Antibiotics Impact the Cytotoxicity and Cytopathic Effect of Helicobacter pylori Extracellular Vesicles Against Gastric Cells
Paweł Krzyżek, Mateusz Chmielarz, Edyta Bożemska, Agnieszka Opalińska, Mateusz Olbromski, Michał Małaszczuk, Barbara Krzyżanowska, Katarzyna Haczkiewicz-Leśniak, Marzenna Podhorska-Okołów, Piotr Dzięgiel, Beata Sobieszczańska

TL;DR
This study shows that antibiotics can change how Helicobacter pylori's extracellular vesicles affect stomach cells, but not always in a harmful way.
Contribution
The study is the first to show that subinhibitory antibiotics modify the virulence of H. pylori extracellular vesicles.
Findings
Subinhibitory antibiotics alter the cytotoxicity and cytopathic effect of H. pylori extracellular vesicles.
EVs from antibiotic-exposed H. pylori showed varied effects on vacuolization and the hummingbird phenotype.
EVs from non-exposed H. pylori had the highest lethal effect on gastric cells.
Abstract
Helicobacter pylori is a spiral microorganism capable of inducing a range of gastric diseases. Among different virulence determinants produced by this bacterium, VacA and CagA are of critical importance for the development of these conditions. Taking into account the ability to chronically colonize the stomach, drug-resistant strains of this pathogen can be repeatedly exposed to subinhibitory antibiotic concentrations, which in turn may reduce or enhance their extracellular vesicles (EVs)-derived virulence towards gastric cells. With the use of different experimental techniques, we were the first to demonstrate that subinhibitory antibiotic concentrations modify both the cytotoxicity and cytopathic effect induced by EVs of H. pylori in gastric cells. The ability to induce vacuolization and the hummingbird phenotype in gastric cells presented an antibiotic-specific pattern. At the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1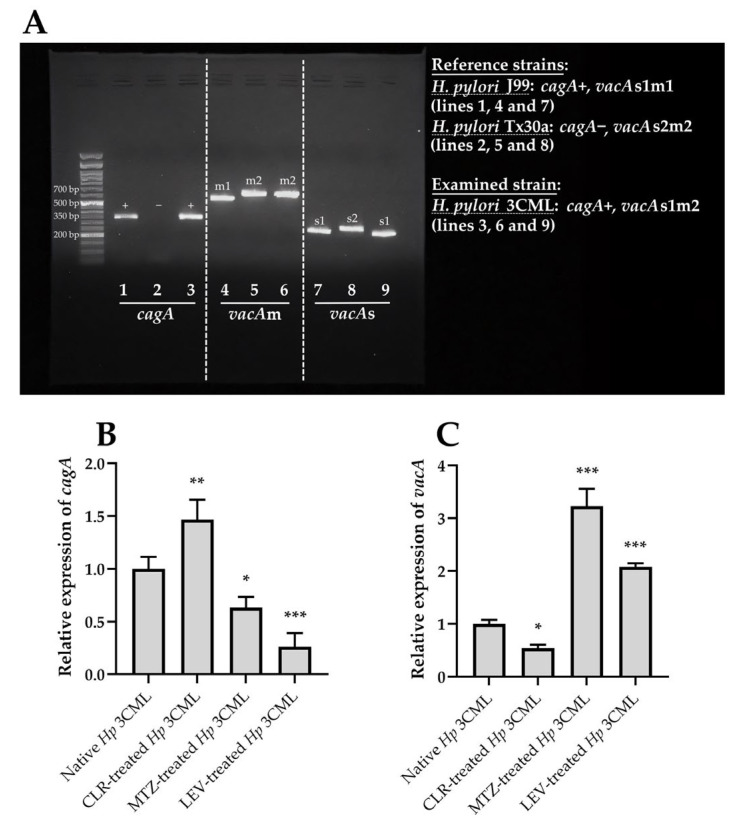 Figure 2
