with pds3 or wild type plants. However, the authors did note misaligned QC cells and irregular divisions among the columella cells, suggestive of additional stem cell divisions (Fig. C). The same was not true for stem cell initials above the QC that go on to differentiate into the vasculature or epidermal cell layers—these appeared normal. The columella defects prompted closer investigation. During columella cell differentiation, the cells acquire starch-containing amyloplasts, which are important for graviperception and gravitropism (Sierra et al. 2023). Upon staining roots, Sierra et al. (2025) observed reduced starch levels in the clb5 columella cells as compared with pds3 and wild type lines, suggesting that clb5 might be less able to sense gravity. Indeed, when they rotated plants by 90°, the roots of clb5 plants were unable to fully adjust their root tips to account for the change in gravity as compared with pds3 or wild type plants. Despite this, Sierra et al. (2025) did detect some starch in clb5, suggesting that amyloplasts had successfully differentiated in the columella cells, unlike the plastids located in leaves that were completely unable to differentiate into chloroplasts.
In sum, Sierra et al. (2025) have further probed the developmental consequences of overproduction of the unknown small molecule ACS1 in the clb5 background. Specifically, they show that high ACS1 levels, which are associated with SL conditions, affect shoot and root meristem organization. Using shift experiments from SL to LL, the authors demonstrated that ACS1 is not a toxic product that causes irreversible developmental damage but rather a dynamic signal that is reversible and acts during a defined developmental window to shape leaf organogenesis. For wild type, the implication is that environmental and metabolic cues may be “read” by the plastids (where ACS1 is assumed to be produced) and communicated to the nucleus by fine-tuning ACS1 levels. This in turn would affect the shoot and root meristem activities, leading to an altered plant body plan more suited to the environment. Of course, the burning questions remain. First, what is the nature of ACS1? Second, what are the direct targets of ACS1? To answer the latter, forward genetics could prove useful to identify mutants that suppress the clb5 radially symmetric leaves; at least a subset of suppressors would be direct ACS1 targets.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Avendaño-Vázquez A-O, Cordoba E, Llamas E, San Román C, Nisar N, De La Torre S, Ramos-Vega M, Gutiérrez-Nava MDLL, Cazzonelli CI, Pogson BJ, et al An uncharacterized apocarotenoid-derived signal generated in ζ-carotene desaturase mutants regulates leaf development and the expression of chloroplast and nuclear genes in Arabidopsis. Plant Cell. 2014:26(6):2524–2537. 10.1105/tpc.114.12334924907342 PMC 4114949 · doi ↗ · pubmed ↗
- 2Bar M, Ori N. Leaf development and morphogenesis. Development. 2014:141(22):4219–4230. 10.1242/dev.10619525371359 · doi ↗ · pubmed ↗
- 3Clark SE, Running MP, Meyerowitz EM. CLAVATA 3 is a specific regulator of shoot and floral meristem development affecting the same processes as CLAVATA 1. Development. 1995:121(7):2057–2067. 10.1242/dev.121.7.2057 · doi ↗
- 4Dubrovsky JG, Vissenberg K. The quiescent centre and root apical meristem: organization and function. J Exp Bot. 2021:72(19):6673–6678. 10.1093/jxb/erab 40534562009 · doi ↗ · pubmed ↗
- 5Escobar-Tovar L, Sierra J, Hernández-Muñoz A, Mc Quinn RP, Mathioni S, Cordoba E, Colas Des Francs-Small C, Meyers BC, Pogson B, León P. Deconvoluting apocarotenoid-mediated retrograde signaling networks regulating plastid translation and leaf development. Plant J. 2021:105(6):1582–1599. 10.1111/tpj.1513433340183 · doi ↗ · pubmed ↗
- 6Fletcher JC . The CLV-WUS stem cell signaling pathway: a roadmap to crop yield optimization. Plants. 2018:7(4):87. 10.3390/plants 704008730347700 PMC 6313860 · doi ↗ · pubmed ↗
- 7Hou X, Rivers J, León P, Mc Quinn RP, Pogson BJ. Synthesis and function of apocarotenoid signals in plants. Trends Plant Sci. 2016:21(9):792–803. 10.1016/j.tplants.2016.06.00127344539 · doi ↗ · pubmed ↗
- 8Laux T, Mayer KFX, Berger J, Jürgens G. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development. 1996:122(1):87–96. 10.1242/dev.122.1.878565856 · doi ↗ · pubmed ↗