Functional Analysis of Hyaluronidase-like Genes in Ovarian Development of Macrobrachium nipponense and Comparative Evaluation with Other Key Regulatory Genes
Zhiming Wang, Hao Dong, Hui Qiao, Wenyi Zhang, Shubo Jin, Yiwei Xiong, Zhenghao Ye, Yan Gong, Sufei Jiang, Hongtuo Fu

TL;DR
This study explores the role of two hyaluronidase-like genes in the ovarian development of a freshwater shrimp and compares their regulatory effects with other genes.
Contribution
The study identifies Mn-HyaL genes as negative regulators of ovarian maturation in Macrobrachium nipponense through RNA interference and comparative gene analysis.
Findings
Knockdown of Mn-HyaL2 significantly accelerated ovarian development in M. nipponense.
Mn-HyaL genes function as negative regulators of ovarian maturation.
Mn-Gonad-inhibiting hormone was the strongest inhibitor of ovarian development.
Abstract
This study conducted a bioinformatic analysis of two Hyaluronidase-like isoforms (Mn-HyaL1 and Mn-HyaL2) in Macrobrachium nipponense and investigated their phylogenetic relationships. The open reading frames of Mn-HyaL1 and Mn-HyaL2 were 1101 bp (encoding 366 amino acids) and 1164 bp (encoding 387 amino acids), respectively. Both isoforms exhibited similar conserved domains, with an amino acid sequence similarity of 60.21%. Quantitative PCR analysis revealed that the expression levels of Mn-HyaL1 and Mn-HyaL2 increased during the mid-to-late phase of each developmental stage, were higher during the reproductive season than in the non-reproductive season, and were more abundant in the hepatopancreas than in other tissues. RNA interference experiments targeting both genes simultaneously demonstrated that knockdown of Mn-HyaL2 significantly accelerated ovarian development in M. nipponense,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1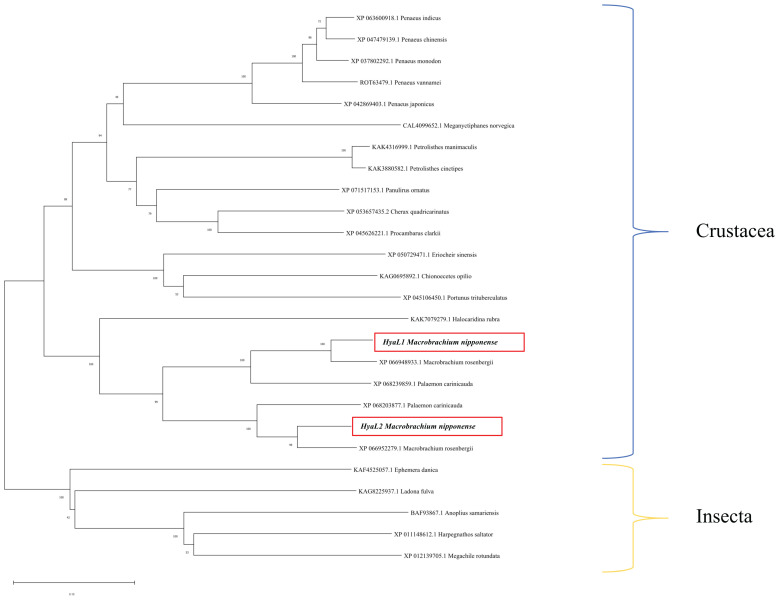 Figure 2
