Stoichiometry of HIV-1 envelope glycoprotein protomers with changes that stabilize or destabilize the pretriggered conformation
Zhiqing Zhang, Saumya Anang, Qian Wang, Hanh T. Nguyen, Hung-Ching Chen, Ta-Jung Chiu, Derek Yang, Amos B. Smith, Joseph G. Sodroski

TL;DR
This study explores how changes in HIV's envelope protein affect its pretriggered conformation, showing how many protein units need to be modified to stabilize or destabilize this state.
Contribution
The study reveals the stoichiometric requirements for stabilizing or destabilizing the HIV-1 Env trimer's pretriggered conformation.
Findings
Strongly stabilizing changes in one Env protomer can stabilize the entire trimer's pretriggered conformation.
Weakly stabilizing changes require all three protomers to be modified for effective PTC stabilization.
Destabilizing changes in a single protomer can disrupt the pretriggered conformation of the trimer.
Abstract
During human immunodeficiency virus (HIV-1) entry into host cells, binding to the receptors, CD4 and CCR5/CXCR4, triggers conformational changes in the metastable envelope glycoprotein (Env) trimer ((gp120-gp41)3). CD4 binding induces Env to make transitions from its pretriggered conformation (PTC) to more “open” conformations that are sensitive to inhibition by antibodies, CD4-mimetic compounds (CD4mcs), and exposure to cold. Changes in functional membrane Envs that either stabilize or destabilize the PTC have been identified. Here, we investigate the stoichiometric requirements for the PTC-stabilizing and -destabilizing changes in the Env protomers. To this end, we generated viruses bearing Envs with mixed protomers exhibiting different degrees of PTC stability and determined the sensitivity of the viruses to cold (0°C) and, in some cases, to a CD4mc. The number of stabilized Env…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
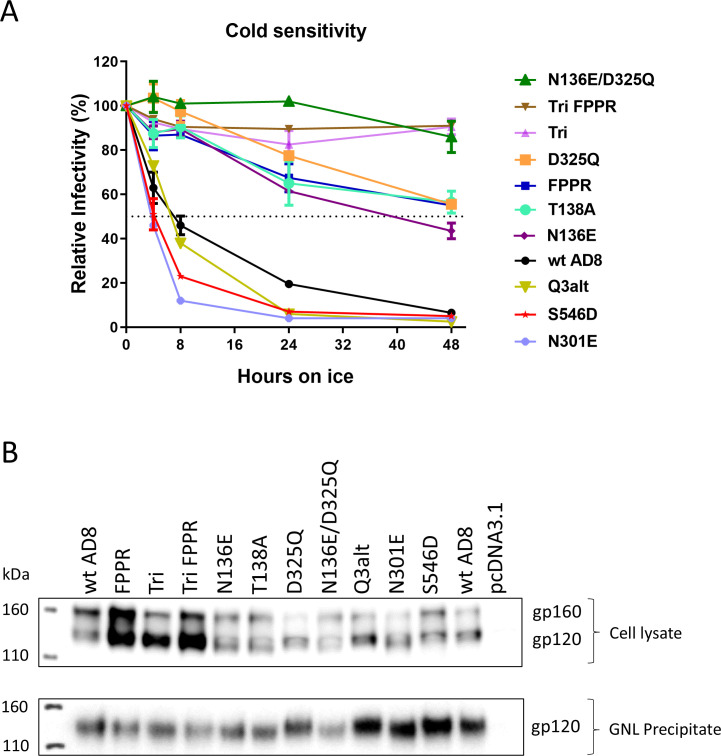 Fig 1
Fig 1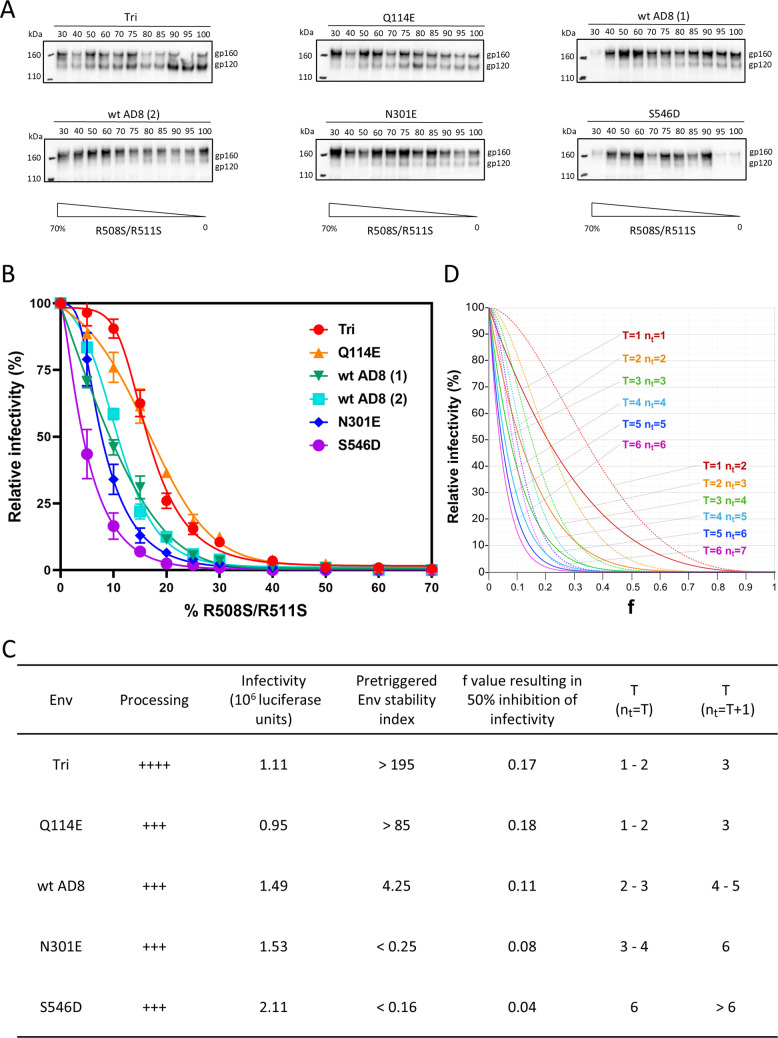 Fig 2
Fig 2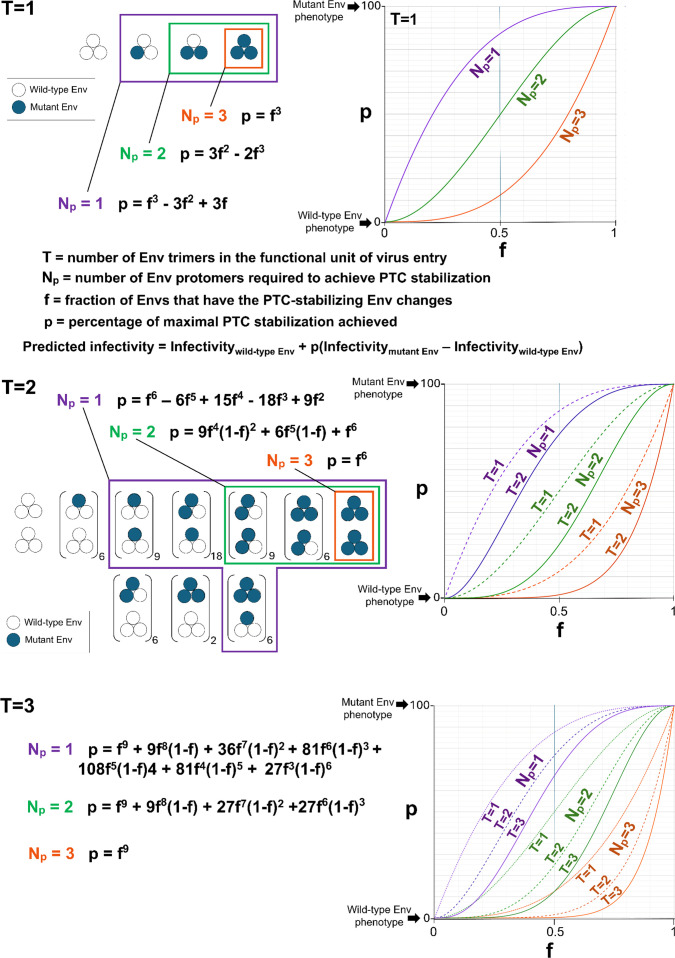 Fig 3
Fig 3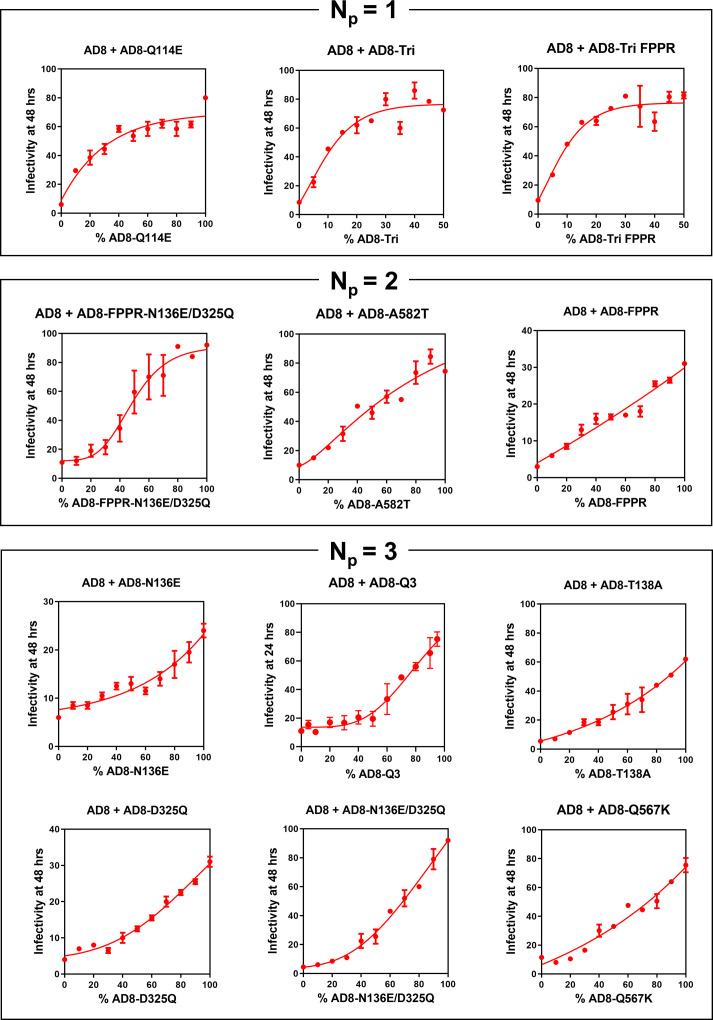 Fig 4
Fig 4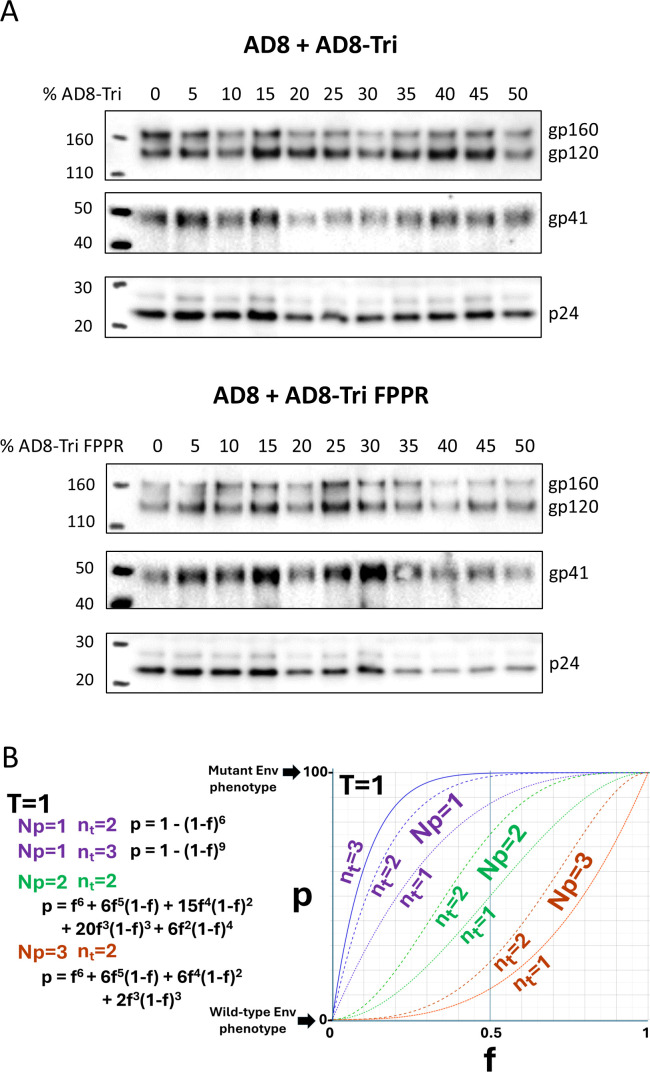 Fig 5
Fig 5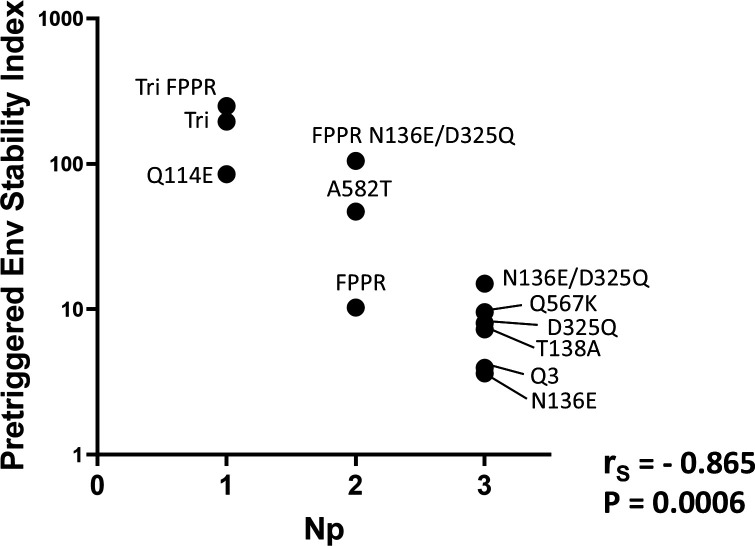 Fig 6
Fig 6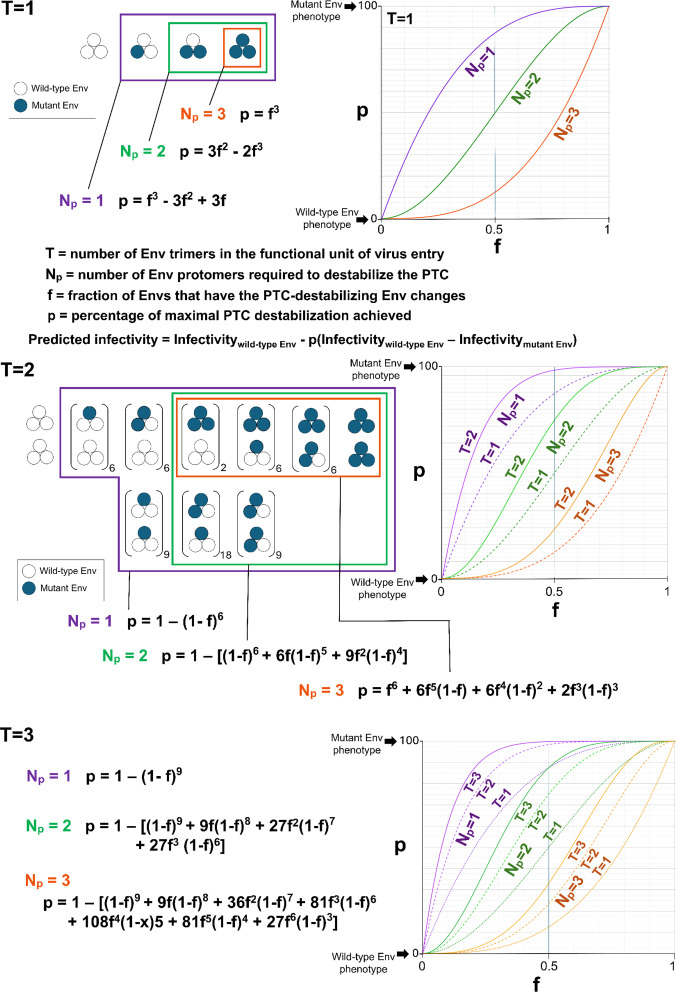 Fig 7
Fig 7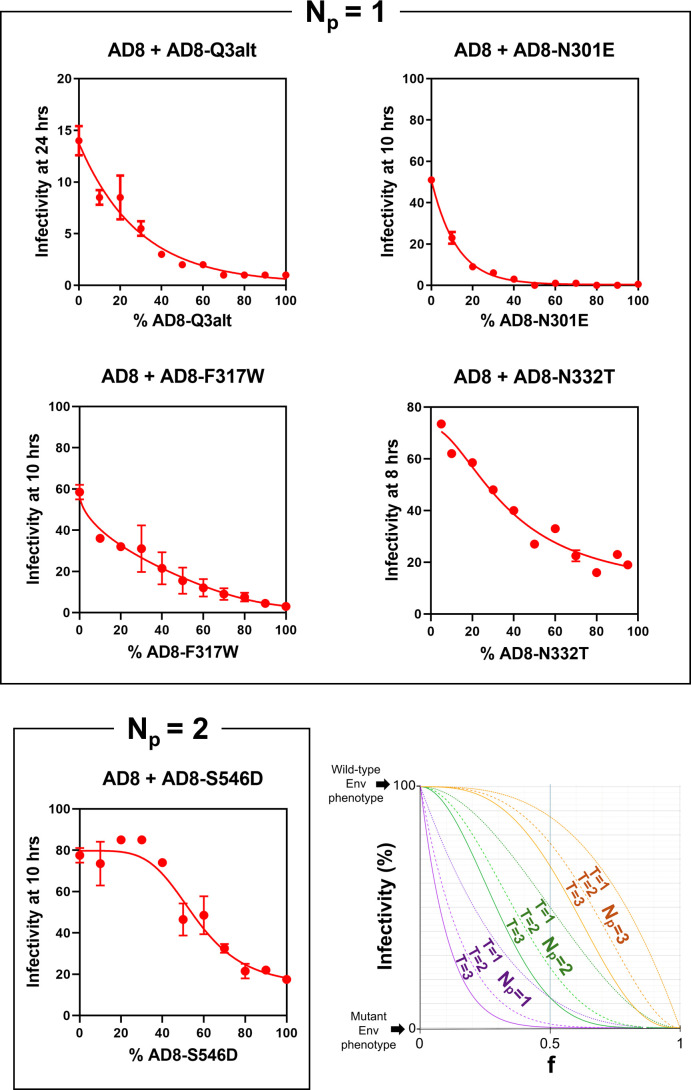 Fig 8
Fig 8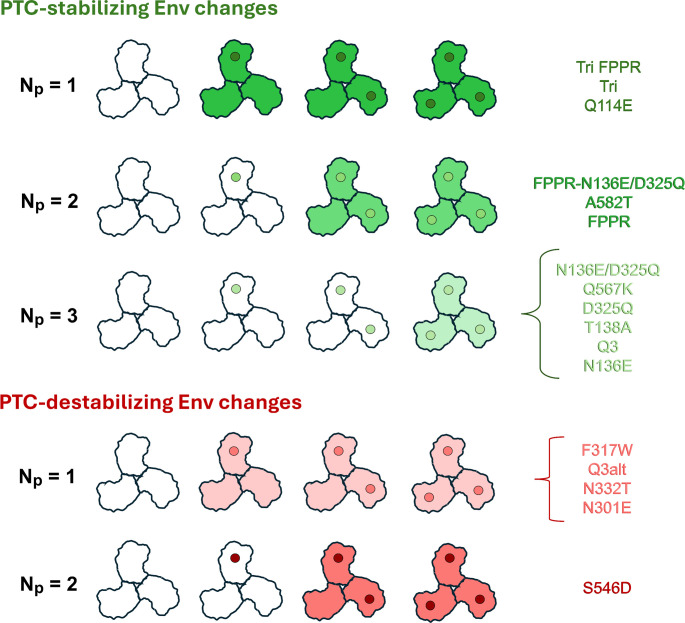 Fig 9
Fig 9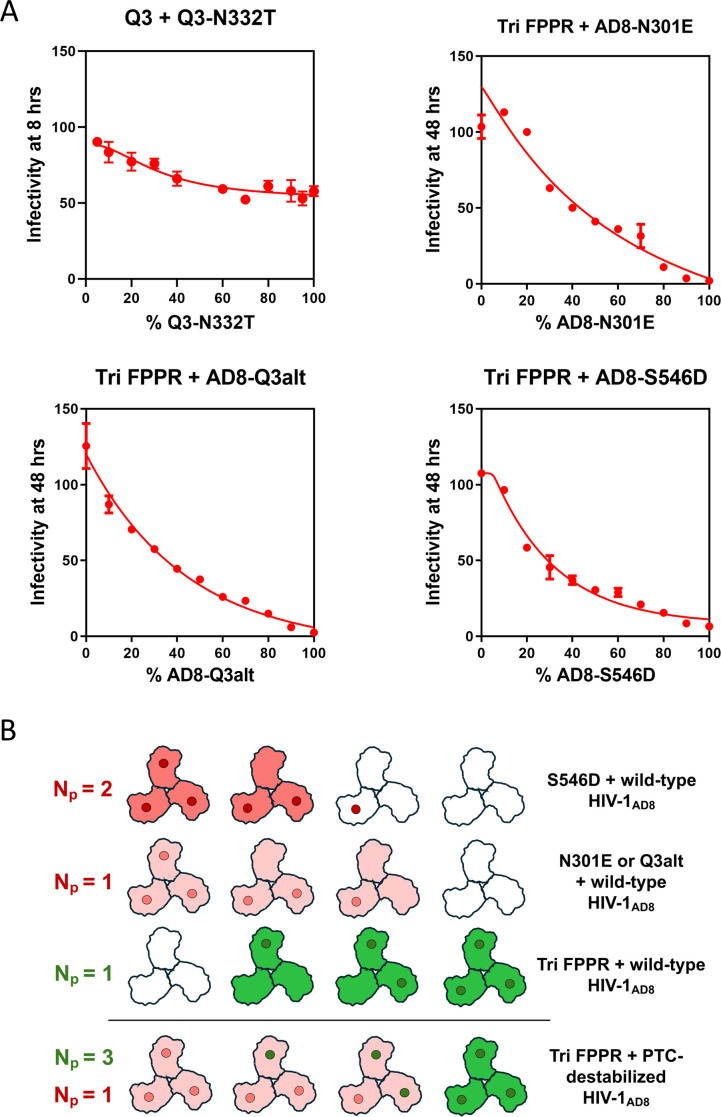 Fig 10
Fig 10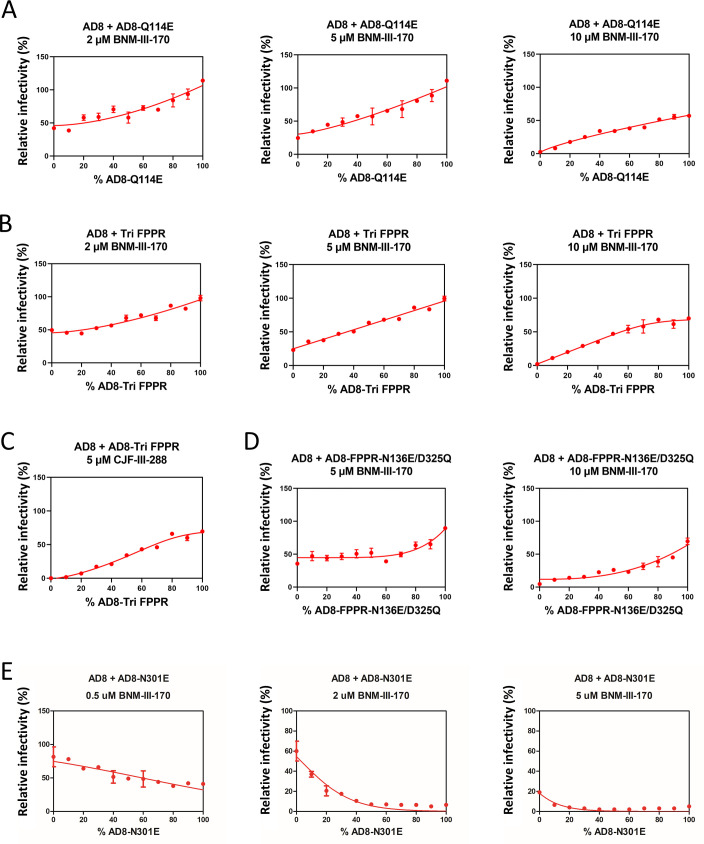 Fig 11
Fig 11| HIV-1AD8 Env | IC50 BNM-III-170 (µM) | Pretriggered Env stability index | Env processing | Reference(s) | |
|---|---|---|---|---|---|
| Wild type | <1 | 4.25 ± 1.47 | 4.25 | +++ | |