Figure 2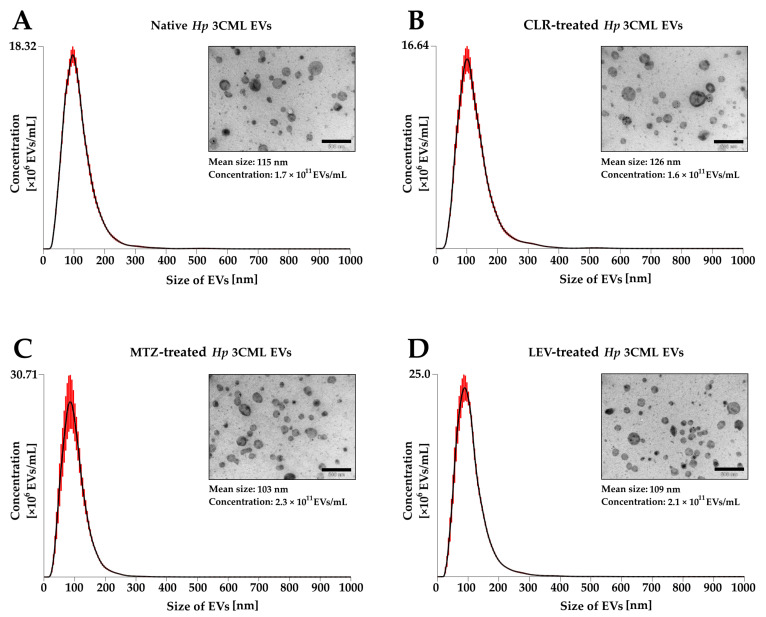 Figure 3
Figure 3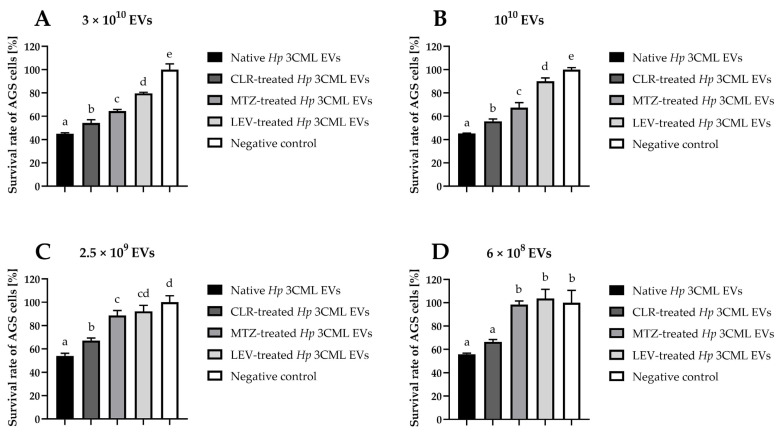 Figure 4
Figure 4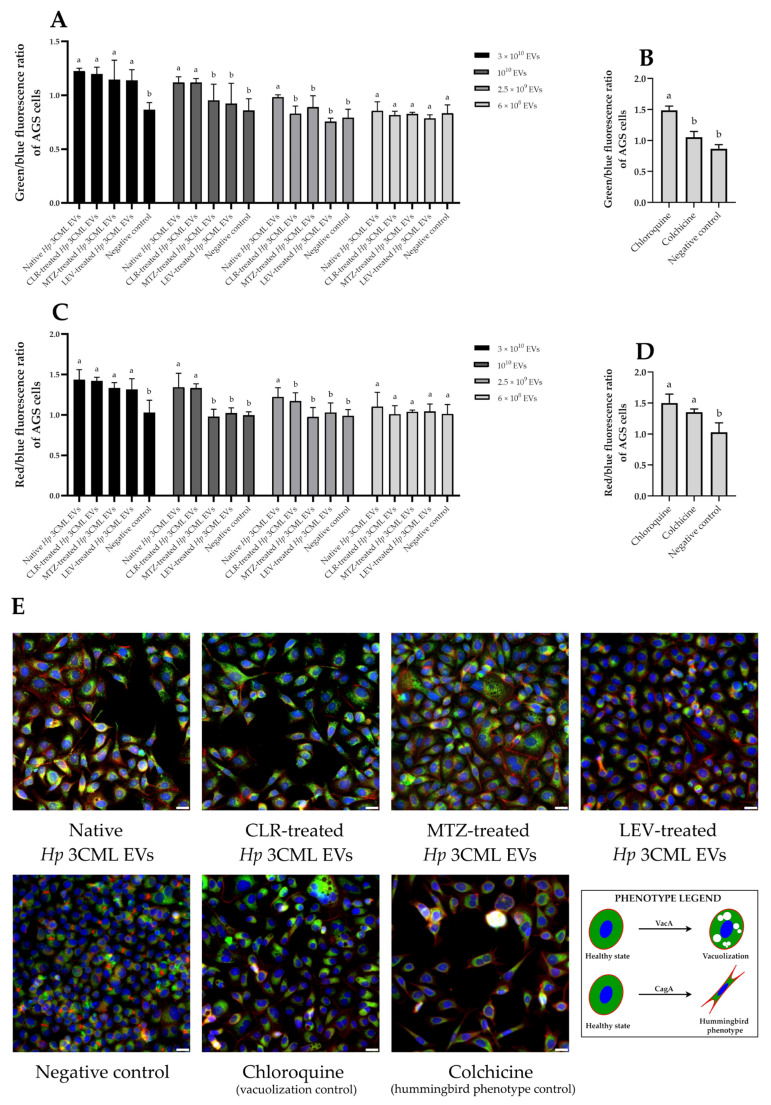 Figure 5
Figure 5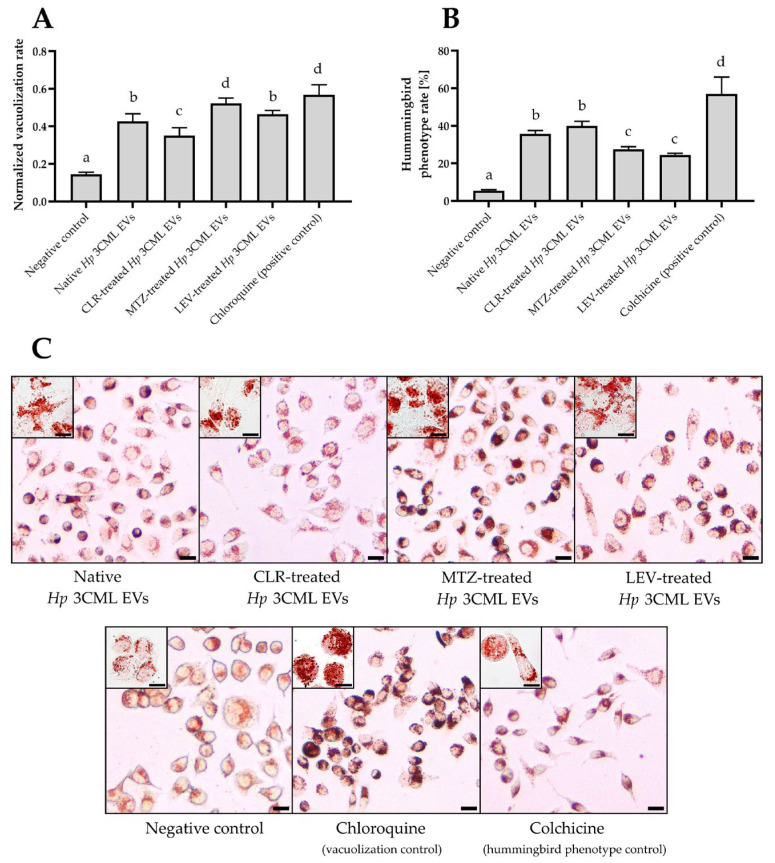 Figure 6
Figure 6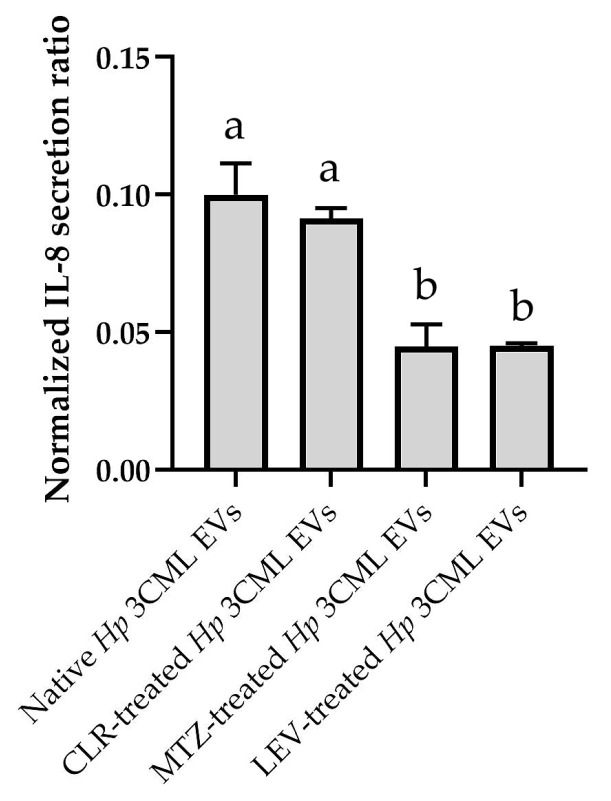 Figure 7
Figure 7- —Wroclaw Medical University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsExtracellular vesicles in disease · Helicobacter pylori-related gastroenterology studies · Bacterial Infections and Vaccines
1. Introduction
Helicobacter pylori is a spiral, microaerophilic, Gram-negative bacterium [1]. This microorganism is capable of colonizing the stomach, thanks to its ability to perceive a chemical gradient and move toward the gastric mucosa [2]. H. pylori infections most often occur in childhood with clinical symptoms remaining undetected by many years [3]. The presence of H. pylori significantly contributes to the physiological and morphological changes in the stomach, which over time can lead to the development of various pathologies of this organ, such as ulceration or carcinogenic processes (adenocarcinomas or mucosa-associated lymphoid tissue lymphomas) [4,5]. Although H. pylori is a non-invasive bacterium, its capacity to induce chronic inflammation and systemically disseminate virulence factors may generate numerous extragastric diseases, ranging from metabolic to cardiovascular and neurological disorders [6]. Among different virulence determinants produced by this bacterium, two toxins are of critical importance for the development of the above-mentioned conditions [4,5,6].
