Rolling Leaf 2 Controls Leaf Rolling by Regulating Adaxial-Side Bulliform Cell Number and Size in Rice
Yu-Jia Leng, Shi-Yu Qiang, Wen-Yu Zhou, Shuai Lu, Tao Tao, Hao-Cheng Zhang, Wen-Xiang Cui, Ya-Fan Zheng, Hong-Bo Liu, Qing-Qing Yang, Ming-Qiu Zhang, Zhi-Di Yang, Fu-Xiang Xu, Hai-Dong Huan, Xu Wei, Xiu-Ling Cai, Su-Kui Jin, Ji-Ping Gao

TL;DR
This study identifies a gene, RLL2, that controls leaf rolling in rice by affecting bulliform cell size and number, which could help improve rice plant architecture and yield.
Contribution
The discovery of RLL2, a calpain-like cysteine proteinase gene regulating leaf rolling through bulliform cell changes in rice.
Findings
The rll2 mutant shows increased leaf rolling due to larger and more numerous bulliform cells.
RLL2 is a recessive nuclear gene encoding a plant-specific calpain-like cysteine proteinase.
Transcriptome analysis revealed 104 differentially expressed genes, including altered transcription factors in the mutant.
Abstract
Leaves represent an important organ in plant photosynthesis, and moderately rolled leaves would be beneficial in establishing an ideal plant architecture and thereby increasing rice yields. In this study, a stable inherited rolled leaf mutant was obtained via ethyl methanesulfonate (EMS) mutagenesis from japonica variety WYJ27, which was named rll2 (rolling leaf 2). rll2 showed a leaf-rolling phenotype at the seedling stage, which increased with growth. Compared with the wild type, the leaves at all levels of rll2 were significantly shorter and narrower, and the leaf-rolling index gradually decreased from the highest leaf to the third-highest