A Rare Case of Profound Pulmonary Venous Malformation in an Elderly Individual With Three Major Comorbidities: Genetic Insights
Andrey Frolov, Sophia Izhar, Samantha M Spence, Beker Karadaghy, Madeleine Schwab, Yun Tan, Daniel T Daly

TL;DR
This paper reports a rare case of an elderly man with an unusual number of pulmonary veins and multiple health issues, revealing genetic factors that may link heart and cancer conditions.
Contribution
The study identifies novel genetic variants and potential pleiotropic genes associated with anomalous pulmonary venous development and comorbid heart and cancers.
Findings
Whole exome sequencing identified 80 rare genes with pathological variants linked to angiogenesis, cardiogenesis, and cancer.
The CRACD gene is highlighted as potentially important in anomalous pulmonary venous development.
A heart failure↔colorectal/prostate cancer axis is suggested involving the C2ORF88 gene.
Abstract
Pulmonary veins play a very important role in normal human physiology by providing oxygenated blood flow to the heart, as well as in human pathophysiology by inducing and controlling atrial fibrillation and participating in the propagation of pulmonary neoplasms, such as non-small cell lung cancer. Gaining additional insights into the mechanism(s) of pulmonary venous development (PVD) would benefit our understanding of heart and lung function under normal and clinical conditions. In the current report, we present the results of postmortem genetic screening of an 87-year-old male with a remarkably high number of pulmonary veins - 12 in total - with normal connection, who died of arteriosclerotic heart disease and heart failure with additional comorbidities being prostate and colon cancers. To advance our understanding of molecular mechanisms underlying such anomalous PVD, genetic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
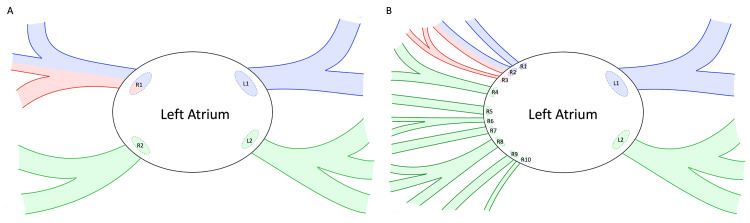 Figure 1
Figure 1| Gene | Protein Function |
| ABCB9 | ATP Binding Cassette Subfamily B Member 9. |
| ANTXR1 | ANTXR Cell Adhesion Molecule 1 (TEM8). |
| ARMT1 | Acidic Residue Methyltransferase 1. |
| ASB10 | Ankyrin Repeat And SOCS Box Containing 10. |
| ATM | ATM Serine/Threonine Kinase. |
| BDP1 | B Double Prime 1, Subunit of RNA Polymerase III. |
| C2ORF88 | Chromosome 2 Open Reading Frame 88 (Small Membrane A-Kinase Anchor Protein, SMAKA Protein). |
| CD19 | CD19 Molecule. |
| CIBAR2 | CBY1 Interacting BAR Domain Containing 2 (FAM92B). |
| CITED4 | Cbp/P300 Interacting Transactivator With Glu/Asp Rich Carboxy-Terminal Domain 4. |
| CMA1 | Chymase 1. |
| COL22A1 | Collagen Type XXII Alpha 1 Chain. |
| CPXM2 | Carboxypeptidase X, M14 Family Member 2. |
| CRACD | Capping Protein Inhibiting Regulator of Actin Dynamics. |
| CTSZ | Cathepsin Z. |
| CYB5D2 | Cytochrome B5 Domain Containing 2. |
| DNASE1 | Deoxyribonuclease 1. |
| EFL1 | Elongation Factor Like GTPase 1. |
| ESYT3 | Extended Synaptotagmin 3. |
| ETFDH | Electron-Transferring-Flavoprotein Dehydrogenase. |
| FAT1 | FAT Atypical Cadherin 1. |
| FBLN2 | Fibulin 2. |
| FBXO36 | F-Box Only Protein 36. |
| FHDC1 | FH2 Domain Containing 1 (INF1). |
| FLACC1 | Flagellum-Associated Containing Coiled-Coil Domains 1. |