The first is the vacuolating cytotoxin A (VacA), whose gene is present in all H. pylori strains, albeit in different allelic versions [7,8,9]. The s1/m1 and s1/m2 alleles are thought to cause more severe gastric disorders than the s2 allele. VacA is produced as a 140 kDa protoxin that undergoes limited proteolytic cleavage to form an 88 kDa secretory protein composed of two subunits (p33 and p55). VacA is released into the extracellular environment via the type 5 secretion system (T5SS). When it interacts with the host cell, it disrupts signaling processes, depolarizes the membrane, and induces anion-selective efflux of carbonate and organic ions from the cell interior. In this way, VacA facilitates H. pylori survival by increasing the amount of nutrients obtained from the gastric mucosa. Changes in cell membrane permeability and an increase in osmotic pressure contribute to the formation of vacuoles in host cells—a phenotype giving this toxin its name.
The second key virulence factor of H. pylori is the cytotoxin-associated gene A (CagA) [10,11,12]. It is estimated that approximately 70% of strains worldwide produce this toxin, including 60% from Western and 95% from East Asian countries. CagA is a 125–145 kDa oncoprotein injected into host cells via the type 4 secretion system (T4SS). In the cytoplasm of effector cells, CagA undergoes phosphorylation, modulating their signaling and inducing a number of characteristics typical of cancer cells, such as cell death resistance, invasiveness, and genetic instability. Furthermore, CagA disrupts the cytoskeleton of host cells, which consequently contributes to their elongation and increased motility. This phenomenon is referred to as the “hummingbird phenotype” and is the most characteristic pathological change induced by this toxin.
Microorganisms have evolved a variety of single- or two-step secretion systems responsible for the transport of effector proteins into host cells [13,14]. However, an alternative secretion system involving the production of extracellular vesicles (EVs) has been postulated [15]. EVs possess the characteristics of nanoscale vectors, constituting a cocktail of bioactive macromolecules (lipids, proteins or nucleic acids) protected by biological membranes [16]. In this way, microbial components enclosed within the EV lumen are sheltered from enzymatic decomposition or degradation by extreme values of pH or redox potential. Additionally, EVs enable the simultaneous delivery of various virulence factors, allowing for the induction of synergistic pathological effects [17]. Therefore, EVs are considered a crucial element of intercellular pathogen-host interactions [18]. Selective transport of toxins and other lytic factors into host cells via EVs has been demonstrated for many microbial species [17,19]. Although H. pylori typically delivers VacA and CagA toxins to host cells using T5SS and T4SS, respectively, the vesiculation process is currently considered to be highly important in the transfer of these virulence factors [20,21,22,23].
The pathological effect of VacA and CagA toxins from H. pylori on gastric cells is well known and described [1,4,5]. On the other hand, studies on the transfer of these toxins via EVs represent a new research avenue that may deepen the understanding of phenomena underlying the development of diseases caused by this bacterium [22]. As demonstrated for various microbial species, antibiotic therapy may significantly impact the secretion and toxicity of microbial EVs against host cells [24,25,26,27,28,29]. Knowledge regarding the effects of antimicrobial substances on EVs produced by H. pylori is however currently in its infancy. To date, only three articles focusing on this topic have been published [30,31,32]. Two of these described the ability of H. pylori EVs to induce tolerance to the tested antimicrobials [30,31], while the third, conducted by our team, determined the effect of selected antibiotics on the physicochemical parameters and fatty acid profile of EVs from this bacterium [32].
Building on our previous research and aiming to address the knowledge gap, this article aims to assess the effect of antibiotics on the toxicity of H. pylori EVs towards gastric cells (Figure 1). Using a range of experimental techniques, we were the first to demonstrate that antibiotics modify the cytopathic effect induced by EVs of H. pylori in gastric cells.
2. Results
For our current studies on the cytotoxicity and cytopathic effect of EVs toward gastric cells, we selected the clinical H. pylori 3CML strain. This choice was dictated by two factors. Firstly, this strain is multidrug-resistant (resistant to clarithromycin [CLR], metronidazole [MTZ], and levofloxacin [LEV]) [33] and therefore provides an excellent model for testing the effect of subinhibitory (clinically ineffective) antibiotic concentrations on the activity of EVs against gastric cells. Secondly, among all the characterized H. pylori strains from our collection, this isolate is equipped with one of the highest adaptive properties under antibiotic pressure, as evidenced by the EV secretion, biofilm production, and coccoid transformation [33,34]. At the same time, from our strains with the highest adaptive capacities, H. pylori 3CML presents the highest level of EV production, both with and without antibiotic exposure [32].