leaf. Semi-thin sections showed that the bulliform cells of rll2 were significantly larger than those of the wild type, and the number of cells was significantly higher than that of the wild type. Genetic analysis showed that rll2 is controlled by…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
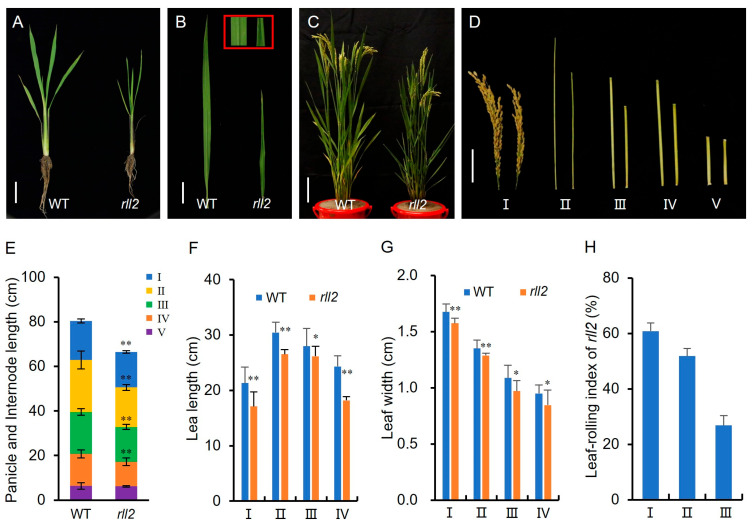 Figure 1
Figure 1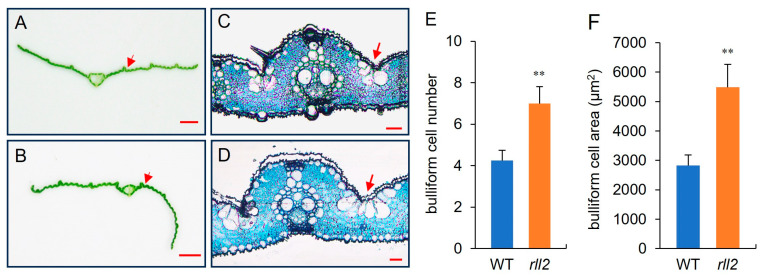 Figure 2
Figure 2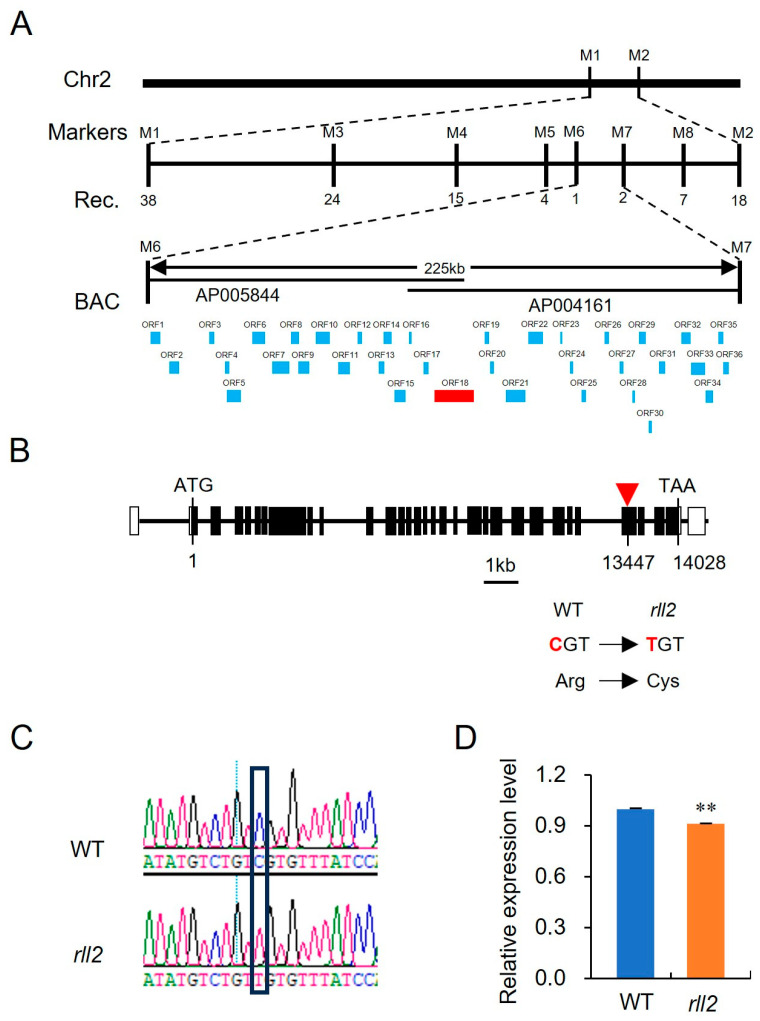 Figure 3
Figure 3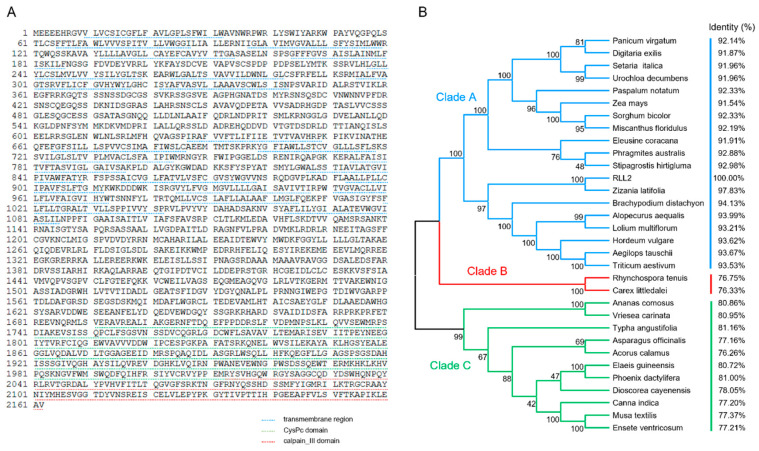 Figure 4
Figure 4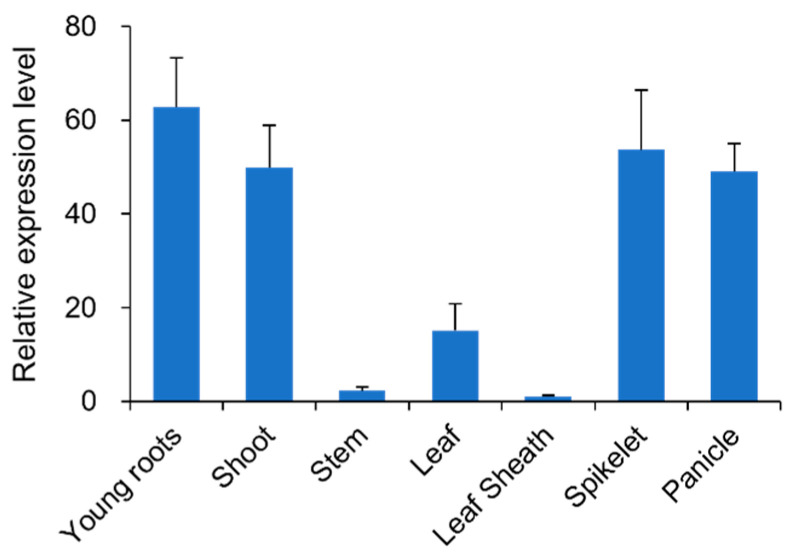 Figure 5
Figure 5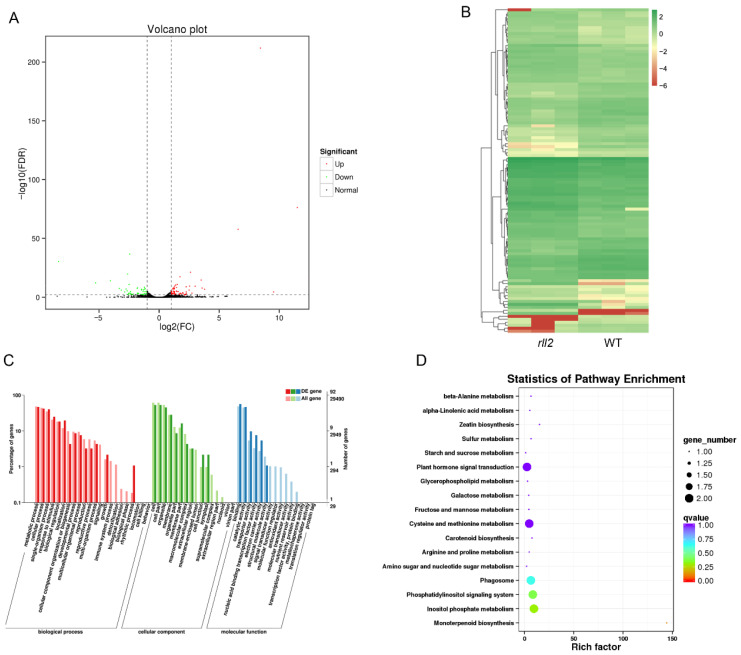 Figure 6
Figure 6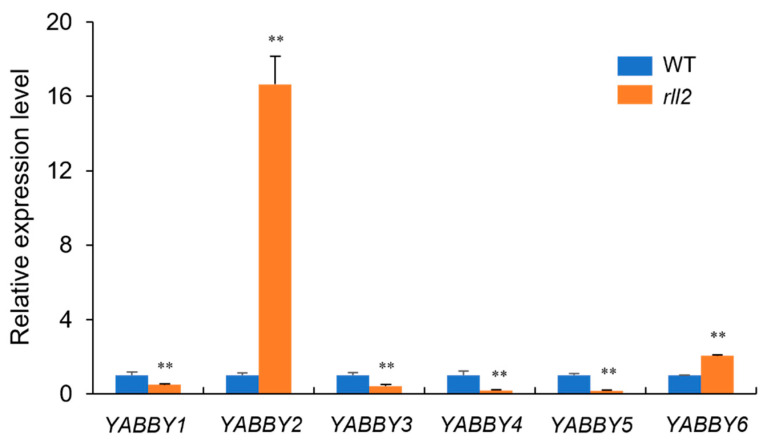 Figure 7
Figure 7- —Jiangsu Province Government
- —Natural Science Foundation of the Jiangsu Higher Education Institutions of China
- —Jiangsu Key Laboratory of Crop Genomics and Molecular Breeding
- —Priority Academic Program Development of Jiangsu Higher Education Institutions
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPolysaccharides and Plant Cell Walls · Plant Molecular Biology Research · Plant Reproductive Biology
1. Introduction
Leaves represent the main site of photosynthesis in plants and provide energy for plant growth. As an important part of plant architecture, leaf morphology is closely related to rice yields [1]. Moderate leaf rolling not only contributes to improving light acceptance, delaying leaf senescence, and accelerating the accumulation of dry matter but also helps to improve root system activity and strengthen lodging resistance [2,3]. Therefore, the study of leaf rolling has high practical value in high-yield rice breeding.