Figure 2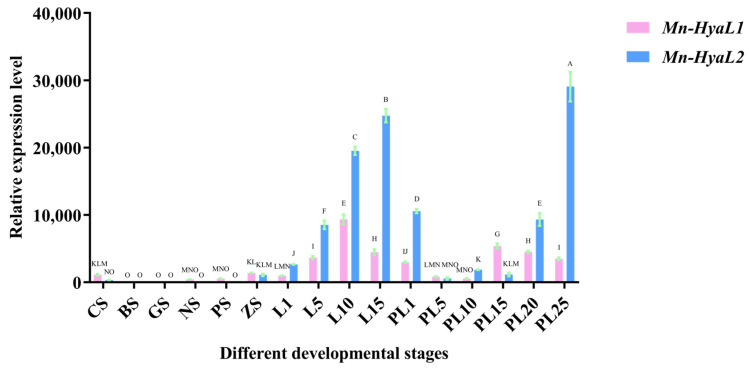 Figure 3
Figure 3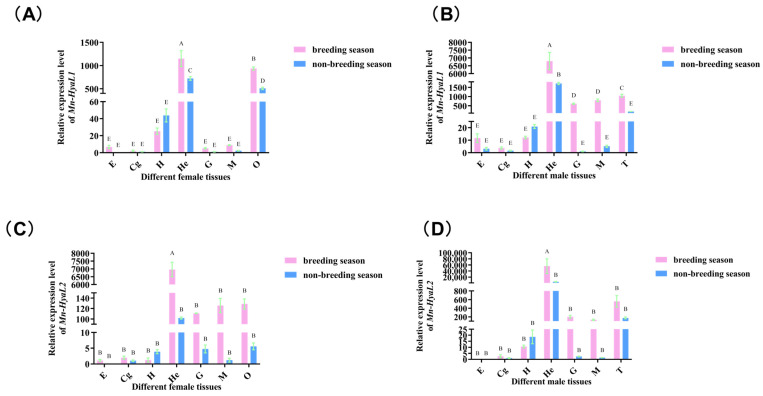 Figure 4
Figure 4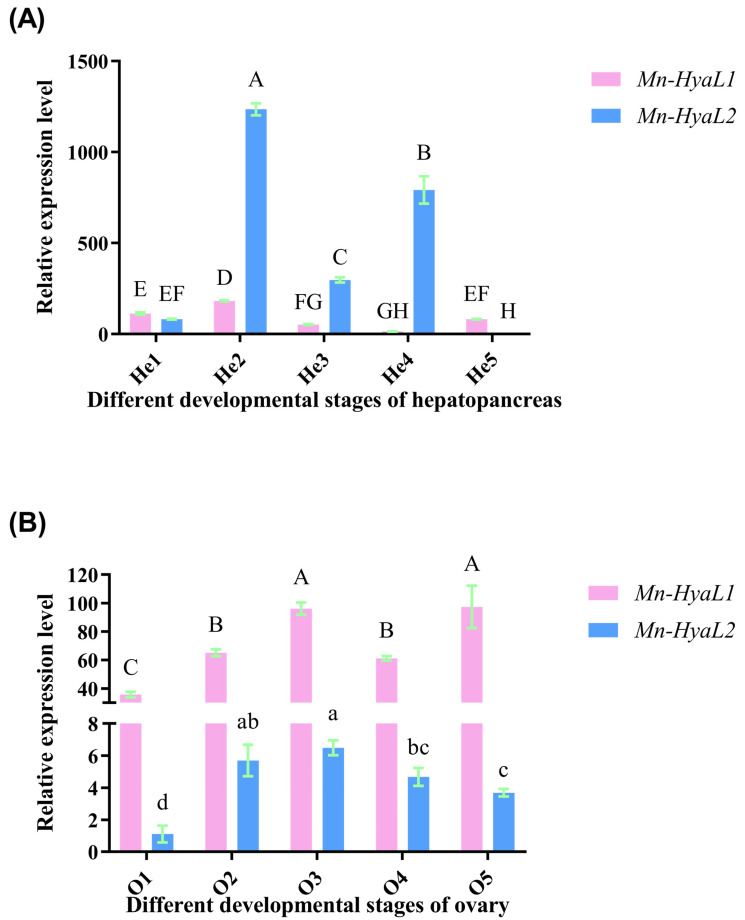 Figure 5
Figure 5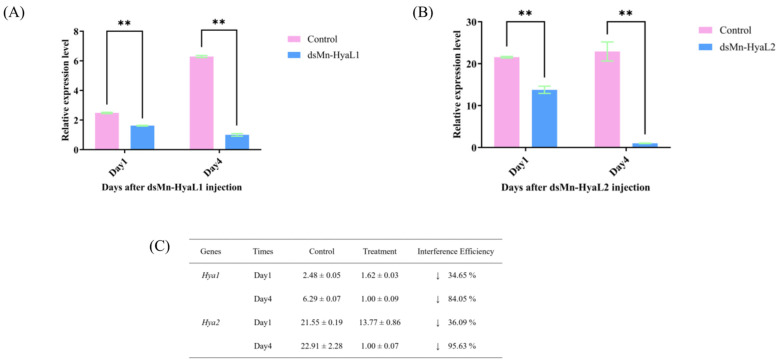 Figure 6
Figure 6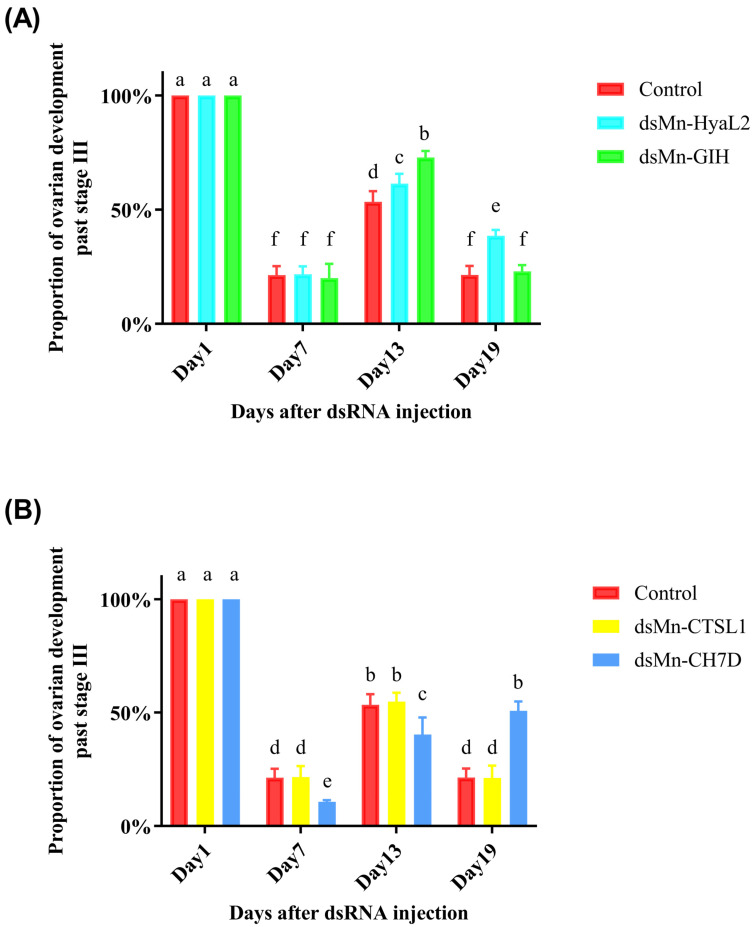 Figure 7
Figure 7- —Central Public-interest Scientific Institution Basal Research Fund CAFS
- —the earmarked fund
- —Seed Industry Revitalization Project of Jiangsu Province
- —Jiangsu Province Platform for the Conservation and Utilization of Agricultural Germplasm
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCephalopods and Marine Biology · Crustacean biology and ecology · Reproductive biology and impacts on aquatic species
1. Introduction
Macrobrachium nipponense is one of the most economically important freshwater aquaculture species in China, with substantial production and significant economic value across East and Southeast Asia. The rapid development of female M. nipponense during the breeding season results in the co-occurrence of individuals at vastly different developmental stages under aquaculture conditions. This heterogeneity within populations leads to significant production challenges, including reduced growth rates, low survival rates, and decreased disease resistance due to high-density crowding [1,2]. Therefore, investigating the underlying molecular mechanisms of precocious sexual maturation in M. nipponense is of great significance for resolving these production challenges.
