Molecular insights into nucleocapsid assembly and transport in Marburg and Ebola viruses
Yuki Takamatsu, Olga Dolnik, Ai Hirabayashi, Kenta Okamoto, Tomomi Kurashige, Hu Shangfan, Catarina Oda Harumi, Takeshi Noda

TL;DR
This study reveals how nucleocapsids in Marburg and Ebola viruses assemble and move, identifying a key protein motif that could be targeted for antiviral treatments.
Contribution
The study identifies the conserved PPxPxY motif in NP as a critical regulator of nucleocapsid formation and viral replication in filoviruses.
Findings
NP, VP35, and VP24 are sufficient to form transport-competent nucleocapsid-like structures in Marburg and Ebola viruses.
The PPxPxY motif in NP is essential for NP-VP30 interactions and viral replication across filoviruses.
VP30 can support replication in heterologous systems despite protein incompatibility between Marburg and Ebola viruses.
Abstract
Live-cell imaging enables visualization of the spatiotemporal dynamics of signals in cells. Intracytoplasmic movement of nucleocapsids is crucial during the life cycle of enveloped viruses; however, the molecular mechanisms governing their assembly and transport are not fully understood. Using a Marburg virus (MARV) live-cell imaging system, we identified three nucleocapsid proteins—nucleoprotein (NP), VP35, and VP24—that are necessary and sufficient to form transport-competent nucleocapsid-like structures (NCLSs). These findings are consistent with observations in Ebola virus (EBOV). Interestingly, despite incompatibility among these proteins, VP30 interacts with nucleocapsid proteins from both MARV and EBOV, supporting viral transcription and replication in heterologous systems. Furthermore, we show that the conserved PPxPxY motif at the C-terminus of NP regulates NP-VP30 interactions…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
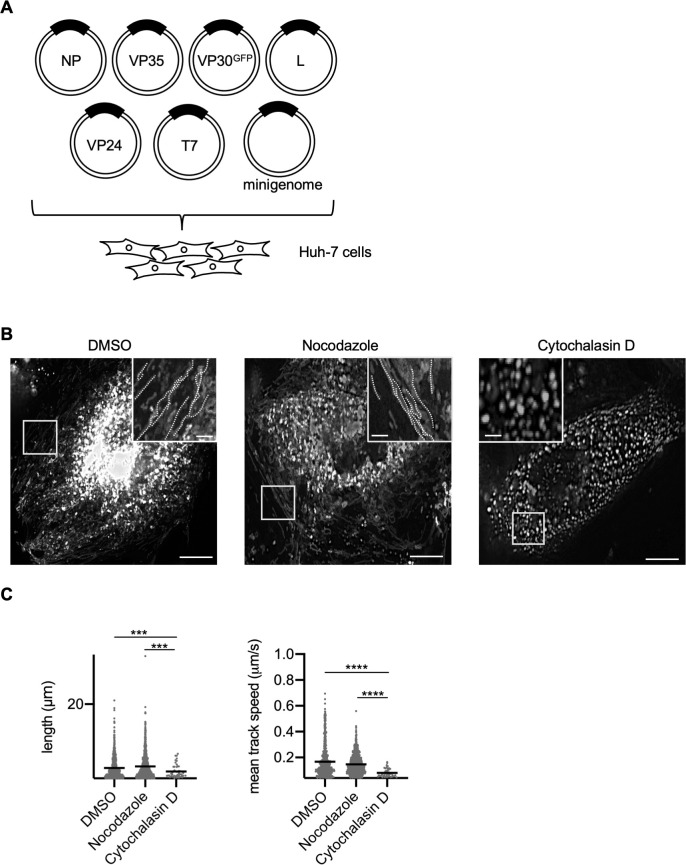 Fig 1
Fig 1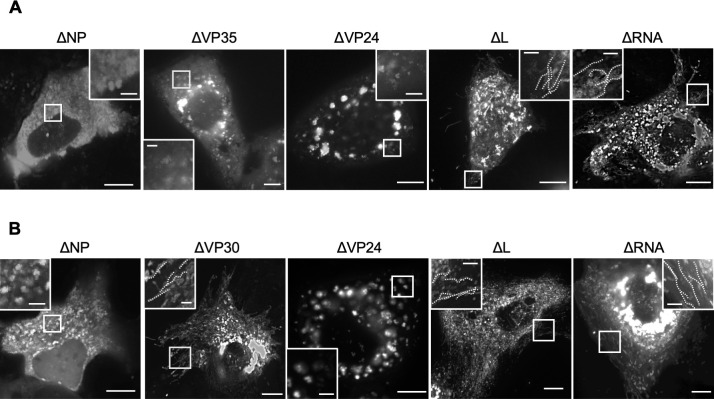 Fig 2
Fig 2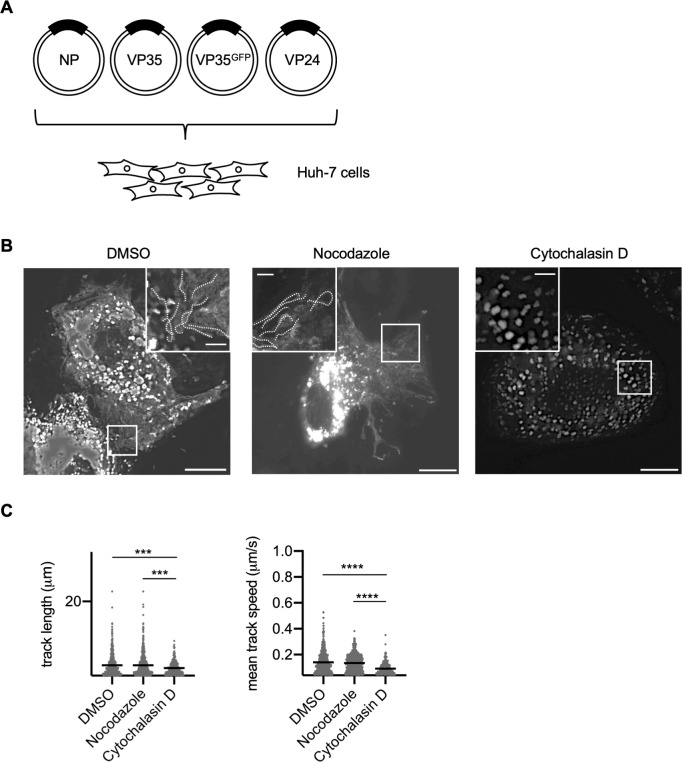 Fig 3
Fig 3 Fig 4
Fig 4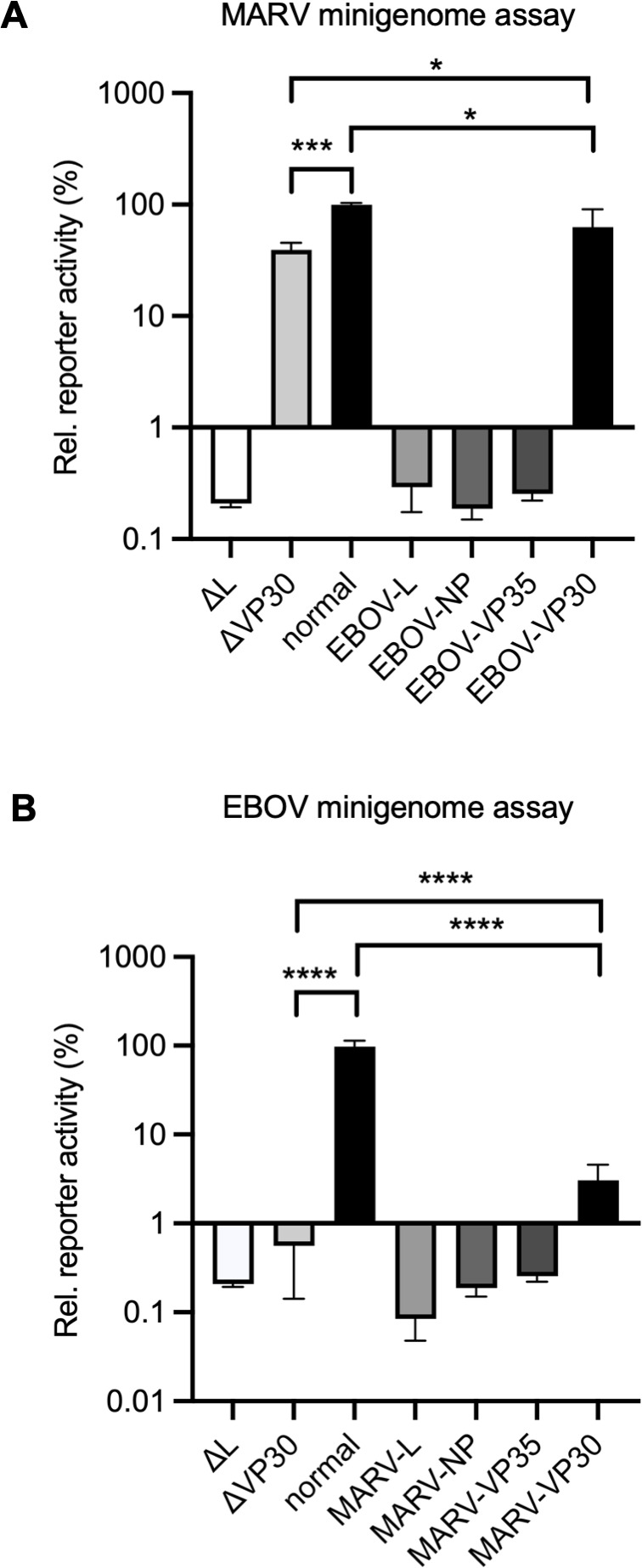 Fig 5
Fig 5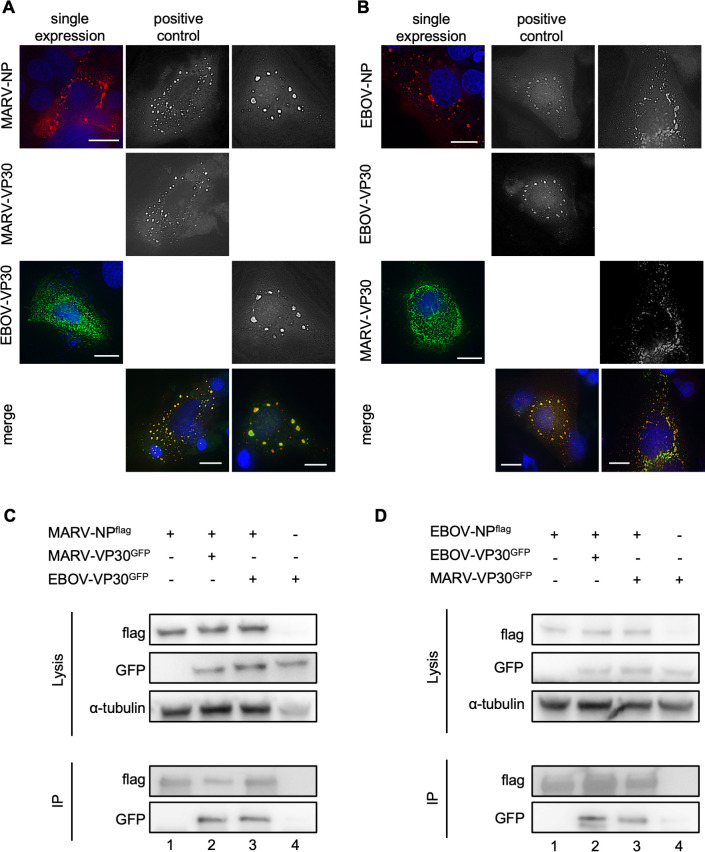 Fig 6
Fig 6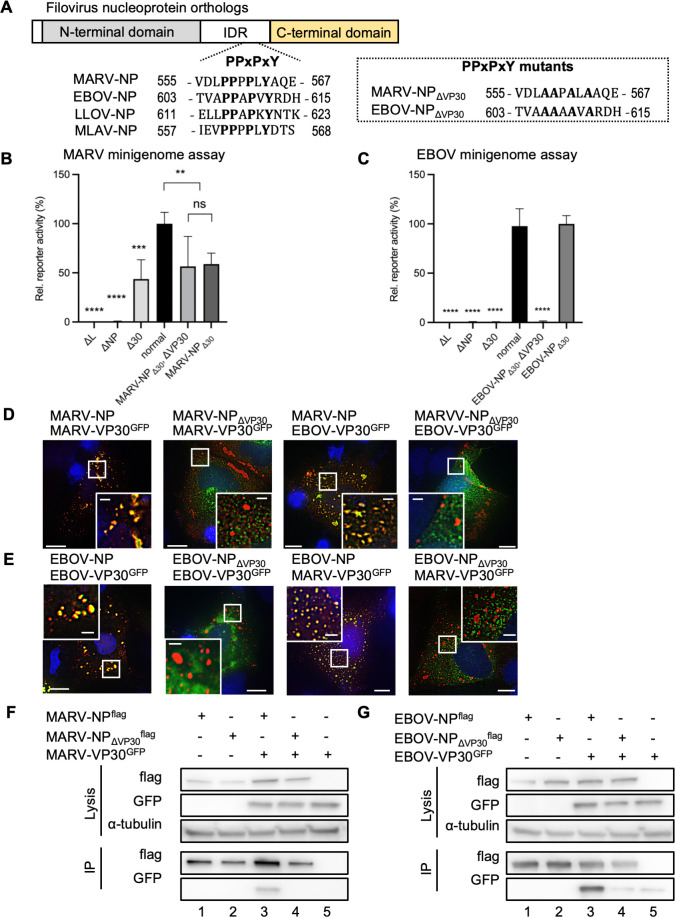 Fig 7
Fig 7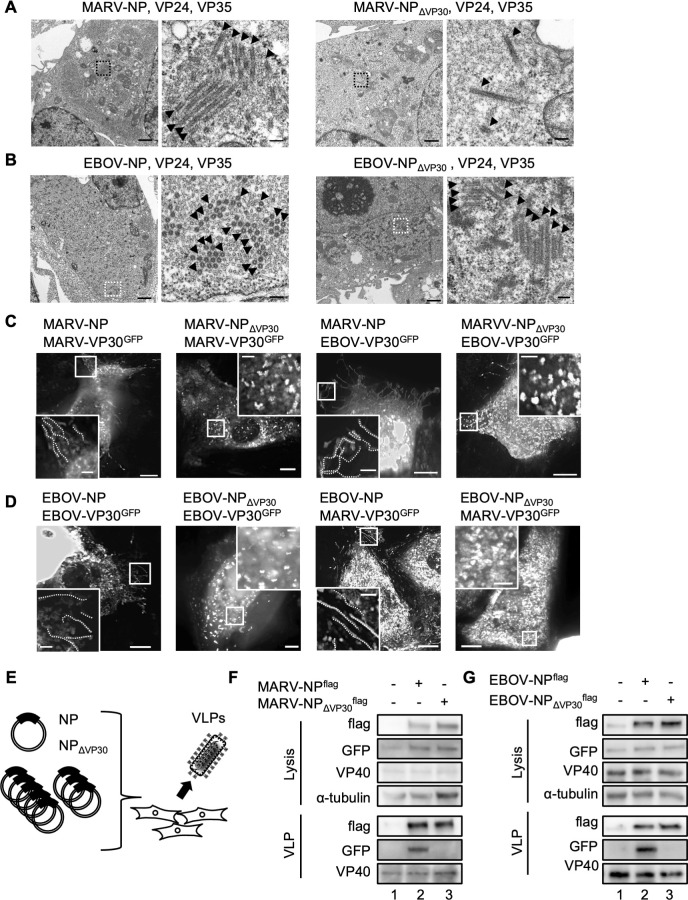 Fig 8
Fig 8 Fig 9
Fig 9- —Japan Society for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100001691
- —Asahi Glass Foundationhttp://dx.doi.org/10.13039/100007684
- —Joint Usage/Research Center on Tropical Disease
- —Joint Usage/Research Center program of Institute for Frontier Life and Medical Sciences Kyoto University
- —Japan Agency for Medical Research and Developmenthttp://dx.doi.org/10.13039/100009619