Leaf development is a complex process comprising cell division and expansion, axis determination, and tissue differentiation and specification [4]. In rice, leaf development involves three stages: the initiation of leaf primordia, polarity establishment and maintenance, and leaf flattening and intercalary growth [5,6]. The leaf primordium originates from the peripheral zone of the apical shoot meristem (SAM) [7], develops along the adaxial–abaxial, medial–lateral, and apical–basal polarity axes, and finally forms mature leaves [8,9]. This developmental process is regulated by complex regulatory networks involving numerous functional genes, transcription factors, phytohormones, and microRNAs [1,3]. To date, many genes controlling leaf rolling have been identified in rice, such as SHALLOT-LIKE1/ROLLED LEAF 9 (SLL1/RL9) [10,11], SHALLOT-LIKE1 (SLL2) [12], SEMI-ROLLED LEAF 1 (SRL1) [13,14], SEMI-ROLLED LEAF 1 (SRL2) [2], ABAXIALLY CURLED LEAF 1 (ACL1) [15], ABAXIALLY CURLED LEAF 2 (ACL2) [15], NARROW AND ROLLED LEAF 1 (NRL1) [16], NARROW LEAF 7 (NAL7) [17], Rolling-leaf14 (RL14) [18], ADAXIALIZED LEAF 1 (ADL1) [19], ARGONAUTE 1b (AGO1b) [20], Rolled and Erect Leaf 1 (REL1) [21], lateral organ boundaries domain (LBD) gene LBD3-7 [22], zinc finger homeodomain class homeobox gene OsZHD1 [23], class IV homeodomain gene CFL1 [24], class III HD-Zip gene OsHOX32 [25], RICE OUTERMOST CELLSPECIFIC GENE (Roc) family gene Roc5 [26], and Roc8 [27]. Most of these genes regulate leaf rolling by changing the morphology of leaf bulliform cells, sclerenchyma cells, cells in vascular bundles, cuticle, epidermal cells, and mesophyll cells.