The rapid ovarian development process is characterized by an intensive period of protein synthesis [3,4]. Consistent with this, our study observed a marked increase in protein content in the ovary concomitant with a decrease in the hepatopancreas throughout ovarian maturation. This pattern of nutrient redistribution has been well-documented in various crustacean species, including Fenneropenaeus merguiensis [5] and Portunus trituberculatus [6], supporting the hypothesis that proteins or their precursors are transported from the hepatopancreas to the developing ovary via the hemolymph. Therefore, in our preliminary experiments, we conducted a comparative transcriptomic analysis of hepatopancreas samples from female M. nipponense across ovarian stages I to V. The comparison between stage I and stage II revealed that the expression levels of two homologous Hyaluronidase-like genes (Mn-HyaL1 and Mn-HyaL2) were significantly upregulated from ovarian stage I to stage II [7].The gene is annotated to the glycosaminoglycan degradation pathway, a component of the lysosomal pathway. Existing evidence has established that this pathway is critically involved in regulating ovarian maturation in M. nipponense [8]. Previous studies have established that several genes within this pathway play important roles in regulating ovarian development in Macrobrachium nipponense [1,9,10,11]. These results led us to hypothesize that Mn-HyaL1 and Mn-HyaL2 could be functionally implicated in the ovarian development of M. nipponense, which became the focus of the present investigation.
Based on functional studies published by previous researchers, several genes have been confirmed to regulate ovarian development in M. nipponense (Table 1). A major limitation, however, lies in the heterogeneity of experimental settings—such as season, tank size, and measured parameters—which complicates cross-study comparisons. To address this gap, a controlled comparative analysis is needed to rank these genes by efficacy, information vital for aquaculture productivity. Based on an analysis of published functional studies, key regulatory genes of ovarian development in Macrobrachium nipponense were selected for comparative RNA interference (RNAi) assays. Among these, Cholesterol 7-desaturase (Mn-CH7D) [2] and Mn-Cathepsin L1 (Mn-CTSL1) [10] were identified as strong promoters of ovarian development, consistently inducing an advance of more than two developmental stages in RNAi-silencing experiments, while Mn-Gonad-inhibiting hormone (Mn-GIH) [12] acted as a potent suppressor. These well-characterized regulators were therefore prioritized for further functional comparison under identical long-term RNAi conditions.
In this study, bioinformatic analyses were conducted to characterize the sequences and phylogenetic relationships of Mn-HyaL1 and Mn-HyaL2. The expression patterns of these two genes were examined by qPCR across various developmental stages, sexes, seasons, and tissues, with a particular focus on their expression during ovarian and hepatopancreas development. Furthermore, RNA interference (RNAi) was employed to knock down Mn-HyaL1, Mn-HyaL2, and other previously validated key regulators of ovarian development in M. nipponense in parallel experimental groups. This approach enabled us not only to investigate the regulatory functions of these genes in ovarian maturation but also to compare their relative functional strengths. The findings of this study may provide valuable insights for addressing the issue of precocious ovarian maturation in female M. nipponense.
2. Results
2.1. Characterization of the Full-Length Sequences of Mn-HyaL1 and Mn-HyaL2
The open reading frame (ORF) of the hyaluronidase-like1 gene in Macrobrachium nipponense was 1101 bp in length, encoding a protein of 366 amino acids, and it was designated as Mn-HyaL1 (GenBank accession no. PV941746). The ORF of the hyaluronidase-like2 gene in M. nipponense was 1164 bp, encoding a protein of 387 amino acids, and was designated as Mn-HyaL2 (GenBank accession no. PV941747). The cDNA and deduced amino acid sequences of both Mn-HyaL1 and Mn-HyaL2 are presented in Figure 1.
Bioinformatic analysis revealed that the molecular weight (Mw) of the Mn-HyaL1 protein is 41,906.46 Da, with a theoretical isoelectric point (pI) of 5.41. Analysis of the full-length amino acid composition indicated that glycine (G) was the most abundant residue, constituting 7.4% of the sequence. The protein contains 36 positively charged residues (Arg + Lys) and 49 negatively charged residues (Asp + Glu). The predicted molecular formula is C_1877_H_2841_N_497_O_554_S_21_. Secondary structure prediction showed that Mn-HyaL1 comprises 9 α-helices, 7 η-helices, 9 β-sheets, and 3 β-turns; the sequential arrangement of these elements is provided in Figure S1. Furthermore, Mn-HyaL1 was predicted to lack both a signal peptide and transmembrane domains.