- —Japan Society for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100001691
- —Japan Society for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100001691
- —Joint Usage/Research Center program of Institute for Frontier Life and Medical Sciences Kyoto University;
- —Joint Usage/Research Center on Tropical Disease, Nagasaki University
- —Takeda Science Foundationhttp://dx.doi.org/10.13039/100007449
- —Japan Agency for Medical Research and Developmenthttp://dx.doi.org/10.13039/100009619
- —Ichiro Kanehara Foundation for the Promotion of Medical Sciences and Medical Carehttp://dx.doi.org/10.13039/501100003837
- —MSD Life Science Foundation, Public Interest Incorporated Foundationhttp://dx.doi.org/10.13039/501100013236
- —Japan research foundation for clinical pathology
- —Terumo Foundation for Life Sciences and Artshttp://dx.doi.org/10.13039/501100008670
- —NOVARTIS Foundation (Japan) for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100011746
- —Kurozumi Medical Foundationhttp://dx.doi.org/10.13039/501100008660
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral Infections and Outbreaks Research · Viral Infections and Vectors · Hepatitis B Virus Studies
INTRODUCTION
The viral genome is encapsulated by proteins, forming nucleocapsids (NCs), to protect it from recognition by cellular defense mechanisms (1). In Mononegavirales virus infection, the newly synthesized nucleocapsid associated with viral RNA-dependent RNA polymerase complex is transported to the plasma membrane for virion formation and release (2, 3). Cryo-electron microscopy has revealed the high-resolution helical structure of RNA-bound nucleoprotein (NP) of Ebola virus (EBOV), Marburg virus (MARV), Cueva virus, Nipah virus, and measles virus (4–8), which provides the molecular mechanisms driving and stabilizing the basic structure of assembled nucleocapsids. However, the complex structure of nucleocapsid and the molecular mechanisms of NP-phosphoprotein (P) association have been largely unrevealed. Measles virus P is mostly disordered, while three distinct interacting sites between it and NP contribute to efficient viral genome transcription (9). In paramyxoviruses, polymerase and nucleoprotein are connected via the P protein, and a flexible conformational change may occur during RNA processing (9, 10). The P protein is conserved in mononegaviruses, the roles of which are assumed to be divided on VP30 and VP35 in filoviruses (11–16), whereas the interplay of nucleocapsid proteins during nucleocapsid assembly has been largely concealed.