| PTC-stabilized Envs | |||||
| N136E | <1 | 3.63 ± 2.69 | <3.63 | +++ | ( |
| Q3 | 1–2 | 2.64 ± 2.20 | 3.96 | +++ | ( |
| T138A | <1 | 7.29 ± 4.74 | <7.29 | +++ | ( |
| D325Q | <1 | 8.03 ± 2.32 | <8.03 | +++ | ( |
| Q567K | 1–2 | 6.38 ± 2.69 | 9.57 | +++ | ( |
| FPPR | 1–2 | 6.87 ± 6.07 | 10.3 | +++ | ( |
| N136E/D325Q | 1–2 | 10.03 ± 9.19 | 15.0 | +++ | ( |
| A582T | 3–5 | 11.75 ± 4.67 | 47.0 | +++ | ( |
| Q114E | >5 | >17 | >85 | +++ | ( |
| FPPR N136E/D325Q | >5 | >21 | >105 | ++ | ( |
| Tri | >5 | >39 | >195 | ++++ | ( |
| Tri FPPR | >5 | >50 | >250 | ++++ | ( |
| PTC-destabilized Envs | |||||
| F317W | <1 | 1.58 ± 0.37 | <1.58 | +++ | ( |
| Q3alt | <1 | 1.10 ± 0.17 | <1.10 | +++ | ( |
| Q3-N332T | <1 | 0.99 ± 0.50 | <0.99 | ++++ | ( |
| N332T | <1 | 0.48 ± 0.39 | <0.48 | ++++ | ( |
| N301E | <1 | 0.25 ± 0.08 | <0.25 | +++ | ( |
| S546D | <1 | 0.16 ± 0.07 | <0.16 | +++ | ( |
| HIV-1AD8 Env | Neutralization (mean IC50 ± SD) by: | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| bNAbs | pNAbs | sCD4-Ig | ||||||||
| VRC01 | VRC03 | b12 | PGT145 | PGT151 | 10E8.v4 | 447-52D | 19b | 17b | ||
| Wild type | 2.54 ± 1.12 | 0.28 ± 0.10 | 1.42 ± 0.47 | 0.42 ± 0.13 | 0.28 ± 0.06 | >10 | >50 | >50 | >50 | 30.5 ± 5.02 |
| F317W | 3.29 ± 0.05 | 0.34 ± 0.02 | 1.46 ± 0.43 | 0.79 ± 0.30 | 0.53 ± 0.29 | >10 | >50 | >50 | >50 | 15.8 ± 0.73 |
| N332T | 0.59 ± 0.33 | 0.05 ± 0.04 | 0.23 ± 0.16 | 0.15 ± 0.12 | 0.09 ± 0.06 | 2.05 ± 2.21 | >12 | >50 | >50 | 6.38 ± 1.23 |
| N301E | 0.58 ± 0.18 | 0.10 ± 0.02 | 0.25 ± 0.15 | 0.51 ± 0.29 | 0.20 ± 0.10 | 5.86 ± 4.44 | 7.37 ± 6.35 | >50 | >50 | 3.25 ± 1.21 |
| S546D | 0.56 ± 0.46 | 0.10 ± 0.09 | 0.04 ± 0.03 | 0.20 ± 0.16 | 0.13 ± 0.08 | 0.13 ± 0.12 | 0.07 ± 0.07 | 0.03 ± 0.03 | 0.16 ± 0.18 | 0.26 ± 0.13 |
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHIV Research and Treatment · HIV/AIDS drug development and treatment · Mosquito-borne diseases and control
INTRODUCTION
The human immunodeficiency virus (HIV-1) envelope glycoprotein (Env) trimer is composed of three protomers, each of which consists of a gp120 exterior Env and a gp41 transmembrane Env (1–4). During HIV-1 entry into host cells, gp120 binds the receptors, CD4 and CCR5/CXCR4, and gp41 mediates the fusion of the viral and target cell membranes (1, 2, 5–10). The Env trimer is metastable, and CD4 binding induces conformational changes from the pretriggered (state 1) conformation (PTC) to more “open” default intermediate (state 2) and full CD4-bound (state 3) conformations (11–13). CCR5/CXCR4 binding to the full CD4-bound state triggers additional conformational changes that lead to the formation of a very stable gp41 six-helix bundle (1, 2, 14–16). The free energy difference between the metastable PTC and the six-helix bundle is used to drive the fusion of the viral and target cell membranes (1, 2, 16).