Bioinformatic analysis indicated that the Mn-HyaL2 protein has a molecular weight (Mw) of 44,331.21 Da and a theoretical isoelectric point (pI) of 4.99. Analysis of its full-length amino acid composition showed that leucine (L) was the most abundant residue at 9.3%. The protein contains 37 positively charged residues (Arg + Lys) and 56 negatively charged residues (Asp + Glu), with a predicted molecular formula of C_1986_H_3031_N_515_O_598_S_20_. The predicted secondary structure consists of 9 α-helices, 5 η-helices, 10 β-sheets, and 5 β-turns; the sequential arrangement is detailed in Figure S1. Mn-HyaL2 was predicted to contain one transmembrane domain located between residues 28 and 45. Additionally, a signal peptide (Sec/SPI type) was identified at the N-terminus (residues 1–47), with a cleavage site between residues 47 and 48. The signal peptide sequence was determined to be MMWSALHQTNKREKLPQTSAKMGRMKELAFALTLLAVCVLTGYTTEA. The region from residue 48 to 387 was predicted to constitute the non-cytoplasmic domain.
Sequence analysis revealed that both Mn-HyaL1 and Mn-HyaL2 contain the same conserved domain, Hyaluronidase (Accession: pfam01630), located at residues 30–336 in Mn-HyaL1 and residues 48–351 in Mn-HyaL2. Furthermore, four identical functional domains were identified in both proteins: Hyaluronidase (IPR018155), Aldolase-type TIM barrel (IPR013785), Glycoside hydrolase superfamily (IPR017853), and Glycoside hydrolase family 56, bee venom hyaluronidase (IPR001329).
2.2. Analysis of Species Similarity and Phylogeny of Mn-HyaL1 and Mn-HyaL2
Sequence alignment of Mn-HyaL1 and Mn-HyaL2 was performed using DNAMAN 9.0, revealing a similarity of 60.21% between the two isoforms. Multiple sequence alignment was then conducted using Mn-HyaL1 and Mn-HyaL2 alongside homologous genes from other species, which included, in order: Macrobrachium rosenbergii, Palaemon carinicauda, Halocaridina rubra, Cherax quadricarinatus, Penaeus indicus, Penaeus chinensis, and Penaeus vannamei. Nucleotide sequence similarities were calculated using the p-distance method. The results showed that Mn-HyaL1 shared sequence similarities of 93.56%, 63.77%, 50.99%, 46.93%, 46.58%, 45.35%, and 44.86% with the corresponding genes from these species, respectively. Similarly, Mn-HyaL2 exhibited similarities of 61.78%, 79.01%, 54.98%, 46.79%, 48.84%, 46.18%, and 47.58%, respectively (Figure S2).
A phylogenetic tree was constructed using MEGA 11.0 based on the amino acid sequences of Mn-HyaL1, Mn-HyaL2, and their homologs from other species. The analysis revealed that Mn-HyaL1 and Mn-HyaL2 did not cluster directly with each other. Instead, each first clustered with distinct homologs from Macrobrachium rosenbergii and Palaemon carinicauda (both belonging to the family Palaemonidae), before subsequently grouping together. This combined cluster then grouped with other crustaceans, such as Halocaridina rubra, and finally with insects (Figure 2).
2.3. Analysis of Spatiotemporal Expression Patterns of Mn-HyaL1 and Mn-HyaL2
2.3.1. Analysis of Stage-Specific Expression Patterns of Mn-HyaL1 and Mn-HyaL2
The expression patterns of Mn-HyaL1 and Mn-HyaL2 during embryonic and post-embryonic developmental stages are shown in Figure 3. During early embryonic development (from CS to ZS), Mn-HyaL1 expression remained consistently high at both the Cleavage stage (CS) and Zoea Stage (ZS) with no significant difference between these two stages, whereas Mn-HyaL2 expression peaked specifically at ZS. During the post-embryonic developmental stage (from L1 to L15), Mn-HyaL1 expression was highest at the L10 stage, while Mn-HyaL2 expression peaked at the L15 stage. In the subsequent stage (from PL1 to PL25), Mn-HyaL1 expression reached its maximum at the PL15 stage, whereas Mn-HyaL2 showed the highest expression at the PL25 stage. Overall, the highest expression level of Mn-HyaL1 was observed at the L10 stage, and that of Mn-HyaL2 was observed at the PL25 stage.
2.3.2. Analysis of Tissue-Specific Expression Patterns of Mn-HyaL1 and Mn-HyaL2
As shown in Figure 4, the highest expression levels of both Mn-HyaL1 and Mn-HyaL2 were detected in the hepatopancreas, followed by the gonads. Furthermore, in most tissues examined, the expression of these genes was generally higher during the reproductive season than during the non-reproductive season, with this pattern being observed in both female and male shrimp.
2.3.3. Analysis of Stage-Specific Expression Patterns of Mn-HyaL1 and Mn-HyaL2 in Hepatopancreatic and Ovarian Tissues
In the hepatopancreas (Figure 5A), the expression levels of both Mn-HyaL1 and Mn-HyaL2 increased significantly from stage He1 to He2 (p < 0.01), peaking at stage He2. The expression at He2 was significantly higher than that at all other stages (p < 0.01). The lowest expression level of Mn-HyaL1 was observed at stage He4, whereas Mn-HyaL2 exhibited its minimum expression at stage He5.
In the ovaries (Figure 5B), the expression levels of both Mn-HyaL1 and Mn-HyaL2 increased significantly from stage O1 to O2 (p < 0.01). The highest expression of Mn-HyaL1 was observed at stage O5, while Mn-HyaL2 peaked at stage O3. The lowest expression for both genes occurred at stage O1, which was significantly lower than that at all other stages (p < 0.01). Additionally, the expression level of Mn-HyaL1 was significantly higher than that of Mn-HyaL2 at each ovarian stage (p < 0.05).