| FOS | Fos Proto-Oncogene, AP-1 Transcription Factor Subunit. |
| FOXD4L1 | Forkhead Box D4 Like 1. |
| GAPDHS | Glyceraldehyde-3-Phosphate Dehydrogenase, Spermatogenic. |
| GNL1 | G Protein Nucleolar 1 (Putative). |
| GREB1 | Growth Regulating Estrogen Receptor Binding 1. |
| ITGB3 | Integrin Subunit Beta 3. |
| KIF6 | Kinesin Family Member 6. |
| KIR2DL1;KIR2DL2;KIR2DL3;KIR2DS2;KIR2DS3;LOC102725023 | Killer Cell Immunoglobulin-Like Receptor, Two Ig Domains And Long Cytoplasmic Tail 1. |
| KRT12 | Keratin 12. |
| KRT38 | Keratin 38. |
| KRT6B | Keratin 6B. |
| KRT82 | Keratin 82. |
| LDHAL6B | Lactate Dehydrogenase A Like 6B. |
| METRN | Meteorin, Glial Cell Differentiation Regulator. |
| MIER2 | MIER Family Member 2. |
| MRI1 | Methylthioribose-1-Phosphate Isomerase 1. |
| MUC21 | Mucin 21, Cell Surface Associated. |
| MXRA5 | Matrix Remodeling Associated 5. |
| MYO5B | Myosin VB. |
| NAV2 | Neuron Navigator 2. |
| NDUFA10 | NADH:Ubiquinone Oxidoreductase Subunit A10. |
| NINL | Ninein Like. |
| NUAK1 | NUAK Family Kinase 1. |
| NUDT19 | Nudix Hydrolase 19. |
| OR1L6 | Olfactory Receptor Family 1 Subfamily L Member 6. |
| OR5H2 | Olfactory Receptor Family 5 Subfamily H Member 2. |
| PKD2L2 | Polycystin 2 Like 2, Transient Receptor Potential Cation Channel. |
| PNMA8A | PNMA Family Member 8A. |
| PRAMEF6 | PRAME Family Member 6. |
| PRMT9 | Protein Arginine Methyltransferase 9. |
| RGL1 | Ral Guanine Nucleotide Dissociation Stimulator Like 1. |
| RIOK3 | RIO Kinase 3. |
| RNF207 | Ring Finger Protein 207. |
| RRP12 | Ribosomal RNA Processing 12 Homolog. |
| SERPINB2 | Serpin Family B Member 2 (PAI2). |
| SFXN5 | Sideroflexin 5. |
| SHCBP1 | SHC Binding And Spindle Associated 1. |
| SIDT1 | SID1 Transmembrane Family Member 1. |
| SIGLEC6 | Sialic Acid Binding Ig Like Lectin 6. |
| SIPA1L3 | Signal Induced Proliferation Associated 1 Like 3. |
| SPATA20 | Spermatogenesis Associated 20. |
| TMEM253 | Transmembrane Protein 253. |
| TMPRSS5 | Transmembrane Serine Protease 5. |
| TRIM35 | Tripartite Motif Containing 35. |
| TRPM1 | Transient Receptor Potential Cation Channel Subfamily M. |
| TRPM4 | Transient Receptor Potential Cation Channel Subfamily M. |
| TTC38 | Tetratricopeptide Repeat Domain 38. |
| TTN | Titin. |
| TVP23C | Trans-Golgi Network Vesicle Protein 23 Homolog C. |
| TYR | Tyrosinase. |
| UCKL1 | Uridine-Cytidine Kinase 1 Like 1. |
| XDH | Xanthine Dehydrogenase. |
| ZBTB24 | Zinc Finger And BTB Domain Containing 24. |
| ZFP2 | ZFP2 Zinc Finger Protein. |
| Biological Process/Pathological Condition | Gene Name |
| Angiogenesis | CMA1, CRACD, EFL1, FBXO36, ITGB3, METRN, NDUFA10, SERPINB2 |
| Cardiogenesis | C2ORF88, CIBAR2, FOXD4L1, GNL1, MRI1, NUAK1 |
| Cardiogenesis and Angiogenesis | ANTRX1, FBLN2, FOS, MIER2, MYO5B, TRPM4, ZFP2 |
| Cardiovascular Disease | C2ORF88, COL22A1, CTSZ, ESYT3, FOS, GREB1, KIF6, MXRA5, RNF207, RRP12, SFXN5, SHCBP1, SIGLEC6, TMPRSS5, TTC38, TVP23C |
| Heart Failure | ARMT1, CPXM2, SERPINB2 |
| Cancer | ANTRX1*, C2ORF88***, COL22A1, CRACD*, CTSZ, CYB5D2***, ESYT3, ETFDH, FAT1, FBLN2, FHDC1, FOXD4L1, GREB1*, ITGB3, MUC21, MYOB5, NINL, NUAK1, RIOK3, RNF207, RRP12*, SHCBP1**, SIDT1, SIGLEC6*, SIPA1L3, SPATA20*, TMEM253, TMPRSS5**, TRPM4***, TTC38*, TVP23C, TYR, XDH |