Our experiments began by analyzing the toxin profile of H. pylori 3CML. We demonstrated that this strain is capable of producing the CagA toxin (cagA+) and possesses one of the more toxic variants of VacA (vacAs1m2) (Figure 2A). Next, using reverse transcription-quantitative polymerase chain reaction (RT-qPCR), we determined how exposure to subinhibitory concentrations of antibiotics affects the expression of genes encoding these two toxins. We observed an interesting phenomenon, consisting of an increased cagA expression and decreased vacA expression in bacterial cells exposed to CLR, whereas the opposite was noticed for MTZ or LEV (Figure 2B,C). This suggests that subinhibitory concentrations of classically used antibiotics can significantly modulate the toxigenicity of this pathogen.
Knowing the profile of toxins and the effect of antibiotics on their expression, we managed to isolate and characterize EVs of H. pylori 3CML according to our established methodology of culturing bacteria in conditions devoid of animal-derived EVs in serum applied to liquid culture media [32]. Firstly, we demonstrated that the subinhibitory antibiotic concentrations did not significantly affect the growth of H. pylori 3CML in these conditions (Figure S1). Secondly, based on nanoparticle tracking analysis (NTA), we confirmed a high EV secretion capacity for this strain. The concentration and the mean size of EVs were in the range of 1.6–2.3 × 10^11^ EVs/mL and 103–126 nm, respectively (Figure 3). Interestingly, we observed a pattern of increasing EV dimensions with decreasing total concentration (1.6 × 10^11^ EVs/mL and 126 nm in CLR-treated bacteria and 2.3 × 10^11^ EVs/mL and 103 nm for those exposed to MTZ). The presence of EVs in the samples and the absence of contaminating structures (e.g., flagella) was additionally confirmed by us using transmission electron microscopy (TEM) (Figure 3).
After characterizing the strain’s toxin profile and isolating its EVs, we proceeded to the first stage of the main experimental phase by determining the cytotoxicity of these structures towards AGS cells. As expected, we observed a dose-dependent toxic effect (p < 0.05) (Figure 4). Among the EV variants tested, those derived from native bacteria (unexposed to antibiotics) or CLR-treated ones exhibited a higher level of cytotoxicity than EVs isolated from the strain treated with MTZ or LEV (p < 0.05). Regardless of the EV variant used, however, doses of 3 × 10^10^ and 10^10^ caused a statistically significant decrease in AGS viability in all samples (down to 45–79.6% and 45.2–90%, respectively; Figure 4A,B). At the lowest dose (6 × 10^8^), only EVs from native and CLR-treated bacteria maintained their cytotoxicity (survival rates of 55.9% and 66.4%, respectively; Figure 4D).
To further increase our understanding of the effects of the studied H. pylori EVs on gastric cell physiology, we performed a series of fluorescent microscopic observations and selective staining of cellular components (Figure 5). Analyzing the post-microscopic data, we observed that EVs used at the highest dose (3 × 10^10^) induced a significant increase in the fluorescence of the analyzed structures (Figure 5A,C). This phenomenon also occurred when using reference substances (chloroquine—a vacuolization control [35], and colchicine—a hummingbird phenotype control [36]; Figure 5B,D), but not with lower EV doses (Figure 5A,C). Linking the facts together, we hypothesized that EVs at a dose of 3 × 10^10^ might induce changes in the permeability of AGS cell membranes and stimulate an increase in the uptake of fluorescent dyes from the extracellular environment. Because changes in host membrane permeability are characteristic of both VacA and CagA [37,38], we closely examined the phenotypic changes in AGS cells exposed to 3 × 10^10^ EVs of H. pylori 3CML. Indeed, we confirmed that many of them showed changes visually resembling vacuolation and/or elongation (Figure 5E).
In the third stage of the main experimental phase, we decided to confirm the observations made with fluorescence microscopy. To achieve this, we used light microscopy and a neutral red staining—a dye targeting vacuoles [39] (Figure 6). This type of monochromatic image analysis facilitated the easier interpretation of the results. In all samples, we noted an augmented absorption of neutral red (Figure 6A,C). Simultaneously, we observed that AGS cells exposed to EVs coming from MTZ- or LEV-treated H. pylori 3CML had higher levels of absorbed dye (0.52 and 0.46, respectively) than after exposure to EVs derived from native bacteria (0.43) or those treated with CLR (0.35). Additionally, based on the shape, we categorized AGS cells into physiological and elongated ones, and confirmed the ability of the analyzed EVs to induce the hummingbird phenotype (Figure 6B,C). In this context, the highest level of cellular elongation was observed for EVs of CLR-treated bacteria (40%), while the lowest for EVs of LEV-treated ones (24.5%). The data of the current experimental stage correlate well with toxins’ gene expression in H. pylori 3CML (Figure 2B,C), suggesting that EVs secreted by this strain contain toxins whose levels and ability to induce cytopathic effects in AGS cells are modulated by subinhibitory antibiotic concentrations.
In the final step, we aimed to establish whether the EVs used were capable of stimulating proinflammatory cytokines in the AGS line and whether antibiotics could influence this effect. Indeed, we confirmed an increase in IL-8 levels in the post-culture supernatant of AGS cells exposed to all EV variants (Figure 7). Furthermore, we observed a strong correlation between the MTT-based viability analyses (Figure 4) and the present data of IL-8 secretion levels (Figure 7). In this regard, the highest proinflammatory induction was observed for EVs isolated from native H. pylori 3CML cells, followed by those exposed to CLR, then both MTZ and LEV.
3. Discussion
The ineffectiveness of antimicrobial therapies is a long-standing challenge for the healthcare sector. The latest and most comprehensive study analyzing global mortality over the past three decades revealed this to be one of the greatest problems [40]. In 2021 alone, nearly 5 million deaths were attributed to infections caused by antibiotic-resistant bacteria, and this number is expected to rise to 8 million by 2050. On a worldwide scale, the economic burden of antibiotic resistance costs hundreds of trillions of dollars annually [41]. The issue of drug resistance and its associated costs is also very current for infections caused by H. pylori. A recent international, multicenter study showed that antibiotic resistance of H. pylori to the most commonly used and effective antimicrobials (CLR, MTZ, and LEV) is increasing [42]. The prevalence of antibiotic resistance in this pathogen varies across the world, although it is significantly higher in developing countries and is greatly fueled by the overuse and misuse of antimicrobial drugs [43]. In this context, a strong correlation between the level of outpatient consumption of macrolides, fluoroquinolones, and nitroimidazoles in the treatment of other microbial diseases and the development of H. pylori resistance is noticed [42,44]. Furthermore, we believe that this phenomenon may play a significant role not only in generating H. pylori strains resistant to a given class of antimicrobial compounds, but also in modulating their virulence traits once such resistance has developed.