Many HIV-1 biological properties are determined by Env triggerability or reactivity, defined as the propensity of Env to undergo transitions from the PTC to states 2/3 (12, 17, 18). The metastable pretriggered Env resides in a local energy well, and the height of the activation energy barrier separating the PTC and the default intermediate state is inversely related to Env triggerability (12, 17, 18). Env triggerability governs the virus requirements for levels of CD4 on target cells, sensitivity to soluble CD4 and CD4-mimetic compounds (CD4mcs), and susceptibility to inactivation by prolonged exposure to cold (0°C) (12, 17–19). Primary HIV-1 strains exhibit a range of Env triggerabilities (12, 17, 18). Some natural polymorphisms in Env amino acid residues stabilize or destabilize the PTC (12, 17, 20–30). In several cases, combinations of individual PTC-stabilizing amino acid changes in Env result in additive viral phenotypes (20, 24, 25, 28, 29). This additive property has allowed the creation of HIV-1 Env variants that are stabilized in the PTC to a degree beyond that of natural virus strains (25). These extreme examples exhibit very stable Env trimers that are resistant to activation/inactivation by sCD4 and CD4mcs and are less efficient at supporting cell-cell fusion and virus entry (25).
Here, we address the following question: How many Env protomers must be modified by PTC-stabilizing or -destabilizing changes for the functional Env trimer to achieve stabilization or destabilization, respectively, of the PTC? By evaluating the phenotypes of viruses with mixed Env trimers, we find that the number of protomers that need to be modified to achieve PTC stabilization depends upon the degree of PTC stability achieved by the introduced Env change(s). Strongly PTC-stabilizing Env changes need to be present in only a single protomer to stabilize the pretriggered Env trimer, revealing interprotomer cooperativity in regulating PTC stability. On the other hand, weakly PTC-stabilizing changes must be present in all three protomers to achieve stabilization of the PTC. The presence of most destabilizing Env changes in a single protomer was sufficient to disrupt the functional pretriggered Env. These complementary results provide valuable insights into the symmetric nature of the functional pretriggered Env conformation and the interprotomer relationships that govern its maintenance and conversion into downstream conformations.