MARV and EBOV belong to the family Filoviridae and have an approximately 19 kb non-segmented, single-stranded, negative-sense RNA genome. Although temporal epidemics have been reported over several years in central-western Africa, the first MARV epidemic was reported following an EBOV epidemic in 2021 in Guinea, Western Africa (17, 18), where the largest ever EBOV epidemic occurred during 2014–2016 with over 11,000 deaths (19). Since antiviral therapeutics for MARV and EBOV diseases are not well established, it is imperative to understand the molecular mechanisms of viral replication to establish countermeasures against these viruses. In this regard, revealing the intracellular dynamics of filoviruses is crucial because nucleocapsid assembly and transport are prerequisites for virion formation.
Oligomerized NPs lead to the formation of inclusion bodies, which are sites for viral genome transcription, replication, and nucleocapsid synthesis (13, 20–23). The core structure of filovirus nucleocapsids includes a nucleocapsid-like structure (NCLS) composed of NP, encapsulating single-stranded viral genomic RNA (24, 25), together with the nucleocapsid protein VP24 and the polymerase cofactor VP35, both of which are essential structural elements that directly interact with NP to build a helical nucleocapsid approximately 800 nm–1,000 nm in length and 50 nm in diameter (20, 21, 24, 26). Viral polymerase L and transcription factor VP30 are also associated with the nucleocapsid (11, 27). An immunoelectron microscopy study of nucleocapsids demonstrated that NP, VP24, and VP35 are located from the center in this order, forming NCLS, and phosphorylation-mediated VP30 association was observed at the peripheral region of NCLSs (28, 29). Although the structural elements and viral protein properties are suggested to be highly conserved in filoviruses (24, 30), a detailed nucleocapsid protein interaction has yet to be clarified, even in a recent cryo-electron microscopy analysis (31, 32).
Here, we applied an EBOV live-cell imaging system to MARV and revealed the essential viral factors and compatibility of NC proteins during NC assembly and transport in MARV and EBOV. Moreover, a common and distinct machinery of NC formation was identified between the two viruses. Interestingly, the PPxPxY motif mediates NP-VP30 interactions in filoviruses; however, in EBOV, it had no significant effect on transcription or replication. In contrast, VP30 lost its ability to support MARV transcription and replication.
RESULTS
Intracellular transport of MARV NCLSs nucleocapsids
The EBOV viral proteins VP30 and VP35 were used to visualize nucleocapsid and NCLS transport in live-cell imaging systems (2). green fluorescent protein (GFP)-conjugated VP30 and VP35 were analyzed and confirmed to be equivalent to their wild type in expression levels, subcellular localization, minigenomic functions, and colocalization with other NCLS proteins (Fig. S1). Moreover, no differences in nucleocapsid transport characteristics were observed in MARV-VP30^GFP^- or MARV-VP35^GFP^-expressing cells (Fig. S2; Movies S1 and S2) in accord with previous reports in EBOV infection (3, 33).
We previously reported that MARV NCLS transport can be visualized using a virus-like particle (VLP) system expressing NP, VP35, VP24, L, VP40, glycoprotein (GP), and minigenome, together with VP30^GFP^ (34). Since VP40 and GP were not indispensable for EBOV NCLS transport (2), we transfected only MARV nucleocapsid components in Huh-7 cells, and live-cell imaging was performed from 20 h post-transfection (p.t.) (Fig. 1A). Smooth, long lines were observed in dimethyl sulfoxide (DMSO)- or nocodazole-treated cells, whereas they disappeared in cytochalasin D-treated cells (Fig. 1B). The frequency of NCLSs translocated to a distance of >5 µm, as well as the mean velocity of NCLS transport, was drastically decreased in cytochalasin D-treated cells (Fig. 1C). In summary, NCLSs formed with NPs, VP35, VP30, VP24, L, and the minigenome exhibited long-distance transport via actin filaments, with characteristics similar to those of MARV nucleocapsid transport.
*Live-cell imaging analysis of MARV nucleocapsid-like structure transport. (A) Schematic representation of the experimental settings for MARV NCLS transport. Huh-7 cells were transfected with plasmids encoding NP, L, VP35, VP24, MARV-specific minigenome, T7 polymerase, and VP30GFP. (B) The cells were observed from 20 h p.t. At 17 h p.t., different cytoskeleton-modulating drugs were added to the culture medium: 0.15% DMSO, 15 µM nocodazole, or 0.3 µM cytochalasin D, and the cells were incubated for an additional 3 h. The image shows the maximum-intensity projection of time-lapse images of cells, recorded for 3 min; images were captured every 3 s. The small boxed areas are enlarged at the four corners. Scale bars: 10 µm (scale bar in insets, 2 µm). The dotted lines indicate typical NCLS trajectories. (C) The detected signals were analyzed using Imaris software. The length of the NCLS trajectories and velocity of NCLS transport were evaluated. The y-axis represents the signal counts. Lines indicate the mean ± SD. Asterisks indicate statistical significance; ***P < 0.001 and ***P < 0.0001.
Identification of viral components necessary for MARV NCLS transport
Next, we applied a reductionist approach to identify viral factors required for NCLS transport. The experiments were repeated with the omission of each viral component (NP, VP35, VP24, L, or minigenome). Using live-cell imaging with MARV-VP30^GFP^ to track NCLS transport, we observed that transport-competent NCLSs were still formed even when polymerase L or the EBOV-specific minigenome was omitted. In contrast, the omission of NP, VP35, or VP24 resulted in failure to form transport-competent NCLSs (Fig. 2A). Next, the experiments were repeated using MARV-VP35^GFP^ to monitor the role of VP30 in NCLS transport. Although the presence of NP and VP24 is crucial for NCLS transport, VP30 was dispensable for NCLS transport (Fig. 2B). These data indicate that NP, VP35, and VP24 proteins are essential, whereas polymerase L, VP30, and the minigenome are not necessary for NCLS transport.
Determination of essential viral components for MARV NCLS transport. (A) Huh-7 cells were transfected with plasmids encoding VP30GFP, NP, VP35, VP24, L, MARV-specific minigenome, and T7 polymerase, except that one of the expression plasmids was omitted, as indicated. (B) Huh-7 cells were transfected with plasmids encoding VP35GFP, NP, VP30, VP35, VP24, L, MARV-specific minigenome, and T7 polymerase except that one of the expression plasmids was omitted, as indicated. Live-cell imaging analysis was started from 20 h p.t. The image shows the maximum-intensity projection of time-lapse images of cells recorded for 3 min; images were captured every 3 s. The small boxed areas are enlarged at the four corners. Scale bars: 10 µm (scale bar in insets, 2 µm). The dotted lines indicate typical NCLS trajectories.