| Heart Failure and Cancer | ABCB9, ATM, CD19, CITED4, MXRA5*, TRIM35, TRPM1**, TTN***, UCKL1** |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVascular Anomalies and Treatments · Tracheal and airway disorders · Pulmonary Hypertension Research and Treatments
Introduction
Pulmonary veins represent a unique anatomical structure because they are the only type of veins in the human body that carry oxygenated blood. The main physiological function of pulmonary veins is to provide oxygenated blood flow from both lungs to the heart [1], with an additional important function being regulation of pulmonary circulation via active vasomotion [2]. Pulmonary venous development (PVD) is a complex process that is intricately linked to cardiogenesis, the prenatal development of the heart, and commences at 27-29 days of gestation [3,4]. Normal PVD yields four pulmonary veins that drain into the left atrium, and as such, this pattern is present in 60-70% of the population [5], whereas approximately 38% of the population shows different anatomical configurations [6]. The pulmonary vein pathophysiology is mostly associated with atrial fibrillation [1], non-small cell lung cancer propagation [1], and pulmonary venous stenosis [1,3]. The latter is becoming more common as a result of more aggressive treatment of atrial fibrillation by radiofrequency ablation [1,3]. Despite the very important physiological role of pulmonary veins, there is a paucity of information regarding the molecular mechanism(s) governing their development, particularly the PVD genetic underpinnings. The information with regards to PVD genetic foundation is less scarce in the case of the total anomalous pulmonary venous connection (TAPVC), where a number of candidate genes have been identified, pointing toward TAPVC's polygenic nature [7,8]. Therefore, gaining new insights into the PVD genetic underpinnings would be highly beneficial for our understanding of such complex developmental processes and its associated pathophysiology. This task was approached in the current report by using the next-generation sequencing (NGS) platform for the whole exome sequencing (WES) genetic screening of DNA extracted postmortem from the body of an elderly individual with a remarkably high number of pulmonary veins (12 in total) with normal connections. The respective in-depth anatomical characterization was presented in our recent publication [9]. Because this individual had three major comorbidities - arteriosclerotic heart disease as well as prostate and colon cancers - the respective studies allowed not only identification of CRACD as a potential PVD gene but also pointed toward pleiotropic C2ORF88 as a hub gene in the putative angiogenesis ↔ heart failure ↔ cancer network.
Case presentation
Body donor
The body of an 87-year-old male was received through the Saint Louis University (SLU) Gift Body Program with signed informed consent from the donor. The self-reported medical history included colon cancer surgery, heart bypass surgery, and prostate cancer radiation. The donor’s causes of death were congestive heart failure and arteriosclerotic heart disease.