RESULTS
HIV-1AD8 Env mutants with alterations in the stability of the PTC
HIV-1 Env triggerability/reactivity is a continuous variable, inversely related to the activation energy required to move Env from its metastable pretriggered conformation (PTC) (12, 17, 18). We assembled a panel of well-characterized HIV-1_AD8_ Env mutants exhibiting a range of triggerabilities, including those with unusually low triggerability (high PTC stability) and those with high triggerability (low PTC stability) (Table 1) (20, 22, 24, 25). In this set of closely matched HIV-1_AD8_ mutants, virus resistance to cold (0°C) exposure and to CD4mcs has been shown to correlate closely with the degree of PTC stabilization (20, 24, 25). Cold resistance is a particularly useful surrogate measure of PTC stability for these stoichiometric studies as the assay avoids the use of an Env ligand whose binding could potentially be affected by the introduced Env amino acid changes (see Fig. 1A for example). The HIV-1_AD8_ Env variants selected for study exhibit a wide range of sensitivities to cold inactivation and inhibition by a CD4mc, BNM-III-170 (Table 1) (31, 32). The pretriggered Env stability index, which correlates directly with PTC stability and inversely with Env triggerability (20, 24, 25), is the product of the half-life of the viral infectivity at 0°C and the IC_50_ values for BNM-III-170. In Table 1, the PTC-stabilized and PTC-destabilized Env variants are ranked according to their pretriggered Env stability indices, which exhibit >1,500-fold variation in this panel. The diversity of this panel of HIV-1_AD8_ Env variants will allow evaluation of the impact of differences in the pretriggered Env stability index on Env protomer stoichiometry.
Cold sensitivity, expression, and processing of representative Env variants. (A) The cold sensitivity of recombinant HIV-1 pseudotyped with the indicated Env variants is shown. HEK293T cells were transfected with the luciferase-expressing proviral plasmid, pNL4-3.Luc.R-E-, a Tat-expressing plasmid, and pSVIIIenv plasmids expressing the indicated wild-type and mutant HIV-1AD8 Envs. The pseudoviruses produced in the cell supernatants were incubated on ice (0°C) for the indicated lengths of time and then used to infect TZM-bl cells. After 2 days, the luciferase activity in the TZM-bl cell lysates was measured. The luciferase activity measured for each virus was normalized to that observed for the same virus sample that was not incubated on ice. The results of a typical experiment are shown, with means and standard deviations derived from triplicate measurements. (B) HOS cells were transfected with plasmids expressing Tat and the wild-type (wt) and mutant HIV-1AD8 Envs. Seventy-two hours later, the cells were lysed, and the gp120 glycoprotein in the cell medium was captured on Galanthus nivalis lectin (GNL)-beads. The clarified cell lysates and GNL precipitates were Western blotted with a polyclonal goat anti-gp120 antibody (Invitrogen).
Most of the Env mutants selected for this study were expressed and processed at least as well as the wild-type HIV-1_AD8_ Env (Table 1; Fig. 1B). Some of the Envs (Tri, Tri FPPR, N332T, and Q3-N332T) were processed more efficiently than the wild-type HIV-1_AD8_ Env. Only one mutant (FPPR N136E/D325Q) was processed less efficiently than the wild-type HIV-1_AD8_ Env. The potential impact of differences in Env processing will be considered in the interpretation of the results (see below).
Infectivity of recombinant HIV-1 with mixed Env trimers
Our general approach measures the infectivity of single-round recombinant HIV-1 expressing luciferase and pseudotyped with mixed Env trimers. The predicted infectivity of viruses with mixed Envs can potentially be influenced by the number of Env trimers (T) required for infection, i.e., the number of Env trimers in the functional unit of virus entry. To obtain information about the entry stoichiometry of the Envs in our study, we measured the infectivities of pseudoviruses containing mixtures of selected HIV-1_AD8_ Envs that vary in PTC stability and a dominant-negative, cleavage-defective Env mutant, R508S/R511S (34–37). In addition to the wild-type HIV-1_AD8_ Env, we selected two PTC-stabilized Envs (Tri and Q114E) and two PTC-destabilized Envs (N301E and S546D) for study. The Env content of the viruses with mixed Envs is shown in Fig. 2A. The amounts of Env were comparable in most of the virus preparations. As previously observed (20, 25), the processing of the Tri Env was more efficient than that of the other Envs.
Entry stoichiometry of viruses pseudotyped with selected PTC-stabilized or PTC-destabilized Envs. (A) Recombinant HIV-1 pseudotyped with mixtures of the indicated Envs and the dominant-negative, cleavage-defective HIV-1AD8 R508S/R511S Env were pelleted, lysed, and Western-blotted with a goat anti-gp120 polyclonal antibody (Invitrogen). The percentages of the plasmids expressing the indicated Envs in the mixture are shown above the blot. The results of independent experiments with the wild-type (wt) HIV-1AD8 Env, designated wt AD8 (1) and wt AD8 (2), are shown in panels A and B. (B) Recombinant viruses containing the indicated Envs mixed with different percentages of the R508S/R511S Env were used to infect TZM-bl cells. Forty-eight hours later, luciferase activity in the target cells was measured. The relative infectivity represents the infectivity observed for each virus normalized to the infectivity of the virus with 100% of the indicated Env and no R508S/R511S Env. (C) The processing, infectivity, susceptibility to inhibition by the dominant-negative R508S/R511S Env, and estimated T values are shown for the wild-type HIV-1AD8 Env, two PTC-stabilized Envs (Tri and Q114E), and two PTC-destabilized Envs (N301E and S546D). The values of T deduced by fitting the f-infectivity curves in panel B to the theoretical curves for the nt = T and nt = T + 1 models in panel D are reported. The pretriggered Env stability index loosely correlates with the fraction (f) of the R508S/R511S Env mutant at which 50% inhibition was achieved. (D) Theoretical curves describing the relative infectivity as a function of f, the fraction of Envs represented by a dominant-negative Env mutant. The curves are influenced by T (the number of Env trimers required for virus entry) and nt (the number of functional Env trimers on a virion available to participate in a productive infection). Examples are shown of models where nt = T (solid lines) and nt = T + 1 (broken lines).
The infectivities of the viruses with mixed Envs for TZM-bl target cells are shown in Fig. 2B. The inclusion of increasing amounts of the dominant-negative R508S/R511S Env resulted in decreases in infection for all the viruses. However, differences in the decreases in infectivity were observed for the viruses depending on the Env paired with the R508S/R511S Env. As the fraction (f) of the R508S/R511S Env in the mixture increased, the infectivity of the viruses decreased in the following order: S546D > N301E > wild type AD8 > Q114E > Tri. The susceptibility of these Env partners to the inclusion of the dominant-negative R508S/R511S mutant in the virus mixture is suggestive of an inverse relationship with their pretriggered Env stability indices (Fig. 2C).
By making several assumptions, the above results can provide information about the entry stoichiometry of the Env variants with altered PTC stability. We assume the following: (i) random mixing of the dominant-negative Env and partner Envs and (ii) complete inhibition of the function of mixed Env trimers containing at least one R508S/R511S protomer. These assumptions are reasonable based on experimental observations (34–39). Accurate determination of T requires additional difficult-to-prove assumptions about virion Env functionality, distribution, mobility, and ability to participate in productive Env clusters (35–37, 40–43). These parameters are herein encompassed by the variable nt, which generally describes the number of functional Env trimers on virions that are capable of participating in the virus entry process.
Some analyses, recognizing that retroviral preparations exhibit very low infectivity:particle ratios (35, 44–46), have assumed that nt = T on most infectious particles. In this case, inactivation of Env trimer function by the dominant-negative Env is linearly related to decreases in viral infectivity; the residual infectivity is related to the fraction (f) of the dominant-negative Env by the following equation:
Using these assumptions, the f-infectivity curves for the viruses with mixed wild-type AD8 and R508S/R511S Envs suggested T values of 2–3 (Fig. 2C and D). Viruses with the PTC-stabilized Envs (Tri and Q114E) exhibited f-infectivity curves suggestive of T values of 1–2. The f-infectivity curves of the PTC-destabilized Envs suggested higher T values (3–4 for N301E and 6 for S546D).
We also considered scenarios where nt > T, i.e., where more functional Env trimers are present on most virions than are required for virus entry. In the case where nt = T + 1, the f-infectivity curves assume sigmoidal shapes (Fig. 2D). The T values predicted by fitting the experimental curves to the theoretical f-infectivity curves when nt = T + 1 are greater than the T values predicted when nt = T (Fig. 2C).
Uncertainty about n_t_ precludes a definitive determination of T. Nonetheless, the pattern of resistance to the dominant-negative Env suggests that Env PTC stability is associated with an increase in the nt/T ratio. This ratio is an indicator of the redundancy of functional Envs on the virion particle. PTC stabilization could potentially increase nt, lower T, or both.
The results of the dominant-negative Env mixing experiments established boundaries for the T and nt values of our Env mutants. We consider the effects of these estimates of T and nt in our analyses of the protomer stoichiometry of PTC stabilization and destabilization below.