Bulliform cells, also called hygroscopic cells or motor cells, are among the most important cell structures and are found on the adaxial epidermis of the leaf. A change in osmotic pressure in bulliform cells can cause leaf rolling. Advances in research have suggested that leaf rolling is primarily caused by bulliform cell changes in rice [21]. For example, SLL1 encodes a SHAQKYF-class MYB family transcription factor. The sll1 mutant shows a leaf-rolling phenotype due to the formation of bulliform cells on the abaxial epidermis [10]. SRL1 encodes a putative glycosylphosphatidylinositol-anchored protein, which causes adaxial rolling in leaves by inhibiting the formation of bulliform cells [13]. REL1 encodes a high-degree conservation and unknown-function protein in monocot plants. The rel1 mutant shows a leaf rolling due to an increase in the size and number of bulliform cells [21]. Although many genes relating to leaf rolling have been identified, the molecular mechanisms behind leaf rolling in rice are not yet fully understood. In the present study, we identified and characterized a rolled-leaf mutant, named rolling leaf 2 (rll2), from a japonica variety Wuyunjing27 (WYJ27) EMS mutant library. Using a map-based cloning method, we cloned the RLL2 gene and found that it encodes a conserved and plant-specific calpain-like cysteine proteinase, allelic to the ADL1 gene. The loss of RLL2 function leads to an increase in the number and size of bulliform cells on the adaxial side of the leaf, resulting in abaxially rolled leaves. This study enhances our understanding of the role of RLL2/ADL1 in leaf rolling.
2. Materials and Methods
2.1. Plant Materials
The rice rll2 mutant was obtained from ethyl methanesulfonate-treated (EMS) japonica cultivar Wuyunjing27 (WYJ27). An F_2_ segregation population was obtained by crossing rll2 with an indica cultivar IR36. WYJ27, IR36, rll2, and F_2_ segregation populations were germinated and sown on Yangzhou University’s experimental farms in Lingshui (110°00′ E, 18°31′ N) and Yangzhou (119′ 42 E, 32°39′ N) during the rice-growing season. After 30 days, the seedlings were transplanted into a field at a density of 18 cm × 18 cm. About 150 kg N ha^−1^ (as urea), 130 kg K_2_O ha^−1^, and 60 kg P_2_O_5_ ha^−1^ were added to each field. Field management, including irrigation and pest control, followed normal agricultural practices.
2.2. Measurement of Leaf-Rolling Index and Agronomic Traits
The leaf-rolling index (LRI) was calculated using the following formula: LRI = (Lw − Ln)/Lw × 100% [28]. Lw was defined as the largest leaf width in the fully expanded leaves; Ln was defined as the natural distance of the leaf margins. The plant height (PH), tiller number (TN), primary branch number (PBN), secondary branch number (SBN), and grain number per panicle (GNPP) were measured at 25 days after heading. The plant height was measured from the base to the tip of the tallest panicle. The panicle at the highest point on the plant was selected to investigate the grain number. Except for two marginal plants on each side, twenty independent plants were randomly selected to score the phenotypic data. The rice grains were harvested at 35 days after flowering, dried in the sun, and then transferred to a room-temperature environment. Once the physical and chemical properties were stable, the grain shape traits, including grain length (GL), grain width (GW), and thousand-grain weight (TGW), were determined using the WSeen SC-G automatic seed testing system and thousand-grain weight analysis system (Hangzhou WSeen Detection Technology Co., Ltd., Hangzhou, China). About 150–200 grains were evaluated. Grain thickness (GT) was measured using a digital vernier caliper. A total of ten grains were used to measure GT.
2.3. Histology and Microscopic Observation
Hand-cut sections of WT and rll2 leaves (about 35 days after sowing) were observed on a slide with an OLYMPUS SZX16 stereomicroscope (Olympus Corporation, Tokyo, Japan). To observe leaf anatomy in detail, young leaves were fixed in a formalin–acetic acid–alcohol (FAA) solution (50% ethanol, 5% formaldehyde, 5% glacial acetic acid = 18:1:1 v:v:v) at 4 °C overnight. After dehydration in serial concentrations of ethanol (50%, 70%, 90%, 100%, 100%), they were infiltrated and embedded in Technovit 7100 resin (Heraeus Kulzer, Hanau, Germany). Sectioning (3 μm) was conducted with the Leica RM2265 (Leica, Wetzlar, Germany). Slices were spread on a platform at 42 °C and stained using toluidine blue at room temperature for 15 min. Observations and photography were carried out using the Leica DM2500 LED light microscope (Leica, Wetzlar, Germany).