Antibiotics induce a number of changes in the physiology and virulence of microorganisms [45,46], primarily when used at subinhibitory concentrations [47]. These modifications can affect both their survival capacity (e.g., biofilm formation) and toxicity (e.g., invasiveness, production of toxins or lytic enzymes) [45,46,47]. Although the impact of antibiotics on microbial vesiculation remails a relatively unexplored research area, there are different scientific reports demonstrating the effect of antimicrobials on the quantity and/or toxicity of EVs towards host cells [24,25,26,27,28,29]. In line with this, we believe that antibiotic-resistant H. pylori constitutes an excellent research model for determining the significance of this phenomenon. Because of the ability to chronically colonize the stomach [4], drug-resistant strains of this pathogen can be repeatedly exposed to subinhibitory antibiotic concentrations, which in turn may reduce or enhance the cytotoxicity of their EVs towards gastric cells (Figure 1).
In the current study, we determined the effect of EVs derived from a multidrug-resistant H. pylori strain exposed to subinhibitory concentrations of CLR, MTZ or LEV on a number of physiological characteristics of gastric cells. The methodology of the current EV isolation was consistent with that developed by us previously [32]. For this purpose, we used bacteriological culture in serum devoid of animal-borne EVs and estimated precisely the concentration of isolated EVs by NTA. This approach is consistent with good practices recommended by others [48] and certainly contributed to the reliability of our data. According to the data of Zavan et al. [49], a 3-day incubation of H. pylori contributes to the highest production of EVs and the greatest abundancy of virulence factors carried in these structures, therefore we limited our analysis only to EVs coming from this growth stage.
In the first stage of the experiments, we observed that the cytotoxicity of H. pylori EVs toward gastric cells increased with increasing doses of these structures, which is convergent with results from other studies [50,51]. Interestingly, we observed that the highest level of toxicity was observed for EVs derived from native bacteria (untreated with antibiotics), then exposed to CLR, and finally to MTZ or LEV. At the same time, the data from the cytotoxicity assays correlated well with our observations on the capacity of these structures to induce IL-8 in gastric cells. Although the exact cause of this phenomenon is unknown, literature data allowed us to explain its source. Antibiotics are able to stimulate microbial vesiculation, while the intensity of this process depends largely on the molecular mechanism of their action [52]. Antimicrobials that target nucleic acids, including MTZ or LEV, are potent inducers of vesiculation [24,53,54], whereas substances that inhibit translation (e.g., CLR) do not cause significant changes in the EV synthesis [24]. Our team made similar observations in the previous study on H. pylori [32]. Therefore, we believe that increased EV production during exposure of H. pylori to MTZ or LEV may contribute to a stoichiometric reduction in the virulence factors deposited in EVs and a decrease in their cytotoxicity. Unlike the previous two antibiotics, exposure of H. pylori to CLR does not significantly affect vesiculation and thus the level of cytotoxicity of EVs derived from these conditions is similar with EVs of native bacteria.
Considering that CagA and VacA carried in H. pylori EVs may be responsible for the cytotoxic effect [20,21,22,23], we decided to determine the phenotypic changes occurring in gastric cells exposed to these structures. We noticed that at the highest doses, all types of H. pylori EVs were capable of inducing both cell elongation (a hummingbird phenotype) and vacuolization. The magnitude of this phenomenon was interrelated with the toxins’ gene expression levels, confirming our suspicion that these two toxins are of the highest importance in the cytotoxicity of H. pylori EVs. Nevertheless, we are aware that EVs produced by H. pylori may carry dozens of different virulence factors [49,55,56]. These additional virulence determinants include primarily metabolic enzymes, urease subunits, and adhesins (lipopolysaccharide subunits and outer membrane proteins), which are responsible for nutrient acquisition, neutralization of the acidic environment, and uptake of EVs into host cells, respectively. Therefore, although it appears that the cytotoxicity of the examined EVs was substantially associated with the action of VacA and CagA toxins, the potential existence of synergistic action of several virulence factors cannot be definitively excluded. In line with this, we plan to expand our studies in the future to include proteomic analyses of H. pylori EVs and to determine how exposure of H. pylori to subinhibitory antibiotic concentrations affects the cytotoxicity and cytopathic effect of whole bacterial cells against gastric cells. All of the above will help us to understand better the intricate interactions in the antibiotics-H. pylori-host axis.
4. Materials and Methods
4.1. Microbiological Experiments
4.1.1. Storage and Revival of Bacteria
The methodology for storing and maintaining H. pylori was in accordance with our established procedure [33,34]. A multidrug-resistant clinical H. pylori 3CML strain, belonging to the collection of the Department of Microbiology at Wroclaw Medical University, was chosen for the present study. The strain was stored at −80 °C in Tryptic Soy Broth (TSB; Oxoid, Dardilly, France) supplemented with 30% glycerol until use in experiments. To reactivate the strain, the frozen stock of bacteria was plated on Columbia agar (Difco, Lublin, Poland) enriched with 10% horse blood (GRASO, Starogard Gdanski, Poland) and incubated for 3 days at 37 °C under microaerophilic conditions (Genbox microaer kits, BioMerieux, Marcy l’Étoile, France). Following this, the strain was subcultured onto fresh agar plates and incubated under the same conditions for an additional 3 days. Then, the strain was proceeded to subsequent experimental stages.