Transport characteristics of NP, VP35, and VP24 forming NCLSs
To confirm that these three components are sufficient to form transport-competent NCLSs, live-cell imaging of NP, VP24, and VP35 together with VP35-GFP-expressing cells was started at 20 h p.t. (Fig. 3A) Here again, we demonstrated that the frequency of NCLSs translocated >5 µm, as well as the mean velocity of NCLS transport in nocodazole-treated cells (Fig. 3B and C; Movie S3), was drastically decreased in cytochalasin D-treated cells (Fig. 3B and C; Movie S4). The characteristics of NCLS transport were similar to those cells transfected with all nucleocapsid components (Fig. 1), and those of nucleocapsid transport in MARV-infected cells (Fig. S2). In summary, NP, VP35, and VP24 nucleocapsid proteins are necessary and sufficient to mediate NCLS transport.
*Live-cell imaging analysis of MARV NCLSs formed by NP, VP35, and VP24. (A) Schematic of the experimental settings for NCLS transport, formed by NP, VP35, and VP24. (B) Huh-7 cells were transfected with the plasmids encoding NP, VP24, VP35GFP, and VP35. At 17 h p.t., different cytoskeleton-modulating drugs were added to the culture medium: 0.15% DMSO, 15 µM nocodazole, or 0.3 µM cytochalasin D for 3 h. Subsequently, cells were subjected to live-cell imaging analysis. The image shows the maximum-intensity projection of time-lapse images of cells, recorded for 3 min; images were captured every 3 s. The small boxed areas are enlarged at the four corners. Scale bars: 10 µm (scale bar in insets, 2 µm). The dotted lines indicate typical NCLS trajectories. (C) The detected signals were analyzed using Imaris software. The length of NCLS trajectories and velocity of NCLS transport were evaluated. The y-axis represents the number of signals. The lines indicate the mean ± SD. Asterisks indicate statistical significance; ***P < 0.001 and ***P < 0.0001.
An exchange of NCLS proteins between MARV and EBOV
The helical structure of NCLS, which is formed by NP, VP35, and VP24, is well-conserved in MARV and EBOV (24, 30), although interactions involving nucleocapsid-forming proteins between MARV and EBOV remain largely unexplored. To analyze the compatibility of nucleocapsid proteins between MARV and EBOV, we exchanged each protein in NP, VP35, VP24, and VP30 to observe the assembly and transport of NCLSs. In the MARV live-cell imaging system, EBOV NP, VP35, and VP24 were incompatible, whereas MARV-VP30 was exchangeable with EBOV-VP30 (Fig. 4A; Movie S5). EBOV-VP30 was also exchangeable with MARV-VP30 in the EBOV live-cell imaging system (Fig. 4B; Movie S6). These results indicate that VP30 proteins are structurally compatible with MARV and EBOV proteins.
Live-cell imaging analysis of each NCLS protein replaced. (A) Huh-7 cells were transfected with plasmids encoding MARV proteins NP, VP35, VP24, and VP30GFP, with one protein replaced with the corresponding EBOV protein (highlighted in bold). The indicated protein-coding plasmids were expressed. (B) Huh-7 cells were transfected with plasmids encoding EBOV proteins NP, VP35, VP24, and VP30GFP, but one protein was replaced with the corresponding MARV protein (highlighted in bold). The indicated protein-coding plasmids were expressed. Live-cell imaging analysis was started from 20 h p.t. The image shows the maximum-intensity projection of time-lapse images of cells, recorded for 3 min; images were captured every 3 s. The small boxed areas are enlarged at the four corners. Scale bars: 10 µm (scale bar in insets, 2 µm). The dotted lines indicate typical NCLS trajectories.
Nucleocapsid protein replacement in minireplicon assays
Little information has been published regarding chimeric systems for evaluating genomic RNA replication among filoviruses (35), and the magnitude and mechanisms of possible compatibility are unclear. Here, we replaced either NP, VP35, or VP30 in optimized minigenome transcription and replication assays (36, 37). In the MARV minigenome assay, the exchange of MARV-NP with EBOV-NP and MARV-VP35 with EBOV-VP35 resulted in the abolishment of reporter activity close to the value of the negative control (in the absence of L protein). On the other hand, the exchange of MARV-VP30 with EBOV-VP30 retained approximately 60% reporter activity, which was higher than that in the absence of MARV-VP30, which demonstrated around 40% reporter activity when the value of MARV-VP30 was set as 100% (Fig. 5A). In EBOV minigenome assays, the replacement of EBOV-VP30 with MARV-VP30 retained approximately 3%–5% reporter activity, which was significantly higher than that in the absence of EBOV-VP30 with less than 1% reporter activity when the value of EBOV-VP30 was set as 100% (Fig. 5B). Interestingly, both MARV-VP30 and EBOV-VP30 were at least partially functional transcriptional activators in the heterologous replicon system.
*Minigenome assay with replacement of one viral protein. (A) HEK293 cells were transfected with MARV minigenome assay components where either NP, VP35, or VP30 was replaced with an EBOV protein. (B) HEK293 cells were transfected with EBOV minigenome assay components in which either NP, VP35, or VP30 was replaced with MARV protein. At 48 h p.t., cells were lysed, and reporter activity was measured. The value of normal MARV (A) or EBOV (B) minigenome component-transfected cells was set to 100%. The negative control (absence of L expression, ΔL) represented the background of the assay. Asterisks indicate statistical significance; *P < 0.05, ***P < 0.001, ***P < 0.0001.
Interactions between NP and VP30
To reveal the molecular mechanisms underlying VP30-mediated transcriptional support activity and VP30-NCLS association in heterologous viruses, we performed immunofluorescence and immunoprecipitation assays. Both MARV-NP and EBOV-NP formed perinuclear inclusions, and both MARV-VP30 and EBOV-VP30 were diffusely distributed in the cytoplasmic regions when they were singly expressed (Fig. 6A and B, left lanes) (15, 38, 39). As previously described, MARV-VP30 accumulated in the MARV-NP-induced inclusions when co-expressed (Fig. 6A, middle lane) (3, 39). Similarly, EBOV-VP30 was localized in the EBOV-NP-induced inclusions when co-expressed (Fig. 6B, middle lane) (11, 15, 38). Notably, EBOV-VP30 also accumulated in MARV-NP-induced inclusions, and MARV-VP30 accumulated in EBOV-NP-induced inclusions (Fig. 6A and B, right lanes).
Homogeneous and heterogeneous NP-VP30 protein interactions. (A, B) Immunofluorescence analysis of Huh-7 cells. (A) Cells were transfected with the following plasmids: MARV-NP or EBOV-VP30 (left lane), MARV-NP and MARV VP30GFP (middle lane), or MARV-NP and EBOV-VP30GFP (right lane). (B) Cells were transfected with a single plasmid: EBOV-NP or MARV-VP30 (left lane), a combination of plasmids EBOV-NP and EBOV-VP30GFP (middle lane), or EBOV-NP and MARV-VP30GFP (right lane). Their intracellular distribution was analyzed using the corresponding NP-specific antibodies and/or autofluorescence. Scale bars: 10 µm. (C, D) Immunoprecipitation assays using MARV-NPflag or EBOV-NPflag in HEK293 cells. (C) MARV-NPflag together with MARV-VP30GFP- or EBOV-VP30GFP-encoding plasmids was transfected into cells. (D) EBOV-NPflag together with EBOV-VP30GFP- or MARV-VP30GFP-encoding plasmids was transfected into cells. At 48 h p.t., the cells were lysed and protein complexes were precipitated using mouse anti-Flag M2 agarose. An aliquot of the cell lysate (input) was collected before precipitation. Elution was achieved using SDS sample buffer. Western blot analysis was performed using Flag-, GFP-, and α-tubulin-specific antibodies. Lane numbers are indicated. Lane 4 shows the absence of NP, representing non-specific interactions involving VP30GFP and agarose beads.