2.4. Functional Characterization and Comparative Analysis of Genes
2.4.1. Evaluation of RNA Interference Efficiency
To further investigate the functional roles of Mn-HyaL1 and Mn-HyaL2 in ovarian maturation, RNA interference (RNAi) was employed. As shown in Figure 6, the knockdown efficiencies of dsMn-HyaL1 were 34.65% and 84.05% on day 1 and day 4 post-injection, respectively (p < 0.01). Similarly, dsMn-HyaL2 achieved knockdown efficiencies of 36.09% and 95.63% on day 1 and day 4, respectively (p < 0.01). Both dsRNAs induced significant gene silencing, with dsMn-HyaL2 exhibiting a higher knockdown efficiency. For Mn-CH7D [2], Mn-CTSL1 [10], and Mn-GIH [12], we employed the identical dsRNA sequences and experimental conditions that were previously validated in their original functional studies. These studies reported knockdown efficiencies of 89.92% for Mn-CH7D, 99.79% for Mn-CTSL1, and 83% for Mn-GIH. By maintaining complete consistency with these established protocols, we ensured comparable silencing efficacy across all target genes in the present comparative analysis.
2.4.2. Comparative Functional Analysis of Key Genes in Ovarian Development of M. nipponense
A comparison of ovarian development between the experimental and control groups is presented in Figure 7. At the start of the experiment, all shrimp were at stage O3. By day 6, all groups had entered a new reproductive cycle. From day 6 to day 12, the proportion of shrimp with ovaries beyond stage III gradually increased in all groups. By day 18, all groups except the Mn-CH7D group had entered a third reproductive cycle. Overall, the order of ovarian development speed from slowest to fastest following RNAi was: Mn-CH7D experimental group < Mn-CTSL1 experimental group = control group < Mn-HyaL2 experimental group < Mn-GIH experimental group. Due to a high mortality rate, the Mn-HyaL1 experimental group was excluded from the statistical analysis as the resulting data were insufficient for meaningful interpretation.
3. Discussion
While hyaluronidases are well-characterized in vertebrate and microbial systems for their roles in extracellular matrix remodeling [28,29], their functions in crustacean reproduction remain largely unexplored [30]. The present study provides the first functional characterization of two hyaluronidase-like genes, Mn-HyaL1 and Mn-HyaL2, in the ovarian development of Macrobrachium nipponense.
The HyaL enzymes characterized in this study are categorized as endo-hyaluronidases (hyaluronoglucosaminidase, EC 3.2.1.35) [31], a class of glycoside hydrolases predominantly sourced from vertebrates and venoms, which degrade hyaluronic acid via hydrolysis of β-1,4-glycosidic bonds to produce specific oligosaccharides and can also target chondroitin sulfate or dermatan sulfate [32], thus differing fundamentally from other hyaluronidase types (EC 3.2.1.36; EC 4.2.2.1) [33,34,35].
Evidence from reproductive studies suggests conserved roles for hyaluronidases in fertilization processes, as exemplified by the functional similarity between bee venom hyaluronidase and the mammalian sperm membrane protein PH-20, both facilitating gamete interaction through hyaluronic acid degradation [36,37,38]. Hyaluronidase activity is significantly correlated with fertilization rates [38,39,40]. In ovarian contexts, hyaluronidase activity has been linked to follicular atresia and tissue remodeling [28,41]. Furthermore, hyaluronidase dysregulation has been associated with several ovarian disorders [42,43,44]. An age-related decline in hyaluronic acid content is observed in the ovarian stroma [45]. But its specific role in crustacean ovarian development remains largely uncharacterized. Limited studies have revealed that hyaluronidase activity in hepatopancreas homogenates of the red king crab (Paralithodes camtschaticus) is an order of magnitude higher than that of commercial preparations, suggesting its potential as a high-quality source for hyaluronidase applications in cosmetics and medicine [46,47]. The present study addresses this gap by functionally characterizing two hyaluronidase-like genes in Macrobrachium nipponense, providing the first insights into their regulatory contributions to ovarian maturation in crustaceans.
Bioinformatic analysis revealed that both genes belong to the Glycoside Hydrolase Family 56 (GH56), a group typically associated with arthropod venoms where they act as “spreading factors” by degrading extracellular matrix components [48,49,50,51]. Notably, sequence analysis indicated structural divergence between the two isoforms: Mn-HyaL2 possesses an N-terminal signal peptide and transmembrane domain absent in Mn-HyaL1, suggesting distinct subcellular localization and potential functional specialization.
The screening results of the Mn-HyaL1 and Mn-HyaL2 genes were obtained from a preliminary experiment. In the present study, Mn-HyaL1 and Mn-HyaL2 were identified from a comparative transcriptomic analysis of the hepatopancreas of M. nipponense at ovarian stages O1 and O2, where their expression increased significantly from stage O1 to O2 [7]. Subsequent KEGG enrichment analysis revealed that both genes are involved in the “Lysosome” signaling pathway (KEGG: map04142), which has been previously demonstrated to be closely associated with ovarian development in M. nipponense [8]. The relationship between this pathway and ovarian development, as well as other regulatory genes within the pathway, has been discussed in earlier studies [9].