4.1.2. Genetic Analyses
The analysis of H. pylori 3CML toxin profile was performed using the PCR method and selective primers designed in other study [57]. Genetic material obtained from agar-based bacterial cultures was subjected to DNA isolation using the Syngen DNA Mini Kit (Syngen Biotech, Wrocław, Poland), according to the manufacturer’s instructions. Details of the target gene, amplicon size, primer names and sequences are provided in Table 1. For each PCR reaction, 1–2 μg of DNA was added to a 25-μL PBS-based reaction mixture containing 1 µM of each primer (forward and reverse; Oligo.pl, IBB, Warsaw, Poland), 1 mM MgCl_2_ (ThermoFisher Scientific, Waltham, MA, USA), 1.25 U of a DreamTaq DNA polymerase (ThermoFisher Scientific, Waltham, MA, USA), and 200 μM of each dNTP (Oligo.pl, IBB, Warsaw, Poland). The PCR amplification protocol was performed using a PTC-200 Thermal Cycler (Bio-Rad Laboratories, Hercules, CA, USA) and included an initial denaturation at 95 °C for 3 min, followed by 35 cycles of denaturation at 95 °C for 30 s, annealing for 30 s (see Table 1 for specific temperatures), and extension at 72 °C for 30 s. A final polymerization step was carried out at 72 °C for 5 min. PCR products were separated by electrophoresis on a 1.8% agarose gel. Bands were visualized with ethidium bromide staining and analyzed using the Gel-Doc 2000 software (Bio-Rad Laboratories, Hercules, CA, USA), and their size was estimated using a FastGene 50 bp DNA Marker (50 bp–1500 bp; NIPPON Genetics, Düren, Germany). Genetic material from H. pylori J99 and Tx30a cultures was used as a control when estimating a toxigenic profile of the analyzed H. pylori strain (cagA+, vacA s1m1 and cagA-, vacA s2m2, respectively).
The assessment of the effect of antibiotics on the expression of H. pylori 3CML toxin genes was performed using RT-qPCR and primers designed earlier by others (Table 2) [58,59]. Bacteria were grown in 24-well titration plates (Bionovo, Legnica, Poland) filled with Brain-Heart Infusion broth (BHI; Oxoid) and 10% EV-depleted fetal calf serum (FCS^-EVs^; Gibco, Paisley, Scotland, UK). Each well of the 24-well titration plate was loaded with 1.5 mL of the culture medium, a bacterial suspension (10^8^ CFU/mL), and ¼× minimal inhibitory concentration (¼× MIC) of one of the tested antibiotics (CLR, MTZ or LEV; all from Merck, St. Louis, MO, USA). Wells containing only the bacterial suspension (without antibiotic) were used as a positive control. The plates were incubated for 3 days at 37 °C under microaerophilic conditions with shaking at 100 rpm. After this period, the bacterial cells were centrifuged, the supernatant was discarded, and the remaining pellet was used for the isolation of genetic material. Total RNA was extracted using the RNeasy Mini Kit (Qiagen, Hilden, Germany) and complementary DNA (cDNA) was synthesized using the iScript Reverse Transcription Supermix (Bio-Rad, Hercules, CA, USA). RNA concentration and purity were assessed with a NanoDrop ND-1000 spectrophotometer (ThermoFisher Scientific, Waltham, MA, USA), checking the 260/280 and 260/230 absorbance ratios to ensure there was no contamination from proteins or solvents. RT-qPCR was performed in 20 μL reaction volumes using the SsoAdvanced Universal SYBR Green Supermix (Bio-Rad, Hercules, CA, USA) on an Applied Biosystems 7500 Fast Real-Time PCR System (Foster City, CA, USA). The thermal cycling conditions were as follows: polymerase activation at 95 °C for 15 min, then 45 cycles of denaturation at 95 °C for 15 s, annealing at 60 °C for 20 s, and extension at 72 °C for 20 s. Gene expression levels were normalized to a reference gene (16S rRNA), and the relative mRNA expression was calculated using the ΔΔCt method. Each reaction was run in triplicate (n = 3).
4.1.3. Isolation of EVs and Their Characterization
The methodology for production and isolation of H. pylori EVs was consistent with that established in our previous research [32].
Bacteria were grown in 24-well titration plates filled with BHI and 10% FCS^-EVs^. Each well of the 24-well titration plate was loaded with 1.5 mL of the culture medium, a bacterial suspension (10^8^ CFU/mL), and ¼× MIC of one of the tested antibiotics (CLR, MTZ or LEV). Wells containing only the bacterial suspension (without antibiotic) and only culture medium (without bacteria) were used as positive and negative controls, respectively. The plates were incubated for 3 days at 37 °C under microaerophilic conditions with shaking at 100 rpm. The optical density of bacteria was evaluated spectrophotometrically using an Asys UVM 340 microplate reader (Biochrom Ltd., Cambridge, UK) at OD_600_ each day. The values of negative controls (culture medium without bacteria) were subtracted from the measurements of the experimental samples. Three biological repetitions of this analysis were performed (n = 3). After a 3-day culture, the samples were directed for the EV isolation procedure.
EVs were isolated using a polymer-based method with the Total Exosome Isolation Kit (from cell culture media; ThermoFisher Scientific, Waltham, MA, USA), following the manufacturer’s instructions. In brief, bacterial cultures were first centrifuged at 2000× g for 30 min and the resulting supernatant was filtered through a 0.45 µm cellulose acetate filter (Corning, New York, NY, USA). The filtrate was then combined with the isolation reagent at a 2:1 ratio, vortexed, and incubated overnight at 4 °C. Subsequently, samples were centrifuged at 10,000× g for 60 min, the supernatant was discarded, and the EV-containing pellet was resuspended in PBS (Merck, St. Louis, MO, USA). To ensure the absence of bacterial contamination, 10 µL of the EV solution was plated on Columbia agar supplemented with 10% horse blood and incubated for 7 days at 37 °C under microaerophilic conditions. EV preparations were stored at −80 °C until further analysis.
The concentration, mean size, and size distribution of EVs were assessed using NTA with the NanoSight NS500 system (Malvern Instruments, Malvern, UK), employing a 405 nm blue diode laser as the light source. Measurements were carried out at an ambient temperature of approx. 23 °C. Each biological sample was analyzed through nine independent runs and the average of these values was reported. As particle concentrations exceeded the detection range, samples were diluted with PBS and the dilution factor was incorporated into the final EV concentration calculations.
The ultrastructure of EVs was evaluated using TEM. EV samples suspended in PBS were applied to Formvar-carbon-coated copper grids (200 mesh, Ted Pella, Redding, CA, USA) and fixed with 2% paraformaldehyde (ThermoFisher Scientific, Waltham, MA, USA), and 2.5% glutaraldehyde (Serva Electrophoresis, Heidelberg, Germany). Grids were then counterstained with Uranyless (Electron Microscopy Sciences, Hatfield, PA, USA), followed by 3% lead citrate (Electron Microscopy Sciences, Hatfield, PA, USA). Finally, they were embedded in 0.13% methyl cellulose (viscosity 25 cP; Merck, St. Louis, MO, USA) and examined using a JEM-1011 TEM (Jeol, Tokyo, Japan) operated at an accelerating voltage of 80 kV.