Protomer stoichiometry of Envs with PTC-stabilizing changes
To estimate the number of protomers (Np) required for PTC stabilization, we generated single-round recombinant viruses pseudotyped with mixtures of the wild-type HIV-1_AD8_ Env and PTC-stabilized Envs in varying proportions. The phenotypes of the viruses with 100% PTC-stabilized Env mutants define the maximum degrees of PTC stabilization achievable by the viruses with mixed Envs. Relative to this maximum, the percentage (p) of Env trimers in the population that achieve a stabilized PTC is a function of the fraction (f) of PTC-stabilized mutant Envs in the mixture, the number of protomers (Np) required for PTC stabilization, and the number of Env trimers (T) in the functional unit. As discussed above, an accurate determination of T depends upon currently unknown parameters. Therefore, we considered a range of T values in our evaluation of the Env protomer stoichiometry associated with PTC stabilization/destabilization. Figure 3 shows the theoretical curves governing PTC stabilization for T values of 1, 2, and 3. The predicted values of p determine the expected infectivities of viruses with mixed Env trimers, after cold exposure, according to the equation:
Theoretical analysis of HIV-1 Env protomer stoichiometry for PTC stabilization. The random mixing of wild-type Env protomers (open circles) with PTC-stabilized mutant Env protomers (filled circles) according to a binomial distribution is depicted for T values of 1 and 2, where T represents the number of Env trimers required for virus entry. For T = 2, the number of permutations of mixed trimers is indicated adjacent to the parentheses. The percentage (p) of maximal PTC stabilization achieved depends on the number of Env protomers (Np) required for PTC stabilization, on the number of Env trimers required for virus entry (T), and on the fraction (f) of PTC-stabilized mutant Envs. The formulas and plots of p as a function of f are shown for the different values of Np and T. In the models shown, nt = T. We assume that all T Env trimers within the functional unit of virus entry must be stabilized for that functional unit to achieve PTC stability. For each value of Np, the Env trimer combinations that are considered to result in PTC stability are boxed. The variable p represents the sum of probabilities for the Env protomer/trimer configurations that result in PTC stability. Note that the Np = 1, Np = 2, and Np = 3 curves in the f-p graphs form distinct, readily distinguishable groups. The predicted infectivity of the virus after cold exposure is determined by p and by the infectivities of the viruses with 100% wild-type Env and 100% mutant Env, according to the equation shown.
Our theoretical analyses of PTC stabilization assume the following: (i) a random mixing of wild-type and mutant Envs according to a binomial distribution, (ii) all-or-none Env trimer function, and (iii) a direct relationship between the percentage (p) of functional Env trimers in the population and virus infectivity. The last assumption is valid when nt = T; we consider the effects of higher nt values on the estimation of Np below. Given these assumptions, Fig. 3 shows the ideal curves describing the relationship between f and p. The shapes of these ideal curves qualitatively differ for Np = 1, 2, and 3. Although increasing the value of T shifts the curves and adds slightly to their sigmoidicity, the Np = 1, Np = 2, and Np = 3 curves form distinct groups and can easily be distinguished at all T values. Thus, plotting the empirically observed viral infectivities after cold exposure as a function of f and comparison with the theoretical curves predicted by equation 2 should allow us to deduce the Np values.
Recombinant viruses with mixtures of the wild-type HIV-1_AD8_ Env and PTC-stabilized variant Envs were generated. The infectivity of these viruses was measured after incubation on ice (0°C) for a period of time that resulted in maximal differences between the infectivities of the viruses with 100% wild-type HIV-1_AD8_ Env and 100% mutant Env. Plots of the residual virus infectivity after cold exposure as a function of f (the fraction of mutant Envs) revealed three patterns (Fig. 4). Convex upward f-infectivity curves were observed for three Env variants, Q114E, Tri, and Tri FPPR. The Tri and Tri FPPR Envs have multiple PTC-stabilizing changes, one of which is Q114E (20, 24, 25). The shapes of these f-infectivity curves are consistent with Np = 1, indicating that the presence of these PTC-stabilizing changes in a single protomer is sufficient to achieve cold resistance. For three of the Env variants (FPPR-N136E/D325Q, A582T, and FPPR), the f-infectivity curves were consistent with Np = 2, suggesting that the presence of these PTC-stabilizing changes in two protomers is sufficient to confer cold resistance. A third pattern of concave upward f-infectivity curves was observed for half (6 out of 12) of the PTC-stabilizing changes. These f-infectivity curves indicate that the PTC-stabilizing changes need to be present in all three Env protomers to achieve cold resistance (Np = 3). These results indicate that the number of Env protomers that need to be modified to achieve cold stability of the virus differs for various PTC-stabilizing Env changes.
Cold sensitivity of viruses with mixed wild-type and PTC-stabilized Envs. HEK293T cells were transfected with the luciferase-expressing proviral plasmid, pNL4-3.Luc.R-E-, a Tat-expressing plasmid, and pSVIIIenv plasmids expressing the wild-type and mutant HIV-1AD8 Envs in the indicated proportions. The pseudoviruses produced in the cell supernatants were incubated on ice (0°C) for the indicated time points and then used to infect TZM-bl cells. Two days later, the luciferase activity in the TZM-bl cell lysates was measured. The luciferase activity measured for each virus was normalized to the luciferase activity observed for the same virus preparation that had not been incubated on ice. The means and standard deviations derived from triplicate measurements within a typical experiment are shown. The PTC-stabilized Env variants are grouped according to the Np values deduced from the shape of the f-infectivity curves. As the infectivities of the AD8 + AD8-Tri and AD8 + AD8-Tri FPPR viruses plateau by f = 50%, only the portions of the graphs from f = 0–50% are shown.
High levels of expression and processing of mutant Envs relative to those of wild-type Env could hypothetically lower our estimate of Np. As some PTC-stabilized Env mutants like Tri and Tri FPPR exhibit relatively efficient processing and low estimated Np values (20, 25) (Fig. 1B; Table 1), we wished to evaluate whether mixing these mutants with wild-type HIV-1_AD8_ Env might have biased the outcome. We examined the Envs in pseudoviruses produced by titrating increasing amounts of Tri and Tri FPPR Envs into the wild-type HIV-1_AD8_ Env (Fig. 5A). The levels and processing of the mixed Envs with increasing proportions of Tri and Tri FPPR Envs were similar, particularly when these values were normalized to the levels of Gag p24 capsid protein in the virions. These observations argue against systematic alterations of virion Env levels or processing as a result of mixing the Tri and Tri FPPR Envs with the wild-type HIV-1_AD8_ Env.
Characterization of viruses with mixed wild-type and PTC-stabilized Envs. (A) Pseudoviruses with the indicated proportions of wild-type and mutant HIV-1AD8 Envs were prepared as described in the Fig. 4 legend. Clarified cell supernatants were centrifuged at 14,000 × g to pellet the virus particles. Equal volumes of resuspended virus particles were Western-blotted with a polyclonal goat anti-gp120 antibody (upper panels), the 4E10 anti-gp41 antibody (middle panels), and a polyclonal rabbit antibody against Gag p55/p24/p17 (lower panels). The results of a typical experiment are shown. (B) The theoretical effects of changes in nt (the number of functional Env trimers on a virion that can potentially participate in the virus entry process) on the relationship between f and p are shown for different Np values. The percentage (p) of maximal PTC stabilization achieved is plotted as a function of the fraction (f) of PTC-stabilized mutant Envs. The formulas and f-p graphs are shown for T = 1, where T represents the number of Env trimers required for virus entry.
We also considered how differences in nt might affect the outcome of the mixing experiments. For virions that are potentially infectious, the value of nt must be greater than or equal to T. For any given T, as nt increases, there is an increased probability of forming a group of stabilized Env trimers for entry. Therefore, at a given value of f, an increase in nt results in an increase in p and greater Env stabilization (Fig. 5B). A comparison of the bottom panel of Fig. 3 with Fig. 5B shows that increases in T and nt exert opposite effects on the f-p relationship. These opposing effects may diminish the impact of large T values on the estimation of Np. Finally, we note that the greater-than-expected positive stabilizing effects of PTC-stabilized Envs like Tri and Tri FPPR (Fig. 4, upper panel) may be a result of high nt values (Fig. 5B).
Notably, for our panel of PTC-stabilizing Env changes, the Np values deduced from the analysis of the f-infectivity curves inversely correlated with the pretriggered Env stability indices (Spearman rS = −0.865, two-tailed P = 0.0006) (Fig. 6). Thus, the number of Env protomers that need to be modified to achieve stabilization of the PTC is inversely related to the degree of PTC stabilization that results from the Env change. Weakly and moderately PTC-stabilizing changes need to be present in multiple protomers (Np = 3 or 2, respectively) to achieve cold resistance. Strongly PTC-stabilizing Env changes efficiently stabilize the functional PTC against the effects of cold exposure when present in a single protomer.
Correlation between the number of protomers (Np) required for PTC stabilization and the pretriggered Env stability index. The inverse relationship between the Np values deduced from the shapes of the f-infectivity curves in Fig. 4 and the pretriggered Env stability indices (Table 1) is shown. The Spearman rank correlation coefficient (rS) and two-tailed P value are shown.
Protomer stoichiometry of Envs with PTC-destabilizing changes
For mixtures of the wild-type HIV-1_AD8_ Env and mutant Envs with PTC-destabilizing changes, we generated theoretical curves describing the relationship between f (the fraction of mutant Envs) and the percentage (p) of maximal PTC destabilization achieved (Fig. 7). The assumptions (random Env mixing and all-or-none Env trimer function) underlying these calculations are similar to those described above for the mixtures of wild-type and PTC-stabilized Envs. In addition, for T > 1 and nt = T, we assumed that destabilization of any one of the Env trimers in the functional unit of virus entry results in PTC destabilization of that functional unit. As seen for the mixtures with the PTC-stabilized Envs, distinct shapes of the f-p curves were associated with the different Np values. At higher T values, which may apply to the PTC-destabilized Env mutants (Fig. 2C), the f-p curves shift but do so uniformly, retaining the separation into distinct Np groups (Fig. 7). For a given f value, as T increases, the efficiency of Env PTC destabilization is expected to increase; in contrast, increases in nt should decrease the efficiency of Env PTC destabilization. Thus, increases in T and nt may partially cancel each other’s effect on our estimation of Np. However, at high actual T values, we could potentially underestimate Np, a point that is discussed further below.
Theoretical analysis of HIV-1 protomer stoichiometry for PTC destabilization. The random mixing of wild-type Env protomers (open circles) with PTC-destabilized mutant Env protomers (filled circles) according to a binomial distribution is depicted for T values of 1 and 2, where T represents the number of Env trimers required for virus entry. For T = 2, the number of permutations of mixed trimers is indicated adjacent to the parentheses. The percentage (p) of maximal PTC destabilization achieved depends on the number of Env protomers (Np) required for PTC destabilization, on the number of Env trimers required for virus entry (T), and on the fraction (f) of PTC-destabilized mutant Envs. The formulas and plots of p as a function of f are shown for the different values of Np and T. In the models shown, nt = T. For T > 1, we assume that destabilization of any one of the Env trimers in the functional unit of virus entry results in PTC destabilization of that functional unit. For each value of Np, the Env trimer/protomer combinations that are considered to result in PTC destabilization are boxed. The variable p represents the sum of probabilities for the Env protomer/trimer configurations that result in destabilization of the PTC. Note that the Np = 1, Np = 2, and Np = 3 curves in the f-p graphs form distinct, readily distinguishable groups. The predicted infectivity of the virus after cold exposure is determined by p and by the infectivities of the viruses with 100% wild-type Env and 100% mutant Env, according to the equation shown.