The observed involvement of Mn-HyaL1 and Mn-HyaL2 in ovarian development can be understood within the functional framework of lysosomal activity. In crustaceans, lysosomes contribute to steroidogenesis through two well-established mechanisms: facilitating steroid precursor availability and modulating regulators of steroid production, thereby maintaining endocrine homeostasis [52]. Specifically, lysosomes enable cholesterol release from endocytosed LDL via coordinated action of enzymes and transporters such as NPC1 and NPC2 [53], while also participating in the degradation of key regulatory complexes including LH-LHR, FSH-FSHR, and PGF2α receptors [54,55,56].
As hyaluronidases, the primary function of Mn-HyaL1 and Mn-HyaL2 lies in hydrolyzing hyaluronic acid—a key component of the extracellular matrix. This functional similarity to lysosomal hydrolases suggests their potential involvement in analogous processes during ovarian maturation. Notably, hyaluronidase activity has been shown to play a key role in the early phases of folliculogenesis by negatively regulating ovarian follicle growth and survival [57]. This regulatory function may operate through modulation of the extracellular matrix environment surrounding developing follicles, thereby influencing follicular development and oocyte maturation.
We hypothesize that Mn-HyaL1 and Mn-HyaL2 may facilitate follicular remodeling, nutrient mobilization, and hormonal regulation through the controlled degradation of hyaluronan-rich matrices. Such activity could potentially influence the availability of signaling molecules or growth factors embedded in the extracellular matrix, thereby indirectly supporting the lysosome-mediated pathways essential for ovarian development. The distinct expression patterns and functional characteristics of these two isoforms suggest they may fulfill complementary roles at different stages of ovarian maturation. Further studies are needed to elucidate the specific mechanisms by which these hyaluronidase isoforms contribute to reproductive maturation in M. nipponense, particularly their potential interplay with established lysosomal pathways.
Bioinformatic analysis revealed that Mn-HyaL1 and Mn-HyaL2 share highly similar conserved and functional domains, suggesting overall functional similarity. However, their nucleotide and amino acid sequence alignment showed limited homology, with an amino acid identity of only 60.21%. Furthermore, Mn-HyaL2 possesses an N-terminal signal peptide and a transmembrane domain, which are absent in Mn-HyaL1. Phylogenetic analysis also indicated that Mn-HyaL1 and Mn-HyaL2 do not form a direct cluster. These observations suggest that while the two isoforms may share core functions due to their conserved domain architecture, structural differences could lead to functional distinctions in specific aspects such as subcellular localization or regulatory mechanisms.
The distinct domain architectures of Mn-HyaL1 and Mn-HyaL2 provide important insights into their potential functional differentiation. While both proteins share identical catalytic domains (Hyaluronidase pfam01630) and functional motifs, the exclusive presence of an N-terminal signal peptide and transmembrane domain in Mn-HyaL2 suggests divergent subcellular targeting mechanisms. This structural distinction supports the hypothesis that Mn-HyaL2 may function as a secreted or membrane-associated protein, potentially modifying extracellular matrix components or participating in cell surface signaling events. In contrast, Mn-HyaL1, lacking these targeting signals, likely operates intracellularly, possibly regulating internal pools of hyaluronic acid or other glycosaminoglycans.
This spatial segregation could explain the observed functional specialization between the two isoforms, as they would access distinct substrates and interact with different regulatory partners within separate cellular compartments. The combination of conserved catalytic domains with divergent targeting sequences suggests an evolutionary strategy where core enzymatic function is maintained while cellular localization and substrate specificity are diversified. This pattern aligns with the phylogenetic evidence indicating an ancient gene duplication event followed by functional divergence, ultimately yielding complementary but non-redundant roles in ovarian development regulation.
In the tissue-specific expression analysis, although the expression levels of Mn-HyaL1 and Mn-HyaL2 varied across different stages, both isoforms were generally highly expressed during the mid-to-late phases of each stage. These late phases often represent critical periods in the development of M. nipponense that demand substantial energy expenditure. Considering the fundamental hydrolytic function of hyaluronidase-like genes, we hypothesize that the two Mn-HyaL isoforms are involved in energy regulation during these key stages. This notion is further supported by the concurrent upregulation of both isoforms during the reproductive season, a period of heightened energy demands for activities such as growth, mating, and spawning. Additionally, the expression of both Mn-HyaL1 and Mn-HyaL2 was significantly higher in the hepatopancreas than in other tissues. As the hepatopancreas is the primary organ supplying energy to the ovaries, and given that both isoforms showed increased expression from stage O1/He1 to O2/He2—a period characterized by rapid vitellogenesis and the transfer of energy substrates like glycoproteins from the hepatopancreas to the ovaries [7,8]—these results collectively reinforce the hypothesis that Mn-HyaL isoforms may participate in the energy regulation supporting ovarian development.
In the RNA interference (RNAi) experiment, after 18 days, all experimental groups except the Mn-CH7D group had entered the third reproductive cycle, indicating that Mn-CH7D has the most significant promotive effect on ovarian development. Conversely, the ovarian development speed in the Mn-HyaL2 and Mn-GIH groups was faster than that in the control group, identifying them as inhibitory genes, with Mn-GIH exhibiting a stronger effect than Mn-HyaL2 (Figure 7A). No significant difference was observed between the Mn-CTSL1 group and the control group, a finding consistent with previous reports on Mn-CTSL2 [11]. The Mn-HyaL1 experimental group exhibited an unexpectedly high mortality rate, which precluded meaningful data analysis. This suggests that Mn-HyaL1 may play a critical role in essential physiological processes such as immunity, molting, or other vital growth pathways. This intriguing phenomenon will be the focus of our future investigations. Future studies could explore alternative interference methods (e.g., oral delivery of dsRNA) to mitigate the high mortality observed in this study [58].