4.2. Cell Line Experiments
4.2.1. Storage and Revival of the Cell Line
The methodology for storing and maintaining cell line cultures was in accordance with the procedure used by us previously [60], with minor modifications. Human Gastric Adenocarcinoma AGS cells (CRL-1739; ATCC, Manassas, VA, USA) were kept in liquid nitrogen in cryopreservation medium containing DMEM medium (ThermoFisher Scientific, Waltham, MA, USA), 10% fetal bovine serum (FBS; Corning, New York, NY, USA), 1% penicillin-streptomycin (100 U/mL–100 µg/mL; Corning, New York, NY, USA), and an addition of 10% dimethyl sulfoxide (DMSO; Merck, St. Louis, MO, USA). Cells were frozen in a controlled manner to achieve a cooling rate of approximately −1 °C/min and then were transferred to liquid nitrogen (−196 °C). To revive them, the cryotube with the frozen cells was rapidly thawed in a water bath at 37 °C for approx. 1–2 min. The content was transferred to a conical Falcon tube (Corning, New York, NY, USA) containing 10 mL of pre-warmed culture medium without DMSO, centrifuged (1000 rpm, 5 min), and then the supernatant was removed. The cells were resuspended in the fresh medium and seeded into a T75 flask (Sarstedt, Nümbrecht, Germany) with the appropriate amount of the medium.
AGS cells were routinely cultured in DMEM medium supplemented with 10% FBS and 1% penicillin-streptomycin in an incubator maintaining at 37 °C, 5% CO_2_, and high humidity (Heracell 150i, ThermoFisher Scientific, Waltham, MA, USA). After reaching an appropriate confluence of 80–90%, the cells were detached from a T75 flask using 0.05% trypsin-EDTA (ThermoFisher Scientific, Waltham, MA, USA) and seeded into 24-well titration plates (TPP Techno Plastic Products, Trasadingen, Switzerland), 96-well titration plates (TPP Techno Plastic Products, Trasadingen, Switzerland) or 8-well cell culture slides (SPL Life Sciences, Pocheon, Republic of Korea), depending on the type of microscopic or functional evaluation performed.
4.2.2. Cytotoxicity
The cytotoxicity of H. pylori EVs towards AGS cells was assessed using the MTT assay [51]. AGS cells were seeded at a density of 5 × 10^3^ cells/well into 96-well titration plates containing 100 µL of DMEM enriched with 10% FBS and 1% penicillin-streptomycin. The cells were incubated under standard conditions (37 °C, 5% CO_2_) to allow for their adhesion and achievement of 80–90% confluence. After this, the AGS cells were treated with H. pylori EVs derived from bacteria non-exposed to any antibiotic (a native sample) or those treated with ¼× MIC of CLR, MTZ, or LEV. In each case, various doses of H. pylori EVs were used: 3 × 10^10^, 10^10^, 2.5 × 10^9^, and 6 × 10^8^. The negative control constituted a PBS-resuspended sample from a BHI broth with 10% FCS^-EVs^ without bacteria, which was subjected to the similar EVs isolation steps as described in Section 4.1.3. The volume of the negative controls was adequate to the volume of each tested sample. For all samples, an incubation continued for 24 h under standard culture conditions. Then, the medium in each case was replaced with 100 µL of fresh medium containing an additional 0.5 mg/mL of MTT (Merck, St. Louis, MO, USA). Plates were incubated for 3 h at 37 °C in the dark. After incubation, the medium was removed and the formazan was dissolved in 100 µL of DMSO. Absorbance was measured at 570 nm using an Asys UVM 340 microplate reader (Biochrom Ltd., Cambridge, UK). Results were expressed as the percentage viability compared to the negative controls. The tests were performed in three biological replications (n = 3).
4.2.3. Fluorescent Microscopy Observations
Analysis of phenotypic changes in cell lines using fluorescent microscopy and selective staining of cellular components was performed in line with others [61,62], with some modifications. AGS cells were seeded onto sterile glass slides with a 13 mm diameter (Bionovo, Legnica, Poland) at a density of 5 × 10^4^ cells/well. After this, the glass slides with the AGS cells were placed in wells of 24-well titration plates filled with 100 µL of DMEM enriched with 10% FBS and 1% penicillin-streptomycin, and incubated under standard conditions (37 °C, 5% CO_2_) to achieve 80–90% confluence. Next, the cells were then treated for 24 h with H. pylori EVs coming from bacteria non-exposed to any antibiotic (a native sample) or those treated with ¼× MIC of CLR, MTZ, or LEV. In each instance, a gradient of H. pylori EVs was applied: 3 × 10^10^, 10^10^, 2.5 × 10^9^, and 6 × 10^8^. The negative control constituted a PBS-resuspended sample from a BHI broth with 10% FCS^-EVs^ without bacteria, which underwent the same EVs isolation procedure as outlined in the “4.1.3. Isolation of EVs and their characterization” section. The volume of the negative samples corresponded to the volume of each tested sample. Additionally, chloroquine (0.1 µM; Merck, St. Louis, MO, USA) and colchicine (10 µM; Merck, St. Louis, MO, USA) were used as positive controls for vacuolization [63] and hummingbird phenotype [36], respectively, the concentrations of which were experimentally established by us during the validation stage of the methodology. After incubation, the medium was removed and the cells were washed three times with pre-warmed PBS. The cells were fixed with 4% buffered formaldehyde (Chempur, Piekary Śląskie, Poland) for 10 min at room temperature and then washed three more times with PBS to remove residual formaldehyde.
After fixing the cells, a staining solution containing 20 µL of PBS and 0.5 µL of each fluorescent dye: DAPI, FM 1-43, and phalloidin-conjugated Alexa Fluor 647 (all from ThermoScientific, Waltham, MA, USA) was added to each well. These dyes were used to visualize the cell nucleus [64], cell membrane (including vacuolization [65,66,67]), and cytoskeleton (including a hummingbird phenotype [68,69]), respectively. The cells were left for a 15 min-incubation in the dark and then were observed using a Carl Zeiss inverted fluorescence microscope (GmbH, Jena, Germany). Post-microscopic analysis of the obtained photographs was performed using the ImageJ software version 1.54j. In order to normalize differences in the cell density between analyzed samples, the fluorescence intensities of FM 1-43 and phalloidin-conjugated Alexa Fluor 647 were normalized toward the fluorescence of DAPI, and the values were presented as ratios of these two [70]. The tests were performed in three biological replications (n = 3).