2.4. Genetic Analysis and Map-Based Cloning of RLL2
An F_2_ mapping population was derived from a cross between rll2 and IR36. A total of 859 individuals with the mutant phenotype were used for fine mapping. The rll2 loucs was first mapped to an interval between markers M1 and M2 on chromosome 2. Further, it was narrowed to a 225 kb region between maker M6 and M7. The molecular markers were developed with Primer Premier 5 software (Premier Biosoft International, Palo Alto, CA, USA) based on sequence differences between the japonica variety Nipponbare and the indica variety 93-11. The molecular markers used in this study are listed in Table S1. Gene prediction and sequence analysis were performed using the Rice Genome Annotation Project (http://rice.uga.edu/index.shtml (accessed on 3 September 2023)) databases. For the identification of the candidate gene, the corresponding DNA fragments were amplified from WYJ27 and rll2 using Kod FX (TOYOBO, Osaka, Japan) and sequencing using Applied Biosystems 3730xl DNA Analyzers (Applied Biosystems, Carlsbad, CA, USA). The primers for sequencing in this study are listed in Table S2.
2.5. Phylogenetic Analysis
The amino acid sequences of the RLL2 homological proteins were retrieved by searching the NCBI BLAST database (https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 10 June 2024)) using the total amino acid sequences of RLL2. Phylogenetic trees were constructed with the aligned protein sequences using MEGA 12 software based on the neighbor-joining statistical method with the following parameters: Poisson model, uniform rate, pairwise deletion, and bootstrap (1000 replicates) [29].
2.6. RNA Extraction and qRT-PCR
The total RNA of various tissues in WYJ27 and rll2 was extracted using the RNAprep Pure Plant kit (TIANGEN, Beijing, China), and reverse transcription into cDNA was conducted with the PrimeScript™ RT reagent kit with gDNA Eraser (TaKaRa, Shiga, Japan) according to the manufacturer’s instructions. The cDNA was used for real-time RT-PCR using the TB Green^®^ Premix EX Taq^TM^ kit (TaKaRa, Shiga, Japan) and gene-specific primers (Table S3) with a CFX Connect^TM^ Real-Time System (Bio-Rad Laboratories, Hercules, CA, USA). The rice ACTIN gene was used as an internal standard to normalize the expression level of RLL2 and other tested genes. The relative expression levels were calculated based on the comparative Ct method using the 2^−∆∆Ct^ formula [30].
2.7. RNA-Seq Analysis
Twenty days after sowing, the flat leaves of wild-type WYJ27 and the rolled leaves of the mutant rll2 were selected for transcriptome sequencing, and each sample was pooled for total RNA isolation with three biological replicates. Total RNA was extracted using the RNAprep Pure Plant kit (TIANGEN, Beijing, China) according to the manufacturer’s protocol. Transcriptome sequencing was performed by Biomarker Technologies Corporation using the Illumina HiSeq2500 system (Illumina, San Diego, CA, USA). Transcriptome assembly was performed according to the protocol described previously [31]. In brief, the raw RNA-Seq data were processed using in-house Perl scripts to trim adapters and low-quality sequences and then assembled using Trinity based on left.fq and right.fq [32]. Differential expression analysis was performed using the DESeq R package (version 1.40.0). The fold change (FC) ≥ 2 and false discovery rate (FDR) < 0.01 were used as the screening criteria. Gene function was annotated based on the KEGG (Kyoto Encyclopedia of Genes and Genomes) and GO (Gene Ontology) databases. GO and KEGG pathway enrichment analysis was performed using BMKCloud (www.biocloud.net).
2.8. Statistical Analysis
Data are presented as the mean ± standard deviations of multiple biological replicates (n ≥ 3). Significant differences were assessed using Student’s t test. p < 0.05 and p < 0.01 were defined as significant and highly significant, respectively.
3. Results
3.1. Phenotypic Characterization of the rll2 Mutant
The rll2 mutant was identified by screening an ethyl methanesulfonate (EMS) mutant library for the ‘Wuyunjing 27’ (WYJ27) background and had abaxially rolled leaves throughout the entire growth period (Figure 1A–C). The leaf-rolling index (LRI) of rll2 at the heading stage from the highest (flag leaf) to the third-highest leaf was 60.9%, 51.9%, and 26.9%, respectively, whereas wild-type WYJ27 leaves were nearly flat (Figure 1H). rll2 also displayed short and narrow leaves; the length and width of the highest, second-highest, third-highest, and fourth-highest leaves in rll2 were 80.1%, 87.1%, 93.6%, and 74.8% and 94.0%, 94.5%, 89.2%, and 89.0% of those for the wild type, respectively (Figure 1F,G). Furthermore, the yield traits including plant height, internode length, tillering number, primary branch number, secondary branch number, grain number per panicle, grain length, grain width, and thousand-grain weight in rll2 were significantly lower than those of the wild type (Figure 1D,E; Table 1). Taken together, these results indicate that RLL2 has pleiotropic effects in rice growth and development.