The relationship between Mn-HyaL1 and Mn-HyaL2 genes is a key point to be discussed. The evolutionary relationship between Mn-HyaL1 and Mn-HyaL2 provides important context for interpreting their functional roles. Phylogenetic analysis revealed that Mn-HyaL1 and Mn-HyaL2 did not form a distinct clade, with each instead clustering more closely with its putative ortholog from Macrobrachium rosenbergii (Figure 2). This topology suggests that the divergence of these two genes is an ancient event, likely predating the speciation of M. nipponense and M. rosenbergii, and that they are therefore best classified as orthologs rather than recently duplicated paralogs. However, despite this orthologous origin, several lines of evidence indicate that Mn-HyaL1 and Mn-HyaL2 have undergone significant functional divergence within M. nipponense. Their relatively low amino acid sequence similarity (60.21%), distinct spatiotemporal expression profiles across all examined tissues and developmental stages, and the markedly different phenotypic consequences of their RNAi knockdown (e.g., the high mortality observed following Mn-HyaL1 silencing) collectively support this notion of functional specialization. Thus, while Mn-HyaL1 and Mn-HyaL2 share a common ancestral gene, evolutionary pressures appear to have shaped them into regulators with non-redundant, and potentially complementary, functions in ovarian development.
In summary, this study successfully identified the following key findings and conclusions: (1) The full-length cDNA sequences of Mn-HyaL1 and Mn-HyaL2 were cloned and characterized for the first time. (2) The spatiotemporal expression patterns of Mn-HyaL1 and Mn-HyaL2 were systematically investigated across different developmental stages, sexes, seasons, and tissues, with a particular focus on their expression during ovarian and hepatopancreas development. (3) Functional validation via RNAi confirmed that Mn-HyaL acts as a negative regulator of ovarian development in M. nipponense. This finding is significant as most previously identified regulators in this species are positive regulators. Notably, this study provides the first evidence of a regulatory role for a Hyaluronidase-like gene in ovarian development, and represents the first functional characterization of a HyaL gene in crustaceans. (4) The relative potency of the studied genes in promoting ovarian development was determined to be Mn-CH7D > Mn-CTSL1. The inhibitory potency on ovarian development followed the order: Mn-GIH > Mn-HyaL2. This study is the first to conduct a direct comparative analysis of these key regulators under identical conditions, thereby identifying the most potent positive and negative regulators. These results contribute to elucidating the molecular mechanisms underlying ovarian development in M. nipponense and have important implications for the genetic breeding of improved germplasm resources in aquaculture.
4. Materials and Methods
4.1. Experimental Animals and Breeding Conditions
Healthy female specimens of Macrobrachium nipponense used in this study were obtained from the Freshwater Fisheries Research Center of the Chinese Academy of Fishery Sciences in Wuxi City, Jiangsu Province, China. All selected shrimp were at ovarian stage III and had a uniform body weight (1 ± 0.18 g). The shrimp were reared in indoor recirculating aquaculture systems using fiberglass tanks (95 cm × 45 cm × 45 cm). The tanks were equipped with plastic aquatic plants to provide shelter. The shrimp were fed twice daily (morning and evening) with a commercial diet at a rate of approximately 5% of their total body weight per feeding. The water temperature was maintained at an average of 29 °C.
4.2. Tissue Sample Collection
Various tissues, including the eye, cerebral ganglion, heart, hepatopancreas, gill, muscle, and gonad, were rapidly dissected from M. nipponense and immediately frozen in liquid nitrogen. All collected samples were subsequently stored at −80 °C until further analysis. Additionally, individual shrimp samples at different developmental stages, ranging from embryonic to larval phases, were also collected and similarly stored at −80 °C for subsequent experiments. The specific criteria for classifying ovarian stages and embryonic developmental stages are provided in Table S1 [9,59].
M. nipponense specimens were euthanized by rapid freezing using liquid nitrogen. Live prawns were transferred to a sealed container filled with an adequate volume of liquid nitrogen, where the ultra-low temperature (−196 °C) induced instantaneous loss of consciousness and rapid death. This method minimizes potential suffering and meets ethical requirements for crustacean euthanasia. Death was confirmed by the absence of movement and rigidification of the body.
4.3. Molecular Biology Methods
Total RNA was extracted from M. nipponense tissues at various developmental stages using RNAiso Easy reagent (Takara, Dalian, China). Tissue samples were homogenized by a combination of manual grinding (for larger tissues) and mechanical disruption using a tissue homogenizer to ensure complete cell lysis. For early developmental stages where tissue mass was limited, multiple individuals were pooled to obtain sufficient RNA yield per sample. RNA integrity was assessed by 1.2% agarose gel electrophoresis. First-strand cDNA was synthesized from the RNA templates using the M-MLV Reverse Transcriptase kit (Takara, Kusatsu, Japan) and stored at −20 °C for subsequent quantitative real-time PCR (qPCR) analysis to examine the expression patterns of Hyaluronidase-like genes. The elongation factor gene (EIF), which has been validated in previous studies [60], was used as the internal reference. Gene expression levels were calculated using the 2^−ΔΔCT^ method [61].
4.4. Bioinformatics Analysis
The cDNA fragments of the target genes, Mn-HyaL1 and Mn-HyaL2, were obtained from a cDNA library derived from the hepatopancreas of M. nipponense at different ovarian stages, which was constructed in our laboratory during previous research [7]. The tools and websites used for analyzing the nucleotide and amino acid sequences of the target genes are listed in Table S2. All primers used in this study are provided in Table S3.