Next, we performed immunoprecipitation assays using MARV-NP^flag^ and EBOV-NP^flag^. EBOV-VP30^GFP^ was precipitated using EBOV-NP^flag^, and MARV-VP30^GFP^ was precipitated using MARV-VP30^GFP^, as expected (Fig. 6C, lane 2, Fig. 6D, lane 2). Moreover, EBOV-VP30^GFP^ was precipitated by MARV-NP^flag^, and MARV-VP30^GFP^ was precipitated by EBOV-NP^flag^ (Fig. 6C and D, lanes 3). These results demonstrate that direct interactions between NP and VP30 bring about colocalization of these proteins in viral inclusions, suggesting that heterologous VP30 is partially functional in minigenome transcription and replication.
PPxPxY motif-mediated NP-VP30 interactions
VP30 binds to the PPxPxY motif in NP, which regulates NP-mediated VP30 dephosphorylation (13, 40), which is a key modulator of transcriptional support activity in both EBOV and MARV (12, 15, 41). To reveal interactions between VP30 and the NP PPxPxY motif, mutations were introduced into this motif (MARV-NP_ΔVP30_ and EBOV-NP_ΔVP30_, Fig. 7A). MARV-NP_ΔVP30_ exhibited reporter activity in the MARV minigenome assay, which was similar to that of the NP wild-type without VP30, regardless of the presence or absence of VP30 (Fig. 7B). In contrast, EBOV-NP_ΔVP30_ showed a decrease in reporter activity to the same level as that of the negative control when VP30 was not expressed; however, reporter activity was rescued by VP30 expression (Fig. 7C). As reported previously (11), VP30 in EBOV supports transcription and replication activity even when it loses its interaction with NP. Notably, VP30 does not exhibit transcription and replication support activity in MARV when it loses its interaction with NP.
*PPxPxY motif-mediated interactions between NP and VP30 proteins. (A) Schematic representation of the regional orientation of the filoviral nucleoprotein orthologs. The N-terminal domain, intrinsically disordered region (IDR), and C-terminal domain are illustrated. Amino acid sequences of PPxPxY surrounding regions were selected, and this motif is presented as bold and highlighted. The numbers indicate the number of amino acids counted from the N-terminus. PPxPxY motif mutants are enclosed in dotted boxes, named MARV-NPΔVP30 and EBOV-NPΔVP30. LLOV: Lloviu cuevavirus, MLAV: Mengla dianlovirus. (B, C) Minigenome assay in HEK293 cells. (B) In MARV minigenome assays, cells were transfected with the minigenome components MARV-NP or MARV-NPΔVP30, together with MARV-VP30 or EBOV-VP30. (C) In EBOV minigenome assays, cells were transfected with the minigenome components EBOV-NP or EBOV-NPΔVP30, together with EBOV-VP30 or MARV-VP30. At 48 h p.t., cells were lysed, and reporter activity was measured. The value of MARV-NP- or EBOV-NP-expressing cells was set to 100%. The mean and SD of three independent experiments are shown. Asterisks indicate statistical significance; **, P < 0.01; ***, P < 0.001; ***, P < 0.0001. (D, E) Immunofluorescence assay in Huh-7 cells. Cells were transfected with the indicated protein-encoding plasmids ([D] MARV and [E] EBOV). The intracellular distribution of proteins noted above the images was visualized using NP-specific antibodies and autofluorescence, and merged images were visualized. The small boxed areas are enlarged at the four corners. Scale bars: 10 µm (scale bar in insets, 2 µm). (F, G) Immunoprecipitation assay in HEK293 cells. Cells were transfected with the indicated protein-encoding plasmids ([F] MARV and [G] EBOV). NP-encoding plasmids were fused with a FLAG tag, cells were lysed, and protein complexes were precipitated using mouse anti-FLAG M2 agarose at 48 h p.t. An aliquot of cell lysate (input) was collected before precipitation. Elution was achieved using SDS sample buffer. Western blotting was performed using FLAG-, GFP-, and alpha-tubulin-specific antibodies.
Next, we employed confocal microscopy to visualize the formation of inclusion bodies and the localization of NP and VP30. This analysis demonstrated that MARV-NP_ΔVP30_ could form inclusion bodies but failed to incorporate either MARV-VP30 or EBOV-VP30 (Fig. 7D). Similarly, EBOV-NP_ΔVP30_ did not incorporate EBOV-VP30 or MARV-VP30 into the inclusions (Fig. 7E). The recruitment of L and VP35 encapsulated bodies was not affected by the PPxPxY motif, although L of MARV could not be evaluated (Fig. S3).
To clarify the interaction between NP and VP30 mediated by the PPxPxY motif, we conducted immunoprecipitation experiments. MARV-NP_ΔVP30_ lost its ability to interact with MARV-VP30 (Fig. 7F, lane 4), and EBOV-NP_ΔVP30_ was unable to precipitate EBOV-VP30 (Fig. 7G, lane 4).
These results indicate that the interactions between NP and VP30 are regulated by the conserved PPxPxY motif in NP, which is well conserved among filoviruses without affecting the formation of inclusions.
PPxPxY motif and NCLS assembly, and VLP formation
Microscopic analyses were performed to investigate the role of the PPxPxY motif in NCLS assembly and transport. First, to analyze whether the NP mutants could form NCLSs, transmission electron microscopy was conducted. Tubular-like structures with electron-dense walls, representing condensed nucleocapsids, were detected in the presence of MARV-NP_ΔVP30_ (Fig. 8A, arrowheads) and EBOV-NP_ΔVP30_ (Fig. 8B, arrowheads).
PPxPxY motif and VP30 proteins association to NCLSs, VLPs. (A, B) Huh-7 cells expressing the indicated protein-coding plasmids were fixed at 48 h p.t. and processed for transmission electron microscopy analyses. The absence or presence of electron-dense walls in the tubular-like NCLSs is indicated by arrowheads on the transversal or longitudinal sections of NCLSs. The boxed areas in the left images are enlarged and are shown on the right. Scale bars are as follows: upper images, 1 µm; lower images, 100 nm. (C, D) Live-cell imaging in Huh-7 cells. Cells were transfected with the indicated protein-encoding plasmids together with MARV-VP35- and MARV-VP24-encoding plasmids (C). Cells were transfected with the indicated protein-encoding plasmids together with EBOV-VP35- and EBOV-VP24-encoding plasmids (D). Live-cell imaging analysis was started from 20 h p.t. The image shows the maximum-intensity projection of time-lapse images of cells, recorded for 2–3 min; images were captured every 2–3 s. The small boxed areas are enlarged at the four corners. Scale bars: 10 µm (scale bar in insets, 2 µm). The dotted lines indicate typical NCLS trajectories. (E) Schematic representation of MARV and EBOV VLP assay. (F, G) HEK293 cells were transfected with plasmids expressing all viral structural proteins and a MARV- or EBOV-specific minigenome encoding Renilla luciferase, together with a plasmid encoding firefly luciferase for normalization. The released VLPs were collected from supernatant, which was purified via sucrose cushion at 72 h p.t. Western blotting was performed to determine viral protein expression in producer cells and VLP fractions ([F] MARV, [G] EBOV).
Next, we conducted live-cell imaging analyses of NCLS containing homologous or heterologous VP30-GFP in conjunction with NP or NP_ΔVP30_. As shown in Fig. 4, long sequential lines indicating transport-competent NCLSs were observed in MARV-NP- and EBOV-NP-containing NCLSs (Fig. 8C). In contrast, MARV-NP_ΔVP30_- and EBOV-NP_ΔVP30_-containing NCLSs demonstrated only diffusely distributed signals from both MARV and EBOV live-cell imaging systems (Fig. 8D).
Subsequently, we examined the process of VLP formation following NCLS transport. In the VLP assay, we transfected cells with NP, VP24, VP35, VP30, L, VP40, GP, a minigenome, and T7, and subsequently collected the VLPs secreted into the cell supernatant (Fig. 8E). Purified VLPs were assessed by western blotting. In both the MARV and EBOV VLP assays, we observed the formation of VP40-containing VLPs, even in the absence of NP, whereas VP30 was undetectable (Fig. 8F and G, lanes 1). However, in the presence of wild-type NP, the incorporation of VP30 was confirmed; this was not observed in the NP mutants (MARV-NP_ΔVP30_ and EBOV-NP_ΔVP30_) (Fig. 8F and G, lanes 2 and 3).
In summary, mutations in the PPxPxY motif are crucial for VP30 association with NCLSs and for VP30 incorporation into VLPs in both MARV and EBOV.