Recombinant viruses with mixtures of wild-type HIV-1_AD8_ Env and PTC-destabilized Env mutants were generated and tested for cold sensitivity. In Fig. 8, we show the observed f-infectivity curves, allowing comparison with the theoretical curves (in the lower right panel). For most of the Env mutants, the presence of the destabilizing changes in a single protomer was apparently sufficient to destabilize the PTC and render the viruses sensitive to incubation at 0°C. A protomer stoichiometry of Np = 1 was deduced from the cold sensitivity plots of the Q3alt, N301E, F317W, and N332T Env mutants. Of interest, three of these PTC-destabilizing changes (Q3alt, N301E, and N332T) involve the loss or shift of an N-linked glycosylation site in gp120 (24). The F317W change alters the tip of the gp120 V3 loop (30).
Cold sensitivity of viruses with mixed wild-type and PTC-destabilized Envs. The cold sensitivity of pseudoviruses with mixed wild-type and mutant HIV-1AD8 Envs was evaluated as described in the Fig. 4 legend. The means and standard deviations derived from triplicate measurements within a typical experiment are shown. The experiments were repeated with comparable results. The PTC-destabilized Env variants are grouped according to the Np values deduced from the shape of the f-infectivity curves. Theoretical f-infectivity curves for viruses with mixtures of wild-type Env and a PTC-destabilized Env mutant are shown in the bottom right panel. These theoretical curves apply when the infectivity of viruses with the wild-type Env is 100 and the infectivity of viruses with the mutant Env is 0, following incubation on ice. For other cases, the infectivity of viruses with mixed Envs can be predicted by the equation in Fig. 7. In these models, nt = T.
The f-infectivity curve associated with another PTC-destabilizing change, S546D, suggested that its presence in at least two Env protomers is necessary to destabilize the Env trimer and render the virus sensitive to cold (Fig. 8). Serine 546 is located in the gp41 heptad repeat (HR1_N_) region, and the S546D change does not alter a potential N-linked glycosylation site. Thus, the S546D change differs from the other destabilizing Env mutants tested in the subunit location in the Env trimer, protomeric stoichiometry, and, in some cases, effect on glycosylation.
To evaluate potential relationships between the degree of PTC destabilization and the protomeric stoichiometry (Np) of the S546D and the other tested Env variants, we compared the sensitivity of viruses with these Envs to cold, CD4mcs, sCD4-Ig, and antibodies. The results are shown in [Tables 1 and 2](#T1 T2), where the PTC-destabilized Env mutants are arranged in order of decreasing pretriggered Env stability indices. The sensitivity of viruses with these Envs to sCD4-Ig exhibits the same rank order (Table 2); PTC destabilization is expected to increase Env triggerability and inactivation by sCD4-Ig (12, 17, 19, 23–25). The F317W virus exhibited a twofold increase in sensitivity to sCD4-Ig, but was neutralized by the antibodies tested comparably to the wild-type HIV-1_AD8_ virus. Viruses with the other PTC-destabilized Envs (N332T, N301E, and S546D) were more sensitive than the wild-type virus to CD4BS bNAbs, the 10E8.v4 MPER bNAb, and the 447-52D anti-V3 pNAb. The S546D virus was not only more sensitive to these antibodies than the N332T and N301E viruses but also was neutralized by the 19b anti-V3 and 17b CD4i pNAbs. Thus, compared with the other PTC-destabilizing Env changes, the S546D change results in the highest degree of PTC destabilization, based on the viral pretriggered Env stability indices and sensitivities to antibody/sCD4-Ig neutralization. In this set of PTC-destabilized Env variants, greater PTC destabilization is apparently associated with a higher Np requirement.
The deduced protomer stoichiometries (Np values) and the relationship of the observed phenotypes to the degree of PTC stabilization/destabilization for the panel of studied Env variants is summarized in Fig. 9.
Summary of deduced protomer stoichiometries for PTC-stabilizing and PTC-destabilizing Env changes. A schematic diagram of the HIV-1 Env trimer is shown with PTC-stabilizing changes (green dots) and PTC-destabilizing changes (red dots) in the protomers. The dots are shaded according to the degree of PTC stabilization or destabilization achieved when the respective PTC-stabilizing or PTC-destabilizing Env change is present in all three protomers; the pretriggered Env stability index provides an indication of the degree of PTC stabilization or destabilization achieved by the introduced changes. The specific Env changes assigned to each Np category are listed on the right. Because of the strong relationship between the deduced Np values and the pretriggered Env stability indices (Fig. 6), the Env changes are roughly ranked from highest to lowest pretriggered Env stability index. The deduced phenotypes of the functional Env trimers are indicated by shading, with cold-resistant trimers in green and cold-sensitive trimers in red. The intensity of the Env trimer shading relates to the degree of PTC stabilization or destabilization.
Combination of PTC-stabilizing and -destabilizing changes in Env trimers
We wished to examine the effect of PTC-destabilizing Env changes in the context of an Env trimer that has PTC-stabilizing changes in some or all of its protomers. Several variations were tried.
Q3 + Q3 N332T: in the background of the wild-type HIV-1_AD8_ Env, the N332T change in one protomer (Np = 1) is sufficient for destabilization of the PTC and increased cold sensitivity (Fig. 8). The magnitude of the cold sensitivity phenotype resulting from the N332T change was reduced when the N332T change was introduced into an Env where all three protomers had the PTC-stabilizing Q3 change (Fig. 10A). Nonetheless, the shape of the f-infectivity curve indicated that the protomer stoichiometry remained at Np = 1.Tri FPPR + AD8-N301E: in the wild-type HIV-1_AD8_ background, the N301E change destabilizes the PTC when present in a single protomer (Fig. 8). Likewise, when the AD8-N301E Env was mixed with the Tri FPPR Env, one protomer of AD8-N301E was sufficient to destabilize the PTC of the Env trimer (Fig. 10A).Tri FPPR + AD8-Q3alt: in the wild-type HIV-1_AD8_ background, the Q3alt change in a single protomer is sufficient to destabilize the PTC (Fig. 8). This was also the case when the AD8-Q3alt Env was mixed with the Tri FPPR Env (Fig. 10A).Tri FPPR + AD8-S546D: in the wild-type HIV-1_AD8_ background, two S546D protomers are required to destabilize the PTC of Env (Fig. 8). However, when the AD8-S546D Env was mixed with the Tri FPPR Env, one AD8-S546D protomer was apparently able to bring about PTC destabilization (Fig. 10A).
Cold sensitivity of viruses with mixed PTC-stabilized and PTC-destabilized Envs. (A) The cold sensitivity of pseudoviruses with mixed PTC-stabilized and PTC-destabilized Envs was evaluated, as described in the Fig. 4 legend. The means and standard deviations derived from triplicate measurements within a typical experiment are shown. The experiments were repeated with comparable results. (B) A schematic diagram of the HIV-1 Env trimer is shown with the PTC-stabilizing Tri FPPR changes (green dots) and the PTC-destabilizing (N301E, Q3alt, and S546D) changes (red dots) in the protomers. The Env variants in the mixed virus Envs are shown on the right. The viral Env mixtures above the horizontal line include the wild-type HIV-1AD8 Env as a partner and thus correspond to those depicted in Fig. 9. The deduced Np values of the Tri FPPR Env (green) and the PTC-destabilized Envs (red) are indicated on the left. The deduced phenotypes of the functional Env trimers are indicated by shading, with cold-resistant trimers in green and cold-sensitive trimers in red. Wild-type HIV-1AD8 cold sensitivity is indicated by white/unshaded trimers. Note that compared to the mixtures with wild-type HIV-1AD8 Env, when the Envs with the PTC-stabilized Tri FPPR changes are mixed with the PTC-destabilized Envs, Np = 1 for the PTC-destabilized Envs and the Np value of Tri FPPR increases to 3.
In all four examples above, the presence of a single protomer with a PTC-destabilizing change apparently resulted in PTC destabilization of Env trimers with at least two protomers containing PTC-stabilizing changes. This is not surprising in the case where the PTC-stabilizing change is Q3, which even when present in all three protomers only weakly stabilizes the Env PTC. However, it is unexpected in the three instances with the strongly PTC-stabilizing Tri FPPR changes that, when mixed with wild-type HIV-1_AD8_ protomers, stabilize the PTC when present in a single protomer (Np = 1) (Fig. 10B). Instead, when Tri FPPR protomers are mixed with Env protomers with PTC-destabilizing changes, Env trimers with one or two Tri FPPR protomers are cold-sensitive. Apparently, the Np = 1 stoichiometry of the PTC-destabilizing changes is dominant over the Np = 1 stoichiometry of the PTC-stabilizing Tri FPPR change. The above examples also demonstrate that the Env background in which PTC-destabilizing changes are introduced may preserve or alter the protomer stoichiometry (Np).
Sensitivity of Env chimeras to CD4mcs
The measurement of viral cold sensitivity affords a ligand-free means of evaluating the HIV-1 Env conformational state, thus providing insight into the spontaneous triggerability of Env. Virus sensitivity to CD4mc inhibition provides a second surrogate for the stability of the PTC, allowing an assessment of induced triggerability. Measuring the protomer stoichiometry of Env changes in the latter case is more technically challenging, as multiple CD4mc concentrations must be empirically evaluated to achieve detectable but different levels of infection for the wild-type and mutant viruses. We determined the protomer stoichiometry of three PTC-stabilizing Env changes (Q114E, Tri FPPR, and FPPR-N136E/D325Q), using virus inhibition by the CD4mc BNM-III-170 as a phenotypic readout. None of these PTC-stabilizing changes overlaps with the known gp120 binding site of the CD4mcs (31, 32, 47). We mixed HIV-1_AD8_ Envs containing each of these changes with the wild-type HIV-1_AD8_ Env and tested the susceptibility of the viruses to inhibition by different concentrations of BNM-III-170. As expected (20, 25), compared with viruses with the wild-type HIV-1_AD8_ Env, the viruses with 100% PTC-stabilized Envs were relatively resistant to BNM-III-170 inhibition. For the strongly PTC-stabilized Q114E and Tri FPPR Envs, the f-infectivity curves exhibited mild shape changes as the BNM-III-170 concentration varied; these may reflect the effects of CD4mc occupancy of the gp120 subunits on the stoichiometric requirements for the PTC-stabilizing changes (Fig. 11A and B). However, the f-infectivity curves for both Envs deviated only modestly from the Np = 2 theoretical curve at all CD4mc concentrations. An Np = 2 stoichiometry was also supported by the f-infectivity curve observed for the PTC-stabilizing Tri FPPR changes when a more potent CD4mc, CJF-III-288 (47), was used (Fig. 11C). For the moderately PTC-stabilizing FPPR-N136E/D325Q Env, the f-infectivity curves were consistent with Np = 3 at multiple BNM-III-170 concentrations (Fig. 11D). Thus, for CD4mc resistance, we assign N_p_ values of 2, 2, and 3 for the Q114E, Tri FPPR, and FPPR-N136E/D325Q Envs, respectively. The respective Np values for the cold resistance of these mutants were 1, 1, and 2. Apparently, more protomers require PTC-stabilizing changes to achieve resistance to BNM-III-170 than to increase resistance to cold. It may be easier for PTC-stabilizing changes to resist spontaneous Env transitions from the PTC than the transitions induced by CD4mcs.