4.5. RNA Interference Experiment
Double-stranded RNA (dsRNA) was synthesized using the Transcript Aid™ T7 High Yield Transcription Kit (Fermentas, Inc., Waltham, MA, USA), with primers listed in Table S3. The concentration of the synthesized dsRNA was measured at 260 nm using a biophotometer (Eppendorf, Hamburg, Germany).
For the short-term interference assay, hepatopancreas samples were collected on days 1 and 4 after dsRNA injection for cDNA synthesis. The knockdown efficiency was determined by quantifying the relative expression levels of the target genes in the dsMn-HyaL1 and dsMn-HyaL2 groups via qPCR. The dsRNA yielding the higher interference efficiency was selected for the long-term experiment.
In the long-term trial, a total of 720 healthy female M. nipponense at ovarian stage III were randomly and equally distributed into 18 aquaculture tanks. These represented five experimental groups: Mn-HyaL1, Mn-HyaL2, Mn-CH7D, Mn-CTSL1, Mn-GIH, and a control group, each with three replicates. Shrimp in the treatment groups received an intracardiac injection of the corresponding dsRNA at a dose of 4 μg per gram of body weight [2], while the control group was injected with an equivalent amount of dsGFP. Booster injections were administered every 6 days. The ovarian development stage of each shrimp were recorded daily. The percentage of shrimp exhibiting ovarian development beyond stage III was calculated.
4.6. Statistical Analysis
All quantitative data are presented as the mean ± standard deviation (SD). Data analysis was performed using SPSS Statistics 24.0. One-way analysis of variance (ANOVA) followed by Duncan’s test was used for multiple comparisons to assess significant differences between the control and treatment groups. A significance level of p < 0.05 was applied. Relative gene expression was calculated using the 2^−ΔΔCT^ method.
5. Conclusions
In conclusion, this study provides the first comprehensive functional characterization of Mn-HyaL1 and Mn-HyaL2 in a crustacean species through an integrated approach combining bioinformatics, molecular, and physiological analyses. Notably, we demonstrate for the first time in crustaceans that Mn-HyaL functions as a negative regulator of ovarian development in Macrobrachium nipponense, which also represents the initial discovery of a regulatory role for a Hyaluronidase-like gene in the ovarian development process. Furthermore, this work is the first to conduct a direct comparative analysis of key regulatory genes under identical conditions in M. nipponense, identifying the most potent positive and negative regulators. These findings significantly contribute to elucidating the molecular mechanisms governing ovarian development in this commercially important species and hold substantial promise for the genetic improvement of germplasm resources in aquaculture.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cheng D. Zhang W. Jiang S. Xiong Y. Jin S. Pan F. Zhu J. Gong Y. Wu Y. Qiao H. Cathepsin D Plays a Vital Role in Macrobrachium nipponense of Ovary Maturation: Identification, Characterization, and Function Analysis Genes 202213149510.3390/genes 1308149536011406 PMC 9408384 · doi ↗ · pubmed ↗
- 2Wang J. Jiang S. Zhang W. Xiong Y. Jin S. Cheng D. Zheng Y. Qiao H. Fu H. Function Analysis of Cholesterol 7-Desaturase in Ovarian Maturation and Molting in Macrobrachium nipponense: Providing Evidence for Reproductive Molting Progress Int. J. Mol. Sci.202324694010.3390/ijms 2408694037108104 PMC 10138363 · doi ↗ · pubmed ↗
- 3Li X. Han T. Zheng S. Wu G. Nutrition and Functions of Amino Acids in Aquatic Crustaceans Amino Acids in Nutrition and Health: Amino Acids in the Nutrition of Companion, Zoo and Farm Animals Wu G. Springer International Publishing Cham, Switzerland 2021169198978-3-030-54462-1
- 4Izquierdo M.S. Fernández-Palacios H. Tacon A.G.J. Effect of Broodstock Nutrition on Reproductive Performance of Fish Aquaculture 2001197254210.1016/S 0044-8486(01)00581-6 · doi ↗
- 5Fatima H. Ayub Z. Ali S. Siddiqui G. Biochemical Composition of the Hemolymph, Hepatopancreas, Ovary, and Muscle during Ovarian Maturation in the Penaeid Shrimps Fenneropenaeus merguiensis and F. penicillatus (Crustacea: Decapoda)Turk. J. Zool.20133733434710.3906/zoo-1201-12 · doi ↗
- 6Feng W. Zhao Z. Wang J. Han T. Nutrient Composition of Ovary, Hepatopancreas and Muscle Tissues in Relation to Ovarian Development Stage of Female Swimming Crab, Portunus trituberculatus Animals 202313322010.3390/ani 1320322037893944 PMC 10603639 · doi ↗ · pubmed ↗
- 7Jiang S. Zhang W. Xiong Y. Cheng D. Wang J. Jin S. Gong Y. Wu Y. Qiao H. Fu H. Hepatopancreas Transcriptome Analyses Provide New Insights into the Molecular Regulatory Mechanism of Fast Ovary Maturation in Macrobrachium nipponense BMC Genom.20222362510.1186/s 12864-022-08851-836045344 PMC 9429573 · doi ↗ · pubmed ↗
- 8Jiang S. Qiao H. Fu H. Gu Z. Hepatopancreas Proteomic Analysis Reveals Key Proteins and Pathways in Regulatory of Ovary Maturation of Macrobrachium nipponense Animals 20231397710.3390/ani 1306097736978518 PMC 10044353 · doi ↗ · pubmed ↗