3.2. Bulliform Cell Number and Size Are Increased in rll2
To investigate the cause of abaxial leaf rolling in rll2, we analyzed cross-sections of leaves f at 35 days after sowing (Figure 2A–D). In contrast to the wild type, more bulliform cells occurred between two vascular bundle ridges in the adaxially leaf. The bulliform cell number for rll2 was 7.0 ± 0.8 cells, which was significantly higher than that of the wild type (4.3 ± 0.5) (Figure 2E). Furthermore, the bulliform cell area of rll2 was also significantly higher than that of the wild type (Figure 2F). Those results suggest that the abaxial leaf rolling in rll2 may be caused by an increase in bulliform cell number and size on the adaxial surface of leaves.
3.3. Map-Based Cloning of RLL2
To isolate RLL2 via map-based cloning, we crossed the rll2 mutant with an indica cultivar, IR36. All the F_1_ hybrids exhibited a WT morphological phenotype. In the F_2_ segregating populations, the WT morphological phenotype and rolled-leaf phenotype showed a typical segregation ratio of 3:1 (Table 2). This result indicates that rll2 is controlled by a single recessive nuclear gene. The RLL2 locus was preliminarily mapped to the chromosome 2 between markers M1 and M2 (Figure 3A). By using a large population of 859 homozygous mutant individuals, RLL2 was narrowed down to a 225 kb genomic region between markers M6 and M7 on BAC clone AP004161 (Figure 3A). According to the annotations of the rice genome database (Rice Genome Annotation Project, https://rice.uga.edu/), this region comprises twenty putative genes with annotated functions. A known gene controlling leaf rolling, ADL1 (LOC_Os02g47970), was identified in these ORFs. The adl1 mutant showed abaxially rolled leaves due to an increase in the size and number of bulliform cells. The rll2 mutant displayed a similar phenotype with adl1. DNA sequencing analysis of this ORF revealed the presence of a single nucleotide substitution (C→T) on the 27th exon of LOC_Os02g47970 in rll2 compared to the wild type, which resulted in an Arg-to-Cys substitution (Figure 3B,C). Moreover, the expression level of LOC_Os02g47970 in rll2 was significantly decreased (Figure 3D). We therefore inferred that ADL1 was the gene controlling the rll2 mutant phenotype.
3.4. RLL2 Encodes a Conserved and Plant-Specific Protein
Sequence analysis indicated that RLL2 cDNA is 6489 bp in length and encodes a protein of 2162 amino acid residues, including 22 transmembrane regions, 1 CysPc domain, and 1 calpain_III domain (Figure 4A). We performed a BLASTP search against the NCBI non-redundant protein database, revealing 31 proteins from other plant species with significant homology (≥70% identity at the amino acid level) to RLL2, which suggests that RLL2 is a plant-specific protein and shares conserved biochemical functions. To elucidate the evolutionary relationships among RLL2 proteins, a phylogenetic tree was constructed using the neighbor-joining method in MEGA 12 [29]. The 32 proteins were classified into three clades (Figure 4B). Clade A belongs to Gramineae and shows high identity (>90%) with the RLL2 protein. The identity of clade B and clade C with the RLL2 protein was lower than that of clade A, which ranged from 76.26% to 81.16%. This result indicates that the RLL2 protein is very conserved in evolution.
3.5. Expression Patterns of RLL2
To determine the expression patterns of RLL2 in rice, quantitative real-time RT-PCR (qRT-PCR) was used to detect the expression levels of RLL2 in WYJ27 issues, such as young roots, shoots, stems, leaves, leaf sheaths, spikelets, and panicles. The results showed that RLL2 was expressed in all tissues, with high expression in young roots, shoots, spikelet and panicles and low expression in stems and leaf sheaths (Figure 5).
3.6. GO and KEGG Analyses of Rolling Leaf Regulation via RLL2
To further unravel the function of RLL2 in rice, leaves from wild-type and rll2 plants at 20 d were used for transcriptome analysis. A total of 104 differentially expressed genes (DEGs) were found; 42 genes were up-regulated and 62 were down-regulated in rll2 plants (Figure 6A,B; Table S5). Among these genes, 12 transcription factor genes were found, 10 of which have been reported. We measured the expression levels of these genes using qRT-PCR, and the results showed that the expression levels of three genes (OsbHLH056, OsDREB1B, and OsWRKY71) were significantly increased and those of seven genes (OsDREB1A, OsAP2-39, PCF2, OsERF74, OsNAC5, OsNAC19, and LOC_Os05g50340) were significantly reduced in rll2 leaves (Figure S1; Table S5).
The GO enrichment analysis of DEGs is shown in Figure 6C and Table S6. In the cellular component analysis, DEGs were found to mainly be involved in the cell, cell part, organelle, membrane, and membrane part. In the molecular function analysis, DEGs were found to mainly be involved in binding, catalytic activity, transporter activity, and nucleic acid binding transcription factor activity. In the biological process analysis, DEGs were found to mainly be involved in the metabolic process, cellular process, single-organism process, response to stimulus, localization, and biological regulation. KEGG pathway enrichment analysis showed that the DEGs were mainly concentrated in plant hormone signal transduction, cysteine and methionine metabolism, phagosome, inositol phosphate metabolism, and the phosphatidylinositol signaling system (Figure 6D).