PPxPxY motif and NC protein interactions
The association between NP and VP35 was reported upon NP binding peptide (20–48 a.a. of VP35) to the N-terminal side of NP (residues 25–457), and the basic patch of VP35 (residues 220–251) to the central domain of NP (22, 40, 42–44). Regarding the NP-VP24 interaction, two molecules of VP24 bind to two molecules of NP in distinct configurations, and mutations in NP_R132A_ and NP_H196A_ inhibit the formation of NCLS (31). Given that the structure of the C-terminal region of NP and interactions among NCLS proteins are largely unknown, we used AlphaFold2 and AlphaFold3 (45, 46) to predict the binding of PPxPxY peptides to VP35 and VP24. Confident structural predictions were obtained only for MARV-VP35, EBOV-VP35, and EBOV-VP24 (pLDDT >80, Fig. S4A through C). All the predicted structures indicated that both MARV and EBOV VP35 interacted with the PPxPxY peptide in a similar structural region. These results suggest that the PPxPxY peptide may interact with the positively charged VP35 surface in both MARV and EBOV (Fig. S4D and E). However, a protein complex was not predicted for the peptide with an alanine substitution. Additionally, we were unable to predict a highly confident binding of the PPxPxY peptide to MARV-VP24; in contrast, both PPxPxY and the alanine substitution mutant peptides appeared to interact with EBOV-VP24 (Fig. S4C).
To reveal the NP-PPxPxY motif and VP35 interaction, microscopy and immunoprecipitation assays were performed. Immunofluorescence microscopy analyses revealed that MARV-NP_ΔVP30_ recruited MARV-VP35 to these inclusions (Fig. S5A). Similarly, EBOV-NP_ΔVP30_ recruited EBOV-VP35 (Fig. S5B). Next, we conducted live-cell imaging analyses of NCLSs containing either NP or NP_ΔVP30_ along with their respective VP35-GFP. Both MARV-NP_ΔVP30_ and EBOV-NP_ΔVP30_ exhibited transport-competent NCLSs (Fig. S5C and D). Finally, we performed immunoprecipitation assays, demonstrating that both MARV-NP_ΔVP30_ and EBOV-NP_ΔVP30_ successfully precipitated their respective VP35 (Fig. S5E and F, lanes 4). These results suggest that the PPxPxY motif does not govern the interactions among NCLS-forming proteins; rather, it regulates the interaction between NP and VP30 in a localized manner.
In summary, VP30 exhibits compatibility between MARV and EBOV, with transcription and replication activities partially sustained by heterologous VP30 (Fig. 9A). The binding of NP to VP30 is regulated by the PPxPxY motif; when a mutation is introduced, VP30 is unable to bind to NP. Consequently, there was no association between VP30 and NCLS (Fig. 9B). Conversely, when the motif is intact, heterologous VP30 can bind to NCLS, facilitating intracellular transport (Fig. 9C).
Role of the PPxPxY motif of NP during NCLS assembly in MARV and EBOV. This figure illustrates the current understanding of NCLS assembly and VP30-NCLS associations across three key processes: (1) viral genome transcription/replication, (2) nucleocapsid assembly, and (3) nucleocapsid transport. (1) In filoviruses, NP forms perinuclear inclusions, where viral genome transcription/replication and nucleocapsid synthesis occur. VP30 co-localizes with heterologous NP-forming inclusions and partially restores heterologous minigenome transcription/replication. (2) Mutations in the PPxPxY motif regulate NP-VP30 interactions in both MARV and EBOV. (3) VP30 is associated with heterologous NP, facilitating its linkage to NCLS. This indicates that NCLS is initially formed by NP, VP35, and VP24, with VP30 association occurring during or after NCLS assembly.
DISCUSSION
In the present study, we revealed that the core structure of nucleocapsids, NCLSs, is formed from NP, VP35, and VP24 (Fig. 1 to 3) in agreement with EBOV, using live-cell imaging systems based on the ectopic expression of fluorescently labeled viral proteins in MARV. Based on these observations, we characterized the nucleocapsid compatibility between MARV and EBOV and sought to reveal the molecular interface between the nucleocapsid-forming proteins.
The characteristic filoviral NP-RNA helical complex provides a scaffold for nucleocapsid formation, which is responsible for the transcription and replication of viral RNAs (4, 7, 8, 24). Previously, MARV-VP30 was reported to be partially functional in the transcription and replication of EBOV chloramphenicol acetyltransferase reporter assays (35). Our results demonstrated that EBOV-VP30 also supported transcription and replication of MARV minigenome (Fig. 5B), indicating that filoviruses have conserved machinery for VP30-nucleocapsid associations. On the other hand, VP30 plays different roles in recombinant viruses production; MARV was rescued by EBOV VP30 instead of MARV VP30 expressions, whereas EBOV was rescued only by EBOV VP30 (47). VP30 is a phosphoprotein, and its dephosphorylation is crucial for transcriptional support in EBOV rather than in MARV (11, 13, 41). The LxxIxE and PPxPxY motifs located adjacent to the C-terminus of NP are conserved in filoviruses (13). In EBOV, VP30 binds to NP at the PPxPxY motif, and VP30 is dephosphorylated by PP2A, which is recruited by the LxxIxE motif (13). Notably, mutations involving the PPxPxY motif did not significantly affect the transcriptional support activity in either the MARV or EBOV minigenome assays (Fig. 7B and C). Defective interactions involving NP-VP30 proteins do not cause defective viral genome transcription/replication in EBOV (11, 40), indicating that a high-affinity interaction between NP and VP30 is not strictly required for viral RNA synthesis, and minimal binding of these proteins mediates viral RNA synthesis in EBOV (48). In contrast, the transcription and replication activity of MARV slightly decreased due to mutation of the PPxPxY motif. However, this reduction was consistent with the levels observed in the absence of VP30 in wild-type NP, suggesting that this phenomenon reflects the influence of VP30 rather than the activity of the motif itself in transcription and replication. Multiple studies have demonstrated significant differences in the effects of VP30 on transcription and replication in the minigenome systems of MARV and EBOV, primarily attributed to VP30’s influence on the polymerase complex (11, 41). In EBOV, the absence of VP30 reduces activity to less than 1% of wild-type levels (12, 14), whereas in MARV, even without VP30, 60%–70% of transcription and replication activity is maintained (41). This difference may be due to variations in the sensitivity between the two minigenome assays. Specifically, the original reporter activity values for EBOV were reported to be 10 times higher than those for MARV (Fig. S6). This difference in sensitivity may partly explain why the effects of VP30 are more clearly observed; however, further investigation is warranted for another possibility.
In filoviruses, nucleocapsids form a left-handed helix with an inner nucleoprotein layer decorated with protruding arms composed of VP24 and VP35 (23, 24, 30). Even in a recent model, the intrinsically disordered C-terminal region of NP (aa 450–600), which is critical for nucleocapsid formation in the presence of VP24 and VP35, has been unclear due to its flexibility and insufficient resolution of the electron microscopy (EM) map (31, 32). This study reveals a common aspect of filoviral nucleocapsid assembly, that is, heterologous VP30 associates with NP and supports transcription/replication in the inclusions in both MARV and EBOV ([Fig. 5 and 6](#F5 F6)). Intriguingly, the PPxPxY motif regulates the interaction with NP-VP30 but does not affect the assembly and transport of NCLSs (Fig. 8A and B; Fig. S4C and D). Consequently, introducing mutations in this motif inhibits the association of VP30 to VLPs (Fig. 8F and G). Noteworthy, it has been reported that transcription and replication are inhibited by the addition of competitive peptides against this motif in EBOV minigenome (48). Moreover, PPxPxY motif-bearing proteins, such as RBBP6, hnRNPUL1, and PEG10, modulate EBOV transcription and replication through discrete mechanisms (49), highlighting the importance of this motif in EBOV replication. Given that research on this motif in MARV has not yet been reported, this remains a topic for future investigation.
Using AlphaFold2/3 prediction (45, 46), the PPxPxY peptide may bind to the positively charged surface of VP35, both in MARV and EBOV, although no such protein complexes are predicted when peptides hold alanine substitutions (Fig. S3). In EBOV, amino acid residues R225, H240, K248, and K251 of VP35 are reported to be important for its NP interaction (50), which forms the first basic patch (amino acid residues 222–251) (22). Interestingly, residues Q241, Q244, and K248 located in this basic patch have been shown to be part of the PPxPxY-binding motif of NP, but mutations in this motif did not affect the NP-VP35 interaction or the formation of NCLS (Fig. 8A and B; Fig. S4). AlphaFold predictions are computational and speculative; therefore, biophysical interaction assays to validate predicted interactions between the PPxPxY motif and NC proteins are anticipated in the future.