Genetic analysis
The postmortem genetic screening by WES on the Illumina NGS platform (San Diego, CA, USA) and the respective bioinformatics analysis were performed exactly as previously described [10]. The cumulative exome coverage for 50x depth of coverage was 94%.
Results
The anatomical examination revealed anomalous PVD in the donor, resulting in the presence of significantly more pulmonary veins than anticipated. Expected pulmonary venous anatomy dictates the presence of four pulmonary veins connecting to the left atrium, with two on the left and two on the right side. In this case, the donor had the expected two left-sided pulmonary veins, but 10 right-sided pulmonary veins (Figure 1). Literature consistently reports variations up to a total of six pulmonary veins; however, there is no documented case in the English language with 12 or more pulmonary veins. Notably, the right-sided vessels had a total cross-sectional area that was smaller than expected. This reduced cross-sectional area and subsequently increased vascular resistance may contribute to the development of pulmonary hypertension. The respective results were presented in further detail in our recent publication [9]. By investigating the underlying anatomic variations in the context of his underlying medical conditions (heart disease, prostate cancer, and colon cancer) and normal embryologic development, the genetic underpinnings can begin to be elucidated, so that clinical management may be guided. The genetic screening of the donor by WES revealed 80 rare (minor allele frequency, MAF ≤ 0.01) pathological/deleterious variants (Table 1).
Aberrant pulmonary venous anatomy observed in the present case.A: Typical pulmonary vein (PV) anatomy. B: Aberrant PV anatomy observed in the present case. PVs labeled “R” drain the right lung, and PVs labeled “L” drain the left lung. PVs outlined in blue represent those draining the upper lobe. PVs outlined in red represent those draining the middle lobe. PVs outlined in green represent those draining the lower lobe. R1 (A), representing the right superior pulmonary vein, is shaded in both blue and red to illustrate its physiologic drainage of both the upper and middle lobes of the right lung. Similarly, R2 (B) was found to drain both the upper and middle lobes of the right lung and is depicted accordingly. This diagram (B) is based on our recent publication describing the anatomy of this case [9].Image credit: The authors
Taking into account the intrinsic link between cardiogenesis and the development of new blood vessels, angiogenesis [3], as well as the presence of coexisting medical conditions in the donor such as arteriosclerotic heart disease (eventually leading to heart failure), colon and prostate cancers (see above), the genetic variants most relevant to the present case were grouped into the following categories: Angiogenesis; Cardiogenesis; Angiogenesis and Cardiogenesis; Cardiovascular Disease; Heart Failure; Cancer; Heart Failure and Cancer. The respective data are presented in Table 2.
Discussion
The results of genetic screening were consistent with a polygenic nature of aberrant PVD in the present case, meaning this condition is likely caused by a combination of multiple genetic variants. Indeed, there were multiple entries in the Angiogenesis as well as Cardiogenesis groups (Table 2) signifying an interlink between these two major developmental processes [3]. In the Angiogenesis group, there was a notable presence of the CRACD gene with the only biallelic variant (NM_020722:exon8:c.A1033G:p.R345G) in the entire dataset. CRACD, also known as CRAD, is involved in the negative regulation of barbed-end actin filament capping [11]. Because actin capping plays a key role in the regulation of cell motility [12,13] and angiogenesis [14,15], one could suggest an important role for CRACD in the observed aberrant PVD. The other interesting gene, C2ORF88, was present in three groups: Cardiogenesis, Cardiovascular Disease, and Cancer. This gene was differentially regulated in the bicuspid aortic valve with leaflet redundancy [16] as well as in colon cancer [17]. Yet, C2ORF88 was also related to atrial fibrillation [18] and prostate cancer [19]. Since atrial fibrillation could lead to heart failure [20], C2ORF88 could serve as an important player in the heart failure ↔ colorectal/prostate cancer axis, apparently present in the current case. Intriguingly, there was also a number of genes linked to both angiogenesis and cardiogenesis (Table 2), which could point to a much closer link between these two biological processes, potentially regulated by pleiotropic genes.