CD4mc inhibition of viruses with PTC-stabilized (or PTC-destabilized) Envs mixed with the wild-type HIV-1AD8 Env. Pseudoviruses with mixed wild-type and mutant HIV-1AD8 Envs were prepared as described in the Fig. 4 legend. The viruses were incubated with the indicated concentrations of the CD4mc BNM-III-170 (A, B, D, and E) or the more potent CD4mc CJF-III-288 (C) for 1 hour at 37°C. The virus-CD4mc mixture was then incubated with TZM-bl cells for 2 days, after which luciferase activity in the target cells was measured. The measured luciferase activity was normalized to that observed for the same virus preparation that had not been treated with the CD4mc. Multiple concentrations of the CD4mc were tested to identify the concentration(s) at which the differences between the wild-type and mutant Envs would permit conclusions about the Env protomer stoichiometry. In panels A, B, D, and E, the results obtained with different concentrations of BNM-III-170 are shown. The means and standard deviations derived from triplicate measurements within a typical experiment are shown. The experiments were repeated with comparable results.
We also wished to evaluate how the presence of a PTC-destabilizing change in one or more Env protomers would affect virus sensitivity to BNM-III-170. For this purpose, we mixed HIV-1_AD8_ Envs containing the destabilizing N301E change with the wild-type HIV-1_AD8_ Env and tested the susceptibility of the viruses to inhibition by BNM-III-170. As expected (24), viruses with 100% AD8-N301E Envs were inhibited to a greater extent by BNM-III-170, consistent with their increased triggerability. At low concentrations of BNM-III-170, the viruses with two or more AD8-N301E protomers were inhibited (Fig. 11E). At higher BNM-III-170 concentrations, Envs with one protomer of the AD8-N301E Env were efficiently inhibited. Apparently, the CD4mc-induced inactivation of Env is more efficient when two protomers contain the PTC-destabilizing N301E change. Fewer Env protomers with this PTC-destabilizing change are required for virus inactivation when CD4mc concentrations are higher.
DISCUSSION
In this work, we investigated the number of Env protomers (Np) that must be modified by particular Env changes that stabilize or destabilize the pretriggered conformation (PTC) to achieve the viral phenotypes of increased resistance or sensitivity, respectively, to cold (0°C) or CD4mc exposure. Cold exposure and incubation with CD4mcs drive HIV-1 Env from the metastable PTC by different means: cold inactivation is ligand-free and spontaneous, whereas CD4mcs bind gp120 and induce changes in Env similar to those triggered by CD4 binding (11, 12, 19, 23, 26, 31, 32, 47–50). Increases or decreases in the stability of the Env PTC result in increased HIV-1 resistance or sensitivity, respectively, to both cold and CD4mcs (12, 17–26, 48). The product of the virus half-life on ice and the CD4mc IC_50_ is the pretriggered Env stability index, which correlates with the stability of the PTC better than either factor on its own (25). We used the pretriggered Env stability indices to rank the HIV-1_AD8_ Env mutants in this study with respect to the degree of PTC stabilization or destabilization achieved.
As the Env trimer stoichiometry (T) associated with virus entry can hypothetically influence our estimation of Np, we studied the infectivity of viruses with mixtures of the wild-type HIV-1_AD8_ Env and a dominant-negative, cleavage-defective Env (R508S/R511S) (34–37). The R508S/R511S mutant inhibited the infectivity of viruses with PTC-destabilized Envs more efficiently than the infectivity of viruses with PTC-stabilized Envs. PTC stabilization could lower T (the number of Env trimers required for virus entry) or increase nt (the number of functional Env trimers on virions that can participate in virus entry) or both. Lower T values have been suggested for the Envs of primary HIV-1 than for the Envs of laboratory-adapted HIV-1, which typically exhibit lower PTC stability (18, 37). Determination of actual T values for HIV-1 is complicated by uncertainty about virion Env numbers, distribution, mobility, and ability to contribute to productive entry events, all of which are encompassed in the variable nt. For the purpose of this work, we identified a range of T and nt values that we evaluated for the potential impact on our estimation of Np. The relatively low T and nt values associated with the PTC-stabilized Envs were not predicted to confound our ability to deduce Np values from the f-infectivity curves for these Env mutants. Indeed, in the experiments involving the PTC-stabilized Envs, we observed all three shapes of the f-infectivity curves predicted for the three Np values. Moreover, we observed a strong inverse correlation between the deduced Np values and the pretriggered Env stability indices of the PTC-stabilized Envs. These results argue against severe violations of the assumptions underlying our theoretical analysis of the PTC-stabilized Envs.
The ligand-free cold inactivation assay allowed us to evaluate the cold sensitivity of viruses pseudotyped with mixtures of a great variety of Envs and to estimate the protomer stoichiometry, Np. With respect to cold resistance of the viruses, the number of Env protomers that must be modified to achieve stabilization of the PTC was found to correlate inversely with the degree of PTC stabilization that results from the introduced Env changes (Fig. 6). For strongly PTC-stabilizing Env changes, modification of a single protomer appears to be sufficient to achieve PTC stabilization (Np = 1). This observation suggests that given adequate stability, the PTC-stabilized protomer can influence the conformation of the other two protomers to maintain the PTC. The efficacy of such cross-protomer cooperativity is dependent on the Env partner that is mixed with the strongly PTC-stabilized Env. Thus, for the strongly PTC-stabilizing Tri FPPR changes, Np = 1 when mixed with the wild-type HIV-1_AD8_ Env, but Np = 3 when the Env partner contains PTC-destabilizing changes (N301E, Q3alt, or S546D) (Fig. 10B). In the latter cases, the presence of even a single protomer with the PTC-destabilizing changes completely nullified the ability of the Tri FPPR changes to render the mixed Env trimers cold-resistant. Apparently, disruption of the PTC of a single Env protomer is not readily compensated even by strongly PTC-stabilizing changes in the other protomers. In this situation, the protomer stoichiometries of PTC-destabilizing Env changes are dominant.
For weakly PTC-stabilizing Env changes, all three protomers required modification to achieve cold resistance of the virus (Np = 3). Thus, symmetrical placement of weakly PTC-stabilizing changes in three Env protomers is apparently conducive to the maintenance of the PTC (Fig. 9). A complementary observation is that the presence of weakly PTC-destabilizing Env changes in one protomer was sufficient to increase the cold sensitivity of the virus (Np = 1). The PTC of the wild-type HIV-1_AD8_ Env can be disrupted by the asymmetric introduction of a PTC-destabilizing Env change into a single protomer. This PTC-destabilizing Np = 1 stoichiometry dominated, even when strongly PTC-stabilizing changes were present in the other protomers. In current Env structures (51–56), several of the strongly PTC-stabilizing changes involve amino acid residues that are near subunit interfaces. However, PTC-destabilizing changes often involve changes in N-linked glycosylation sites and are located on the periphery of the Env trimer (see below). Thus, direct interactions between Env structures involved in PTC stabilization and destabilization are unlikely explanations for our observations. A more attractive model explaining these observations is that the PTC represents a C3-symmetric trimer. In this model, Env changes that stabilize the PTC need to maintain conformational symmetry among the protomers. Weakly PTC-stabilizing Env changes, lacking the ability of the strongly PTC-stabilizing changes to act across protomers, must be present in all three protomers (Np = 3). Conversely, PTC-destabilizing Env changes in one protomer can lead to a significant loss of conformational symmetry among the protomers, thereby destabilizing the PTC, with consequent increases in virus sensitivity to cold inactivation and CD4mc inhibition. Supporting this model are the many examples of weakly PTC-stabilizing Env changes where Np = 3 and weakly PTC-destabilizing Env changes where Np = 1 (Fig. 9).
Several of the weakly PTC-stabilizing changes (N136E, T138A, N136E/D325Q, and Q3) involve the loss of a gp120 V1 glycan at Asn 136, which presumably exerts its effects at the Env surface (24, 25). Other weakly PTC-stabilizing changes (Q567K and D325Q) do not alter a potential Env glycosylation site. Apparently, these changes most effectively stabilize the PTC when they are present in all three Env protomers, consistent with the stabilization of a symmetric Env trimer.
For the group of strongly PTC-stabilizing Env mutants (Q114E, Tri, and Tri FPPR), all of which share the Q114E change, alteration of a single Env protomer is apparently sufficient to achieve cold resistance. In available Env trimer models (51–56), Gln 114 is located in the α1 helix of the gp120 inner domain, near the interface with gp41. The mechanism whereby a change in Gln 114 to glutamic acid in one protomer would stabilize the PTC of the entire Env trimer is not apparent in currently available structures. Detailed structures of the membrane Env PTC may shed light on this gap in our understanding.
For most of the PTC-destabilizing Env changes, alteration of a single Env protomer (Np = 1) was apparently sufficient to sensitize the virus to cold inactivation. Loss of a glycan in one protomer (Np = 1) destabilized the PTC in the case of the N301E and N332T changes, which remove glycans that occupy the interprotomer angles of the Env trimer (24). An Np value of 1 was also deduced for the F317W change in the gp120 V3 tip, which does not alter an N-linked glycosylation site. The protomer stoichiometry of these Env variants supports the hypothesis that C3 symmetry contributes to the maintenance of the pretriggered (state 1) Env trimer conformation (Fig. 9). Recent structural studies suggest that the default intermediate (state 2) Env is an asymmetric trimer, which presumably derives from a symmetric pretriggered (state 1) Env (51, 52, 57–59).