Pseudoviruses are useful for studying the entry process (51); however, they do not recapitulate the complexities of nucleocapsid assembly, protein interactions, and intracellular transport. Therefore, VLPs and minigenome systems were employed in this study. However, these methods have some inherent limitations. Notably, the use of cultured cell lines for microscopic analysis may not fully reflect an in vivo environment. Since antigen-presenting cells such as monocytes, macrophages, dendritic cells, and endothelial cells play prominent roles in filovirus infection (52), analyzing the interactions of NCLS proteins and transport mechanisms in these cell types would provide more clinically relevant insights. While live-cell imaging of NCLS can help mitigate biosafety concerns, recombinant viruses such as conditional recombinant virus systems (53) offer a promising approach to study these processes under more physiological conditions. In particular, future investigations employing VP30-deficient recombinant filoviruses (e.g., EBOV-ΔVP30 or MARV-ΔVP30) are anticipated to clarify the role of the PPxPxY motif of VP30 in NC formation and viral transcription, bridging the gap between reductionist systems and natural infection.
In conclusion, our study demonstrated that the PPxPxY motif not only regulates the binding between NP and VP30 but also influences the association of VP30 with NCLS. Interestingly, the interaction between NP and VP30 through the PPxPxY motif is regulated in a somewhat permissive manner, exhibiting compatibility between MARV and EBOV. This suggests a potential for developing drugs that inhibit the replication of a wide range of filoviruses by targeting non-specific binding through this motif.
MATERIALS AND METHODS
Cells and viruses
Huh-7 and HEK293 cells were maintained at 37°C and 5% CO_2_ in Dulbecco’s modified Eagle’s medium (DMEM; Life Technologies) supplemented with 10% (vol/vol) fetal bovine serum (FBS; PAN Biotech), 5 mM L-glutamine (Q; Life Technologies), 50 U/mL penicillin, and 50 µg/mL streptomycin (PS; Life Technologies). MARV (Musoke accession no. DQ217792.1, GenBank) and recombinant MARV were propagated on VeroE6 cells as described previously (54). All work with infectious viruses was performed in a bio-safety level (BSL)-4 facility at Philipps-Universität Marburg following national legislation and guidelines.
Plasmids and transfection
Plasmids encoding the MARV proteins pCAGGS-MARV-NP, pCAGGS-MARV-VP35, pCAGGS-MARV-VP30, pCAGGS-MARV-VP24, pCAGGS-MARV-L, pCAGGS-MARV-VP40, and pCAGGS-MARV-GP, as well as the T7-driven MARV minigenome encoding Renilla luciferase and pCAGGS-T7 polymerase, were used as previously described (36, 37). Plasmids encoding MARV-VP30^GFP^ and MARV-VP35^GFP^ fusion proteins have been described previously (2, 3).
Plasmids encoding EBOV proteins (pCAGGS-EBOV-NP, pCAGGS-EBOV-VP35, pCAGGS-EBOV-VP30, pCAGGS-EBOV-VP24, and pCAGGS-EBOV-L) and the T7-driven EBOV minigenome encoding Renilla luciferase were prepared as described previously (11, 55).
The cloning procedure for plasmids with mutations introduced at the PxPPxY motif with Flag-tagged plasmids (pCAGGS-MARV-NP^flag^, pCAGGS -MARV-NP_ΔVP30_^flag^, pCAGGS-EBOV-NP^flag^, and pCAGGS-EBOV-NP_ΔVP30_^flag^) has been described previously (26).
Transfection was performed in Opti-MEM without phenol red (Life Technologies) using TranSIT (Mirus), according to the manufacturer’s instructions.
SDS-PAGE and western blot analysis
SDS-PAGE and western blot analyses were performed as previously described (11, 56). Protein detection was performed using Image Lab software (Bio-Rad) or Image Reader LAS-3000 (Fujifilm) for horseradish peroxidase (HRP)-conjugated secondary antibodies, as indicated in the antibodies section below.
Immunofluorescence analysis and confocal laser scanning microscopy
Immunofluorescence analyses were performed as previously described (57, 58). Microscopic images were acquired using a Leica SP5 confocal laser scanning microscope with a 63× oil objective, an Olympus FV3000 microscope with a 100× oil objective, or a Keyence BZ-X810 microscope with a 100× oil objective (Olympus). Cells were grown on µ-Slide 8 or 12 wells (ibidi) and fixed with 4% paraformaldehyde 20 h post-transfection. Nuclear staining was performed using Hoechst 33342 (Dojindo).
Antibodies
The following primary antibodies were used for immunofluorescence: mouse anti-MARV-NP (37), rabbit anti-MARV-NP (IBT), rabbit anti-MARV-VP30 (37), guinea pig anti-MARV-VP35 (37), rabbit anti-MARV-VP24 (37), chicken anti-EBOV-NP (12), rabbit anti-EBOV-NP (IBT), and rabbit anti-EBOV-VP30 (11). The corresponding secondary antibodies were donkey anti-mouse Alexa488 (Abcam), donkey anti-mouse Alexa594 (Abcam), donkey anti-mouse-IRDye 680RD (LI-COR), donkey anti-mouse Alexa488 (Abcam), donkey anti-rabbit Alexa594 (Abcam), donkey anti-rabbit IRDye 680RD (LI-COR), goat anti-guinea pig Alexa594 (Abcam), goat anti-chicken Alexa594 (Thermo Fisher Scientific), and donkey anti-chicken IRDye 680RD (LI-COR).
The following primary antibodies were used for the western blot analysis: mouse anti-MARV-NP monoclonal antibody (33), rabbit anti-MARV-NP antibody (IBT), mouse anti-MARV-VP40 monoclonal antibody (33), chicken anti-EBOV-NP polyclonal antibody (see above), rabbit anti-EBOV antibody (IBT), rabbit anti-EBOV-VP40 antibody (IBT), rabbit anti-GFP antibody (Rockland), mouse anti-FLAG M2 monoclonal antibody (Sigma-Aldrich), rabbit anti-HA monoclonal antibody (ROCKLAND), and rabbit anti-α-tubulin antibody (MBL). The corresponding secondary antibodies used were HRP-conjugated goat anti-mouse IgG (Abcam), HRP-conjugated goat anti-rabbit IgG (Abcam), and HRP-conjugated goat anti-chicken IgY (Abcam).
Minigenome reporter assay
MARV minigenome assays were performed as previously described (37). Briefly, plasmids for the minigenome assay (500 ng of pCAGGS-NP, 100 ng of pCAGGS-VP35, 100 ng of pCAGGS-VP30, and 1,000 ng of pCAGGS-L, 500 ng of a MARV-specific minigenome encoding the Renilla luciferase reporter gene, and 500 ng of pCAGGS-T7 polymerase, and 50 ng of pGL-encoding firefly luciferase reporter gene for normalization) were transfected into HEK293 cells. EBOV minigenome assays were performed as previously described (59). Briefly, plasmids for minigenome assays (125 ng of pCAGGS-NP, 100 ng of pCAGGS-VP35, 100 ng of pCAGGS-VP30, and 1,000 ng of pCAGGS-L, with 250 ng of EBOV-specific minigenome encoding the Renilla luciferase reporter gene, 250 ng of pCAGGS-T7 polymerase, and 50 ng of pGL-encoding firefly luciferase reporter gene for normalization) were transfected into HEK293 cells. Cells were lysed and subjected to a luciferase reporter assay (PJK).