4.2.4. Light Microscopy Observations
Analysis of phenotypic changes in cell lines using light microscopy was performed in line with others [71,72], with some modifications. AGS cells were seeded at a density of 3 × 10^4^ cells/well into 8-well cell culture slides filled with 200 µL of DMEM with 10% FBS and 1% penicillin-streptomycin. The cells were incubated under standard conditions (37 °C, 5% CO_2_) to achieve 80–90% confluence. The AGS cells were then exposed to 3 × 10^10^ of H. pylori EVs from bacteria non-exposed to any antibiotic (a native sample) or those treated with ¼× MIC of CLR, MTZ, or LEV. The negative control constituted a PBS-resuspended sample from a BHI broth with 10% FCS^-EVs^ without bacteria, which underwent the same EVs isolation procedure as outlined in Section 4.1.3. Chloroquine (0.1 µM; Merck, St. Louis, MO, USA) and colchicine (10 µM; Merck, St. Louis, MO, USA) were used as positive controls for vacuolization [63] and hummingbird phenotype [36], respectively. Incubation continued for 24 h under standard conditions. After culture, the cells were washed with PBS and incubated for 10 min with neutral red (400 µg/mL). Observations were performed with an Olympus BX50 microscope (Olympus, Tokyo, Japan). Post-microscopic analysis of the obtained photographs was performed using the ImageJ software version 1.54j. The degree of vacuolation was calculated by measuring the intensity of the absorbed red dye [71], the value of which was normalized to the surface area occupied by the observed cells. The hummingbird phenotype was defined when the ratio of the longest to shortest diameters of cells was greater than 2.5 [72]. All cells from the observation fields were counted. For both tested parameters three biological replicates with three technical replicates representing different observation fields were made (n = 9).
4.2.5. Stimulation of Proinflammatory Cytokines
A commercial IL-8 enzyme-linked immunosorbent assay kit (Human IL-8 Elisa Kit; ThermoFisher Scientific, Waltham, MA, USA) was used to determine the level of this cytokine in the supernatant of AGS cells [73,74]. IL-8 concentration was measured in culture medium collected from AGS cells exposed for 24 h to 3 × 10^10^ of H. pylori EVs (for culture conditions, see Section 4.2.4). Analyses were performed according to the manufacturer’s instructions. The obtained data were normalized to the absorbance values acquired in the MTT assay. In each case, the values obtained for the tested samples were subtracted from the negative control results. The tests were performed in three biological replications (n = 3).
4.3. Statistical Analysis
Statistical analyses were conducted using GraphPad Prism version 10 (GraphPad Software, San Diego, CA, USA). The Shapiro–Wilk test was applied to assess the normality of data distribution. As the data followed a normal distribution, subsequent analyses were performed using one-way or two-way ANOVA. A p-value of less than 0.05 was considered indicative of statistical significance.
5. Conclusions
In this study, we demonstrated that antibiotics used at subinhibitory concentrations toward H. pylori affect the expression of cagA and vacA and modulate the cytotoxicity and cytopathic effects of its EVs on gastric cells. These pathological processes induced by EVs of antibiotic-treated bacteria are lower compared to those of EVs from native (antibiotic-unexposed) cells. This suggests that short-term exposure of H. pylori to subinhibitory antibiotic concentrations does not translate into exacerbation of its EVs-driven virulence. However, extensive research in this area is undoubtedly needed to confirm these observations.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Malfertheiner P. Megraud F. Rokkas T. Gisbert J.P. Liou J.M. Schulz C. Gasbarrini A. Hunt R.H. Leja M. O’Morain C. Management of Helicobacter pylori Infection: The Maastricht VI/Florence Consensus Report Gut 2022711724176210.1136/gutjnl-2022-32774535944925 · doi ↗ · pubmed ↗
- 2Johnson K.S. Ottemann K.M. Colonization, Localization, and Inflammation: The Roles of H. pylori Chemotaxis In Vivo Curr. Opin. Microbiol.201841515710.1016/j.mib.2017.11.01929202336 PMC 5862749 · doi ↗ · pubmed ↗
- 3Mebuge D.L. Noel R.J. Gold B.D. Helicobacter pylori in Children: An Individualized Approach to a Worldwide Disease Curr. Treat. Options Pediatr.2025111310.1007/s 40746-025-00328-6 · doi ↗
- 4Malfertheiner P. Camargo M.C. El-Omar E. Liou J.M. Peek R. Schulz C. Smith S.I. Suerbaum S. Helicobacter pylori Infection Nat. Rev. Dis. Prim.202391910.1038/s 41572-023-00431-837081005 PMC 11558793 · doi ↗ · pubmed ↗
- 5Elbehiry A. Marzouk E. Aldubaib M. Abalkhail A. Anagreyyah S. Anajirih N. Almuzaini A.M. Rawway M. Alfadhel A. Draz A. Helicobacter pylori Infection: Current Status and Future Prospects on Diagnostic, Therapeutic and Control Challenges Antibiotics 20231219110.3390/antibiotics 1202019136830102 PMC 9952126 · doi ↗ · pubmed ↗
- 6He J. Liu Y. Ouyang Q. Li R. Li J. Chen W. Hu W. He L. Bao Q. Li P. Helicobacter pylori and Unignorable Extragastric Diseases: Mechanism and Implications Front. Microbiol.20221397277710.3389/fmicb.2022.97277735992650 PMC 9386483 · doi ↗ · pubmed ↗
- 7Chauhan N. Tay A.C.Y. Marshall B.J. Jain U. Helicobacter pylori Vac A, a Distinct Toxin Exerts Diverse Functionalities in Numerous Cells: An Overview Helicobacter 201924 e 1254410.1111/hel.1254430324717 · doi ↗ · pubmed ↗
- 8Foegeding N.J. Caston R.R. Mc Clain M.S. Ohi M.D. Cover T.L. An Overview of Helicobacter pylori Vac A Toxin Biology Toxins 2016817310.3390/toxins 806017327271669 PMC 4926140 · doi ↗ · pubmed ↗