One PTC-destabilizing change, S546D, needed to be present in two or more protomers (Np = 2) to render the virus cold-sensitive. The sensitivity of viruses with the S546D Env to inhibition by the dominant-negative, cleavage-defective Env raises the possibility that, due to higher T, we underestimated Np and the actual Np = 3. In either case, the S546D Np estimates are greater than those for other PTC-destabilizing Env changes. An obvious structural explanation is lacking for the higher Np requirements of this mutant. Serine 546 is located in the highly dynamic gp41 HR1_N_ region, near the trimeric coiled coil formed by HR1_C_ (51–56, 58). The higher Np value of S546D, relative to those of other PTC-destabilizing changes, may reflect the magnitude of PTC destabilization achieved; the global sensitivity of the S546D mutant to antibody neutralization suggests that significant redistribution into state 3-like Envs results from this change.
To achieve HIV-1_AD8_ resistance to CD4mcs, the PTC-stabilizing changes were required in more Env protomers than for cold resistance. Apparently, it is more difficult to resist the direct induction by CD4mcs of entry-related transitions from the PTC than to withstand the more generally disruptive effects of ice formation on Env trimer integrity associated with cold exposure (49, 50). With respect to CD4mc sensitivity, a PTC-destabilizing change, N301E, exhibited different protomer stoichiometries at low (Np = 2) and high (Np = 1) concentrations of the CD4mc. The increased occupancy of Env at higher CD4mc concentrations itself promotes Env transitions from the PTC and likely accounts for the lower protomer requirements for the PTC-destabilizing change.
Our studies of Env protomer stoichiometry suggest the importance of conformational correspondence among the protomers of functional Env trimers. Future efforts will evaluate the hypothesized contribution of Env structural symmetry to PTC integrity and should facilitate further stabilization and characterization of the pretriggered (state 1) Env conformation.
MATERIALS AND METHODS
HIV-1 env mutants
The wild-type HIV-1_AD8_ env cloned in the pSVIIIenv expression plasmid was used as a template to construct HIV-1 Env mutants in this study (20). The signal peptide/N-terminus (residues 1–33) and the cytoplasmic tail C-terminus (residues 751–856) of this Env are derived from the HIV-1_HXBc2_ Env. The dominant-negative, cleavage-defective Env mutant has two changes, R508S/R511S, that eliminate proteolytic processing of the Env precursor (34–36). “Tri” indicates the Q114E/Q567K/A582T changes, and “FPPR” indicates the A532V/I535M/L543Q changes (20, 24, 25). The Q3 and Q3 alt Env mutants were previously reported as Q3 (V1) and Q3 (V1alt), respectively, in reference 24. Env mutants with specific changes were generated by using the QuikChange Lightning site-directed mutagenesis kit (Agilent Technologies). All the Envs contain a His_6_ tag at the carboxyl terminus. The presence of the desired mutations was confirmed by DNA sequencing.
Cell lines
HEK293T, TZM-bl, and HOS cells (ATCC) were cultured in Dulbecco modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 IU penicillin and 100 μg/mL streptomycin (Life Technologies).
Env expression
To evaluate Env processing and subunit association, 3 × 10^5^ HOS cells were seeded in 6-well plates. After 24 hours of incubation, they were transfected with plasmids encoding His_6_-tagged Env variants and Tat at a ratio of 8:1, using the Lipofectamine 3000 transfection reagent (Life Technologies) according to the manufacturer’s instructions. Seventy-two hours after transfection, the cells were lysed in PBS buffer containing 1.0% NP-40 and protease inhibitor (Sigma-Aldrich). Clarified lysates were harvested, boiled, and analyzed by Western blotting with 1:2,500 goat anti-gp120 antibody (Invitrogen) and 1:2,500 HRP-conjugated rabbit anti-goat antibody (Invitrogen). The intensities of the gp120 and gp160 bands from unsaturated Western blots were quantified by using ImageJ software. The Env processing index was calculated by dividing gp120 by gp160 in the cell lysate samples. The processing indices of Env mutants in this study were normalized to those of the wild-type HIV-1_AD8_ Env.
Seventy-two hours after transfection, the supernatants of the transfected HOS cells were collected and incubated with Galanthus Nivalis Lectin (GNL)-agarose beads (Vector Laboratories) for 1.5 h at room temperature. The beads were washed three times with PBS containing 0.1% NP-40 and processed for Western blotting with 1:2,500 goat anti-gp120 antibody (Invitrogen), as described above. The subunit association index was calculated by dividing gp120 in the cell lysate samples by the gp120 in the GNL precipitates. The subunit association indices of the Env mutants were normalized to those of the wild-type HIV-1_AD8_ Env.
Virus infectivity
To produce pseudoviruses, HEK293T cells were cotransfected with Env-expressing pSVIIIenv plasmids, a Tat-encoding plasmid and the luciferase-encoding pNL4-3.Luc.R-E- vector (NIH HIV Reagent Program) at a 1:1:3 ratio using polyethyleneimine (PEI, Polysciences). To investigate Env trimer stoichiometry (T), the pSVIIIenv plasmids encoded mixtures of selected PTC-stabilized and PTC-destabilized Envs with varying proportions of the dominant-negative, cleavage-defective R508S/R511S Env mutant (34–36). To investigate Env protomer stoichiometry (Np), the pSVIIIenv plasmids encoded mixtures of the wild-type HIV-1_AD8_ Env and either PTC-stabilized or PTC-destabilized Env mutants in varying proportions. After 8 hours, the medium was replaced with fresh medium. Seventy-two hours later, the pseudoviruses in the supernatants were harvested and centrifuged (3,500 rpm for 5 min), aliquoted, and either used directly to measure pseudovirus infectivity or stored at −80°C until further use. To evaluate the infectivity of the variants, pseudoviruses were diluted in 96-well plates and incubated with TZM-bl cells in the presence of 20 µg/mL DEAE-dextran. After a 48 hour incubation at 37°C in 5% CO_2_, the TZM-bl cells were lysed, and the luciferase activity was measured using a luminometer.
Virus sensitivity to cold inactivation
To evaluate the sensitivity of viruses with mixed Envs to cold inactivation, pseudoviruses were incubated on ice (0°C) for a period of time. This period of time was chosen to maximize the infectivity difference between viruses with wild-type HIV-1_AD8_ Env and PTC-stabilized/-destabilized mutant Envs. After cold incubation, the infectivity of the pseudoviruses was measured as described above. The level of infectivity of each virus following incubation on ice was normalized to that of the same virus preparation that had not been incubated on ice.
Virus inhibition by a CD4mc
The CD4mcs BNM-III-170 and CJF-III-288 (31, 32, 47) were serially diluted in triplicate wells in 96-well plates. Then approximately 100 to 200 TCID_50_ (50% tissue culture infectious dose) of pseudoviruses was added and incubated at 37°C for 1 h. Subsequently, approximately 2 × 10^4^ TZM-bl cells with 20 µg/mL DEAE-dextran in the medium were added to each well, and the mixture was incubated at 37°C/5% CO_2_ for 48 hours. Then, luciferase activity was measured, as described above. The level of infectivity of each virus with mixed Envs was normalized to that of the same virus preparation that had not been incubated with CD4mcs.
Analysis of Env on virus particles
Approximately 1 mL of the clarified cell supernatant containing pseudovirus was centrifuged at 14,000 × g for 1 h at 4°C. The pelleted virus particles were resuspended in 1 × PBS. Equal volumes of the virus suspensions were then lysed in PBS buffer/1.0% NP-40/protease inhibitor cocktail and analyzed by Western blotting. Western blots were developed with 1:2,500 goat anti-gp120 polyclonal antibody (Invitrogen), 1:2,500 4E10 anti-gp41 antibody, and 1:5,000 rabbit polyclonal antibody against Gag p55/p24/p17 (Abcam). The HRP-conjugated secondary antibodies were 1:2,500 rabbit anti-goat antibody (Invitrogen), 1:2,500 goat anti-human antibody (Invitrogen), and 1:5,000 goat anti-rabbit antibody (Sigma-Aldrich), respectively.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wyatt R, Sodroski J. 1998. The HIV-1 envelope glycoproteins: fusogens, antigens, and immunogens. Science 280:1884–1888. doi:10.1126/science.280.5371.18849632381 · doi ↗ · pubmed ↗
- 2Chen B. 2019. Molecular mechanism of HIV-1 entry. Trends Microbiol 27:878–891. doi:10.1016/j.tim.2019.06.00231262533 PMC 6744290 · doi ↗ · pubmed ↗
- 3Allan JS, Coligan JE, Barin F, Mc Lane MF, Sodroski JG, Rosen CA, Haseltine WA, Lee TH, Essex M. 1985. Major glycoprotein antigens that induce antibodies in AIDS patients are encoded by HTLV-III. Science 228:1091–1094. doi:10.1126/science.29862902986290 · doi ↗ · pubmed ↗
- 4Robey WG, Safai B, Oroszlan S, Arthur LO, Gonda MA, Gallo RC, Fischinger PJ. 1985. Characterization of envelope and core structural gene products of HTLV-III with sera from AIDS patients. Science 228:593–595. doi:10.1126/science.29847742984774 · doi ↗ · pubmed ↗
- 5Klatzmann D, Champagne E, Chamaret S, Gruest J, Guetard D, Hercend T, Gluckman JC, Montagnier L. 1984. T-lymphocyte T 4 molecule behaves as the receptor for human retrovirus LAV. Nature 312:767–768. doi:10.1038/312767 a 06083454 · doi ↗ · pubmed ↗
- 6Dalgleish AG, Beverley PCL, Clapham PR, Crawford DH, Greaves MF, Weiss RA. 1984. The CD 4 (T 4) antigen is an essential component of the receptor for the AIDS retrovirus. Nature 312:763–767. doi:10.1038/312763 a 06096719 · doi ↗ · pubmed ↗
- 7Wu L, Gerard NP, Wyatt R, Choe H, Parolin C, Ruffing N, Borsetti A, Cardoso AA, Desjardin E, Newman W, Gerard C, Sodroski J. 1996. CD 4-induced interaction of primary HIV-1 gp 120 glycoproteins with the chemokine receptor CCR-5. Nature 384:179–183. doi:10.1038/384179 a 08906795 · doi ↗ · pubmed ↗
- 8Trkola A, Dragic T, Arthos J, Binley JM, Olson WC, Allaway GP, Cheng-Mayer C, Robinson J, Maddon PJ, Moore JP. 1996. CD 4-dependent, antibody-sensitive interactions between HIV-1 and its co-receptor CCR-5. Nature 384:184–187. doi:10.1038/384184 a 08906796 · doi ↗ · pubmed ↗