3.7. RLL2 Affects the Expression Levels of Leaf Development-Related Genes
Leaf development is regulated by several genes, such as YABBY genes [33,34,35,36]. We measured the expression levels of YABBY genes in young leaves via qRT-PCR. Compared with the wild type, YABBY2 and YABBY6 were significantly up-regulated in rll2, while YABBY1, YABBY3, YABBY4, and YABBY5 were significantly down-regulated in rll2 (Figure 7). This result suggests that rll2 phenotypes may be related to altered transcriptional activity of YABBY genes.
4. Discussion
4.1. RLL2 Plays an Important Role in Regulating the Size and Number of Leaf Bulliform Cells in Rice
Bulliform cells (motor cells), which are distributed on the upper epidermis of leaves, are the most important cell structures in leaves [27]. Bulliform cells are strongly linked to leaf-rolling phenotypes in rice [37]. Many studies have shown that alternations to bulliform cells arranged on the adaxial surface of the leaf lead to adaxial or abaxial rolling in mature leaves [21]. To date, over 10 genes controlling bulliform cell development have been reported in rice; ACL1 [15], ACL2 [15], NRL1 [16], NAL7 [17], and RL14 [18] positively regulate the development of bulliform cells, while SRL1 [13,14], ADL1 [19], OsLBD3-7 [22], OsZHD1 [23], and Roc5 [26] negatively regulate the development of bulliform cells. In this study, we identified RLL2 as a key regulator of leaf rolling. Compared to the wild type, rll2 showed a higher number and size of bulliform cells, leading to abaxial leaf rolling (Figure 2). Our results strongly suggest that RLL2 plays a negative role in the formation of bulliform cells on the adaxial leaf surface.
4.2. RLL2 Is a New Allelic Variant of ADL1
Leaves are the main photosynthetic organs in plants. Moderate leaf rolling is considered to be an important part of the ideal rice phenotype, the development of which is of great significance in improving photosynthetic efficiency and grain yields [2,10,38]. However, the molecular mechanisms underlying leaf rolling remain unclear. In this study, we identified the rll2 mutant from an EMS-mutagenized japonica variety, WYJ27 (Figure 1). Using a map-based cloning method, gene sequencing, and qRT-PCR, we found that the candidate gene is ADL1/LOC_Os02g47970, which encodes calpain-like cysteine proteinase (Figure 3 and Figure 4). Hibara et al. (2009) reported seven ADL1 allelic mutants (adl1-1, adl1-2, adl1-3, adl1-s1, adl1-s2, adl1-g1, and adl1-g2), which produced a base mutation in exon 30, 26, 4, 26, 19, 27, and 20, respectively, resulting in changes in the amino acids encoded [19]. In our study, we identified an RLL2 mutation on the 27th exon, with a C to T change (Figure 3B,C), indicating that RLL2 is a new allelic mutant of ADL1 [19]. The rll2 mutant showed a rolling leaf phenotype similar to that of the adl1 mutant, but there were some phenotypic differences among the rll2/adl1 mutant alleles; for example, the adl1 mutant showed bulliform cells on the abaxial side of leaves, while the mutant rll2 did not. adl1-1, adl1-2, and adl1-3 showed defects in leaf polarity and leaf primordium insertion in the SAM. adl1-s1 and adl1-s2 produced shootless embryos. The embryos of adl1-g1 and adl1-g2 stopped growing at the globular stage, and the endosperm lacked a ventral aleurone layer [19]. This indicates that different mutant RLL2/ADL1 alleles have different effects on rice development. In addition, KEGG pathway enrichment analysis showed that cysteine and methionine metabolism genes differed between the wild type and rll2 (Figure 6D). These changes may impact the function of calpain-like cysteine proteinase in rll2. Further investigation is needed to fully understand the role of RLL2 in rice.
4.3. Transcription Factor Genes Participate in the Development of Leaves in RLL2
Transcription factors (TFs) are a group of DNA-binding proteins that regulate gene expression and are involved in a diverse range of biological processes in plants [39]. In rice, several transcription factors have been reported as being involved in the regulation of leaf rolling; For example, SLL1 encodes an SHAQKYF-class MYB transcription factor belonging to the KANADI family. It regulates leaf rolling in rice by regulating the development of cells on the abaxial side of the leaf [10]. Roc8 is a member of the plant-specific homeodomain leucine zipper IV (HD-Zip IV) gene family, encoding a class of homeobox transcription factors. The overexpression of Roc8 causes leaf rolling on the adaxial side, and its knockout causes leaf rolling on the abaxial side [27]. OsMYB103L encodes an R2R3-MYB transcription factor. The overexpression of OsMYB103L leads to a rolled-leaf phenotype. Further analysis shows that OsMYB103L regulates cellulose synthesis by targeting CESA genes, thereby affecting leaf rolling [40]. In our study, the expression levels of 10 transcription factor genes changed in 104 DEGs, indicating that transcription factors play an important role in leaf rolling (Figure S1; Table S5).