Live-cell imaging microscopy
A total of 8 × 10^4^, 4 × 10^4^, or 2 × 10^4^ Huh-7 cells were seeded onto a µ-Dish 35 mm, a µ-Slide 4 well, or a µ-Slide 8 well (ibidi) and incubated in DMEM/PS/Q with 10% FBS. To observe MARV nucleocapsid transport, cells were infected with a multiplicity of infection of 1. The inoculum was replaced with fresh medium at 1 h post-infection (33). Subsequently, 500 ng of DNA encoding the green fluorescent fusion protein was transfected. To observe MARV NCLS transport, each well was transfected with the following plasmids encoding MARV proteins: (500 ng of pCAGGS-NP, 100 ng of pCAGGS-VP35, 100 ng of pCAGGS-VP30^GFP^, and 100 ng of pCAGGS-VP24), together with the MARV minigenome-expressing plasmid and T7 polymerase-coding plasmid (36, 37). To observe EBOV NCLS transport, each well was transfected with the following plasmids encoding EBOV proteins: 500 ng of pCAGGS-NP, 200 ng of pCAGGS-VP35, 200 ng of pCAGGS-VP24, and 200 ng of pCAGGS-VP30-GFP. The inoculum was removed at 1 h p.t., and CO_2_-independent Leibovitz’s medium (Life Technologies) with PS/Q, non-essential amino acid solution, and 3%–20% (vol/vol) FBS was added. Live-cell time-lapse experiments were recorded with a Nikon ECLIPSE TE2000-E using a 63× oil objective, GE Healthcare Delta Vision Elite using a 60× oil objective, Keyence BZ-X810 microscope using a 100× oil objective under BSL-2, and Leica DMI6000B under BSL-4 using a 63× oil objective equipped with a remote-control device to operate the microscope outside the BSL-4 facility (3). To visualize representative trajectories, NCs were detected using a size threshold of 1 µm, and NCLSs using a threshold of 0.5 µm. Signals with a length of at least 5 µm that exhibited directional movement were then traced. While many inclusion bodies are larger than NCs and NCLSs, it is difficult to exclude smaller, nucleus-distant inclusion bodies solely based on size. However, within short observation periods, inclusion bodies typically display random, non-directional movements and can be distinguished from the movements of NCs and NCLSs.
Treatment of cells with cytoskeleton-modulating drugs
Cells were treated with 15 µM nocodazole (Sigma), 0.3 µM cytochalasin D (Sigma), or 0.15% DMSO (Sigma), as previously described (3). The chemicals were added to the cell culture medium 3 h prior to observation.
Image processing and analysis
The acquired pictures and movie sequences were processed using the Imaris tracking module (Bitplane; Oxford Instruments, Abingdon, UK) (60). The size of spots >1 µm for nucleocapsids and 0.5 µm for NCLSs was collected. Subsequently, “Quality”-based optimization of the detected spots was performed. The tracking algorithm of “Autoregressive Motion” was applied for tracking with a “Maximum Distance” of 1 and “Maximum Gap” of 1 with filling of gaps with detected objects. Detected trajectories of length >5 µm, duration >15 s, and track straightness >0.2 were processed for the analyses. Moving signals were collected from approximately 10 cells derived from three independent experiments.
Co-immunoprecipitation analysis
Co-immunoprecipitation assays were performed as previously described (26). For SDS-PAGE, elution was achieved with 70 µL of SDS sample buffer (Fujifilm) and subjected to gel electrophoresis and western blot analysis.
Ultrathin section electron microscopy
Huh-7 cells were seeded on a 12-well plate and transfected with 1 µg NP or NP mutant-encoding plasmids, 0.5 µg VP24, and 0.5 µg VP35-encoding plasmids. At 48 h post-transfection, cells were fixed with aldehydes, post-fixed with 1% osmium tetroxide, dehydrated in a graded ethanol series, and embedded in EPON 812 (TAAB, Berks, UK). Ultrathin sections were stained with uranyl acetate and lead citrate and observed under a Hitachi HT-7700 microscope operated at 80 kV (Hitachi Hi-Tech, Tokyo, Japan) (31).
VLP assay
VLP assays were conducted following established protocols (37, 55, 61), with minor modifications. HEK293 cells were seeded in six-well plates and transfected with plasmids encoding all structural proteins and reporter genes for either MARV (500 ng of pCAGGS-NP, 100 ng of VP35, 500 ng of VP40, 500 ng of GP, 100 ng of VP30, 100 ng of VP24, 1,000 ng of L, 500 ng of pANDY-3M5M, 500 ng of pCAGGS-T7 polymerase) or EBOV (125 ng of pCAGGS-NP, 100 ng of VP35, 250 ng of VP40, 250 ng of GP, 100 ng of VP30, 100 ng of VP24, 1,000 ng of L, 250 ng of pANDY-3E5E, 250 ng of pCAGGS-T7 polymerase). Plasmids encoding the firefly luciferase reporter gene were used for normalization. After 72 h post-transfection, culture supernatants were collected, and VLPs were purified by ultracentrifugation using a 20% sucrose cushion. VLPs were analyzed for VP30 incorporation using proteinase K digestion assay (37).
AlphaFold-multimer prediction
To predict the complex structure of the NP-PPxPxY peptide and VP35 or VP24 in MARV or EBOV, we employed AlphaFold structural predictions. The amino acid sequences of EBOV and MARV proteins were obtained from GenBank (ID: EBOV: NC_002549, MARV: DQ217792.2). The structural complex of EBOV and MARV NPs with each peptide sequence was predicted in AlphaFold2 and AlphaFold3 software (45, 46). The most confidently predicted structures (high pLDDT values) were visualized and assessed using UCSF Chimera software (62).
Statistical analysis
Data represent the mean values and standard deviations from at least three independent experiments. Statistical analyses were performed using GraphPad Prism software (version 8.0). Normally distributed samples were analyzed using Student’s t-test. Statistically significant differences are indicated by asterisks (*, P < 0.05; **, P < 0.01; ***, P < 0.001; ****, P < 0.0001).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akira S, Uematsu S, Takeuchi O. 2006. Pathogen recognition and innate immunity. Cell 124:783–801. doi:10.1016/j.cell.2006.02.01516497588 · doi ↗ · pubmed ↗
- 2Takamatsu Y, Kolesnikova L, Becker S. 2018. Ebola virus proteins NP, VP 35, and VP 24 are essential and sufficient to mediate nucleocapsid transport. Proc Natl Acad Sci USA 115:1075–1080. doi:10.1073/pnas.171226311529339477 PMC 5798334 · doi ↗ · pubmed ↗
- 3Schudt G, Kolesnikova L, Dolnik O, Sodeik B, Becker S. 2013. Live-cell imaging of Marburg virus-infected cells uncovers actin-dependent transport of nucleocapsids over long distances. Proc Natl Acad Sci USA 110:14402–14407. doi:10.1073/pnas.130768111023940347 PMC 3761630 · doi ↗ · pubmed ↗
- 4Sugita Y, Matsunami H, Kawaoka Y, Noda T, Wolf M. 2018. Cryo-EM structure of the Ebola virus nucleoprotein–RNA complex at 3.6 Å resolution. Nature 563:137–140. doi:10.1038/s 41586-018-0630-030333622 · doi ↗ · pubmed ↗
- 5Desfosses A, Milles S, Jensen MR, Guseva S, Colletier JP, Maurin D, Schoehn G, Gutsche I, Ruigrok RWH, Blackledge M. 2019. Assembly and cryo-EM structures of RNA-specific measles virus nucleocapsids provide mechanistic insight into paramyxoviral replication. Proc Natl Acad Sci USA 116:4256–4264. doi:10.1073/pnas.181641711630787192 PMC 6410849 · doi ↗ · pubmed ↗
- 6Ker DS, Jenkins HT, Greive SJ, Antson AA. 2021. Cryo EM structure of the Nipah virus nucleocapsid assembly. P Lo S Pathog 17:e 1009740. doi:10.1371/journal.ppat.100974034270629 PMC 8318291 · doi ↗ · pubmed ↗
- 7Fujita-Fujiharu Y, Sugita Y, Takamatsu Y, Houri K, Igarashi M, Muramoto Y, Nakano M, Tsunoda Y, Taniguchi I, Becker S, Noda T. 2022. Structural insight into Marburg virus nucleoprotein-RNA complex formation. Nat Commun 13:1191. doi:10.1038/s 41467-022-28802-x 35246537 PMC 8897395 · doi ↗ · pubmed ↗
- 8Hu S, Fujita-Fujiharu Y, Sugita Y, Wendt L, Muramoto Y, Nakano M, Hoenen T, Noda T. 2023. Cryoelectron microscopic structure of the nucleoprotein–RNA complex of the European filovirus, Lloviu virus. PNAS Nexus 2:pgad 120. doi:10.1093/pnasnexus/pgad 12037124400 PMC 10139700 · doi ↗ · pubmed ↗