The data obtained in the current study could also provide novel insights into the molecular basis of an important link between heart failure and cancer [21,22] due to common risk factors for these pathological conditions [23]. The genetic screening of the donor who had colon cancer, prostate cancer, and succumbed to heart failure identified three genes linked to the latter - ARMT1, CPXM2, and SERPINB2 - along with a plethora of genes associated with cancer, including those of colorectal and prostate (Table 2). Interestingly, in addition to genes linked to colorectal or prostate cancer, there were also two genes known to be associated with both of those cancer types: CYB5D2 [24,25] and TRPM4 [26,27]. More interestingly, there was also a group of pleiotropic genes - ABCB9, ATM, CD19, CITED4, MXRA5, TRIM35, TRPM1, TTN, UCKL1 - linked to both heart failure and cancer. Within these pleiotropic genes, MXR5 [28,29] was associated with colorectal cancer [30]; TRPM1 [31] and UCKL1 [32,33] were linked to prostate cancer [34,35], and TTN [36] was associated with both cancer types [37].
Despite providing important insights into the putative mechanism(s) underlining a pulmonary venous malformation, the current case involved one individual who was studied postmortem. Therefore, the current report can only be viewed as an initial step for the antemortem studies in a clinical setting with a large cohort of patients having similar malformations and/or using respective clinical database(s).
Conclusions
The profound pulmonary venous malformation observed in the donor had most likely pleiotropic genetic underpinnings with a potential involvement of an aberrant barbed-end actin filament capping due to the rare biallelic mutation in the CRACD gene. The pleiotropic C2ORF88 gene could possibly serve as a hub gene linking aberrant angiogenesis, heart failure, and cancer, all present in the donor. These results warrant a study of similar cases using a large clinical database(s).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Learning from the pulmonary veins Radiographics Porres DV Morenza OP Pallisa E Roque A Andreu J Martínez M 99910223320132384296910.1148/rg.334125043 · doi ↗ · pubmed ↗
- 2Role of veins in regulation of pulmonary circulation Am J Physiol Lung Cell Mol Physiol Gao Y Raj JU 026288200510.1152/ajplung.00103.200415640520 · doi ↗ · pubmed ↗
- 3Congenital and acquired pulmonary vein stenosis Circulation Latson LA Prieto LR 10310811520071720045310.1161/CIRCULATIONAHA.106.646166 · doi ↗ · pubmed ↗
- 4Development of the pulmonary vein and the systemic venous sinus: an interactive 3D overview P Lo S One van den Berg G Moorman AF 06201110.1371/journal.pone.0022055 PMC 313362021779373 · doi ↗ · pubmed ↗
- 5CT of the pulmonary veins J Thorac Imaging Lacomis JM Goitein O Deible C Schwartzman D 63762220071732557810.1097/RTI.0b 013e 3180317 aaf · doi ↗ · pubmed ↗
- 6Pulmonary vein anatomy in patients undergoing catheter ablation of atrial fibrillation: lessons learned by use of magnetic resonance imaging Circulation Kato R Lickfett L Meininger G 2004201010720031268199410.1161/01.CIR.0000061951.81767.4E · doi ↗ · pubmed ↗
- 7Next-generation sequencing identifies novel genes with rare variants in total anomalous pulmonary venous connection E Bio Medicine Shi X Huang T Wang J 2172273820183044822510.1016/j.ebiom.2018.11.008PMC 6306349 · doi ↗ · pubmed ↗
- 8Research progress in pathogenesis of total anomalous pulmonary venous connection Methods Mol Biol Shi X Lu Y Sun K 173178220420203271032410.1007/978-1-0716-0904-0_15 · doi ↗ · pubmed ↗