YABBY genes are plant-specific transcription factors that play an important role in the regulation of a diverse range of developmental processes, such as the establishment of adaxial–abaxial polarity, laminal expansion, and floral organ development [36,41,42]. Many YABBY genes have been studied in monocotyledon plants, and several YABBY genes have been cloned in rice. OsYABBY1 shows homology with AtYAB2 and AtYAB5, is expressed in sclerenchyma, and participates in the development of the stamen, carpel, and meristem [43]. YAB3 is closely related to maize ZmYAB14 and Arabidopsis FILAMENTOUS FLOWER (FIL), which is mainly related to the development of leaves [33]. OsYABBY4 is involved in the development of the vascular structure [44]. OsYABBY5 (TOB1) plays a vital role in promoting lateral organ development and maintaining meristem organization in the rice spikelet [36]. DROOPING LEAF (DL), a rice orthologue of CRC, is mainly related to carpel specification and midvein development in rice [45]. In our study, the expression level of YABBY genes in the rll2 mutant was altered. YABBY2 and YABBY6 were significantly up-regulated in rll2, while YABBY1, YABBY3, YABBY4, and YABBY5 were significantly down-regulated. In particular, the expression of YABBY2 was significantly up-regulated (16-fold) in rll2 (Figure 7). This suggests that the rll2 phenotype may be related to altered transcriptional activity in YABBY genes.
Rice leaf morphology is an important component in achieving the ideal plant architecture and significantly impacts rice yields [1]. Using mutagenesis techniques to identify mutants with moderately rolled leaves is beneficial in breeding crops with the desired architecture [2]. Although our study shows that RLL2 plays an important role in regulating leaf rolling, more research is needed on, for example, the effect of RLL2 gene mutation on calpain-like cysteine proteinase activity, how transcription factors genes affect the development of bulliform cells by regulating RLL2, and whether certain RLL2 alleles are preponderant in the improvement of rice leaf morphology. These questions require further study.
5. Conclusions
The rolling leaf 2 (rll2) mutant was obtained through the ethyl methanesulfonate (EMS) mutagenesis of the japonica variety WYJ27. The rll2 mutant displayed abaxially rolled leaves throughout the entire growth period. We demonstrated that rll2 is a new allele of ADL1, which encodes a CysPc domain and calpain_III domain protein. We suggest that the increase in the number and size of bulliform cells is causative of leaf rolling in the rll2 mutant. Moreover, qRT-PCR showed that the expression of some transcription factor genes changed significantly in rll2. Our study provides a novel allelic mutant to explore the function of RLL2. At the same time, it provides useful information for furthering the improvement of rice architecture.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gong X. Chen J. Chen Y. He Y. Jiang D. Advancements in Rice Leaf Development Research Plants 20241390410.3390/plants 1306090438592944 PMC 10976080 · doi ↗ · pubmed ↗
- 2Liu X. Li M. Liu K. Tang D. Sun M. Li Y. Shen Y. Du G. Cheng Z. Semi-Rolled Leaf 2 modulates rice leaf rolling by regulating abaxial side cell differentiation J. Exp. Bot.2016672139215010.1093/jxb/erw 02926873975 PMC 4809286 · doi ↗ · pubmed ↗
- 3Wang J. Xu J. Wang L. Zhou M. Nian J. Chen M. Lu X. Liu X. Wang Z. Cen J. SEMI-ROLLED LEAF 10 stabilizes catalase isozyme B to regulate leaf morphology and thermotolerance in rice (Oryza sativa L.)Plant Biotechnol. J.20232181983810.1111/pbi.1399936597711 PMC 10037157 · doi ↗ · pubmed ↗
- 4Moulia B. Leaves as shell structures: Double curvature, auto stresses, and minimal mechanical energy constraints on leaf rolling in grasses J. Plant Growth Regul.200019193010.1007/s 00344000000411010989 · doi ↗ · pubmed ↗
- 5Wang H. Kong F. Zhou C. From genes to networks: The genetic control of leaf development J. Integr. Plant Biol.2021631181119610.1111/jipb.1308433615731 · doi ↗ · pubmed ↗
- 6Du F. Guan C. Jiao Y. Molecular mechanisms of leaf morphogenesis Mol. Plant 2018111117113410.1016/j.molp.2018.06.00629960106 · doi ↗ · pubmed ↗
- 7Shen W. Sun J. Xiao Z. Feng P. Zhang T. He G. Sang X. Narrow and stripe leaf 2 regulates leaf width by modulating cell cycle progression in rice Rice 202316203110.1186/s 12284-023-00634-337071312 PMC 10113404 · doi ↗ · pubmed ↗
- 8Hasson A. Blein T. Laufs P. Leaving the meristem behind: The genetic and molecular control of leaf patterning and morphogenesis Comptes Rendus Biol.201033335036010.1016/j.crvi.2010.01.01320371110 · doi ↗ · pubmed ↗