RfaH is essential for virulence and adaptive responses in Yersinia pseudotuberculosis infection
Joram Kiriga Waititu, Kristina Nilsson, Gerald Larrouy-Maumus, Tiago R. D. Costa, Kemal Avican

TL;DR
This study shows that RfaH is crucial for Yersinia pseudotuberculosis to cause infection and adapt to stress in the host.
Contribution
The study identifies RfaH as a key regulator of virulence and adaptive responses in Y. pseudotuberculosis beyond O-antigen regulation.
Findings
Loss of RfaH significantly reduces Y. pseudotuberculosis virulence and its ability to establish persistent infection.
RfaH influences a broader set of genes related to virulence and adaptation, beyond O-antigen biosynthesis.
RfaH expression increases under stress conditions like high osmolarity and bile salts.
Abstract
We previously suggested that increased expression of the gene encoding transcriptional antiterminator RfaH during Yersinia pseudotuberculosis transcriptional reprogramming is necessary for adapting to persistent infection. In this study, we examined the role of RfaH in virulence and bacterial physiology under infection-relevant stress conditions, and identified genes differentially regulated in the absence of RfaH in Y. pseudotuberculosis. We employed a mouse infection model and phenotypic assays to test RfaH’s role in virulence and physiology, as well as RNA sequencing, including O-antigen biosynthesis-deficient strains. Our findings demonstrate that loss of RfaH significantly attenuates virulence, reducing the capacity of Y. pseudotuberculosis to establish persistent infection. RfaH expression is increased during the stationary growth phase and under various stress conditions, such as…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
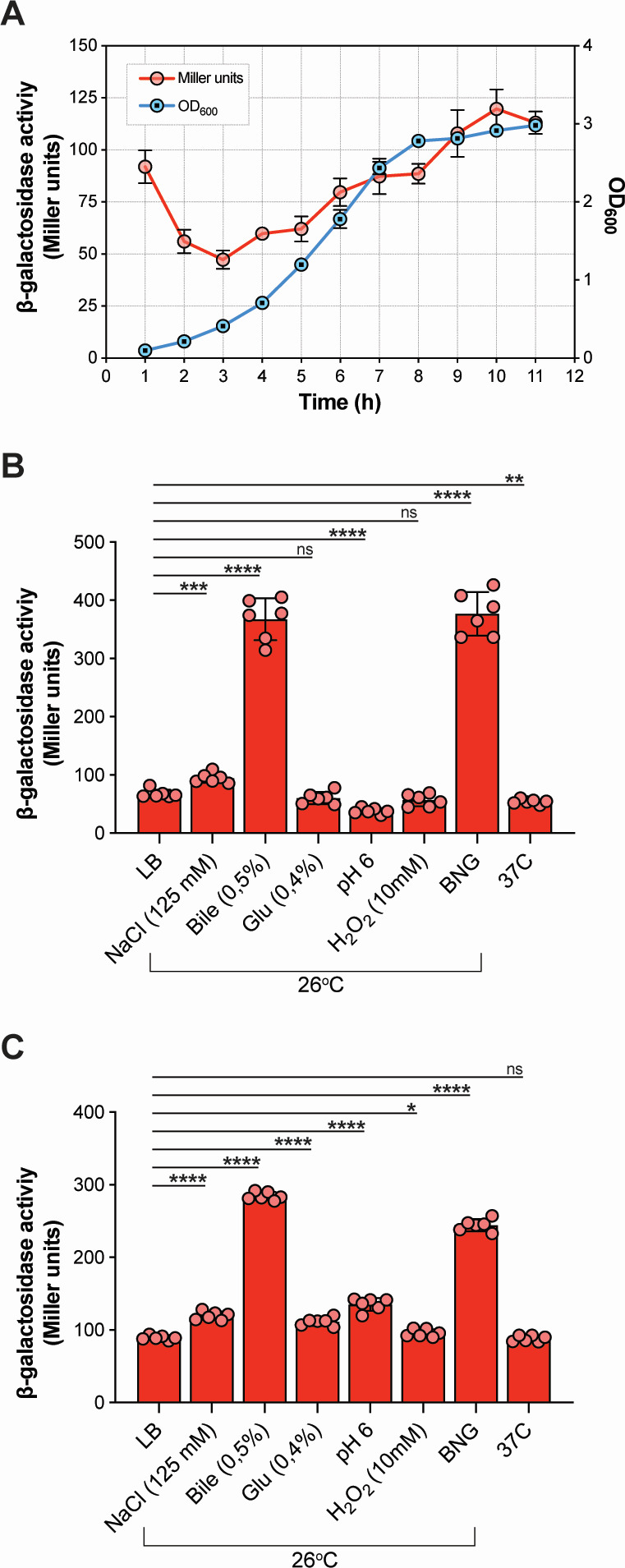 Fig 1
Fig 1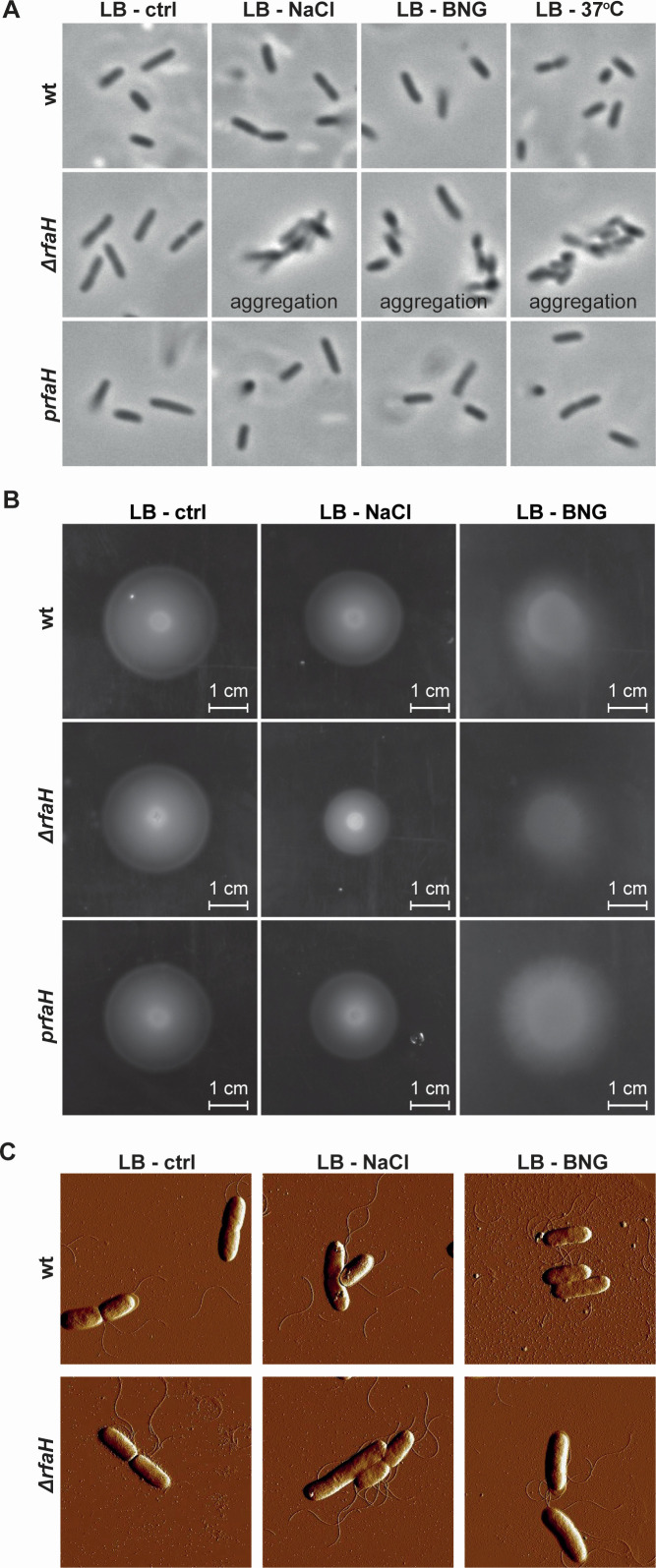 Fig 2
Fig 2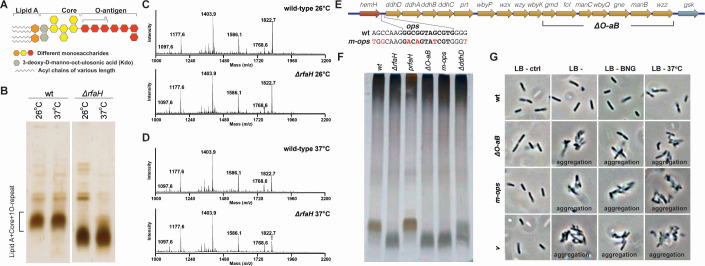 Fig 3
Fig 3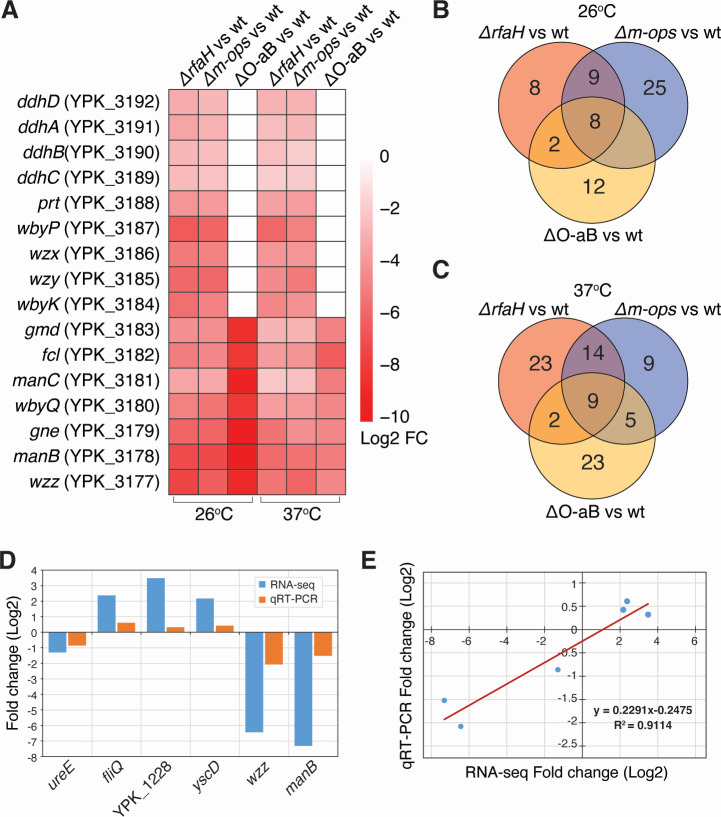 Fig 4
Fig 4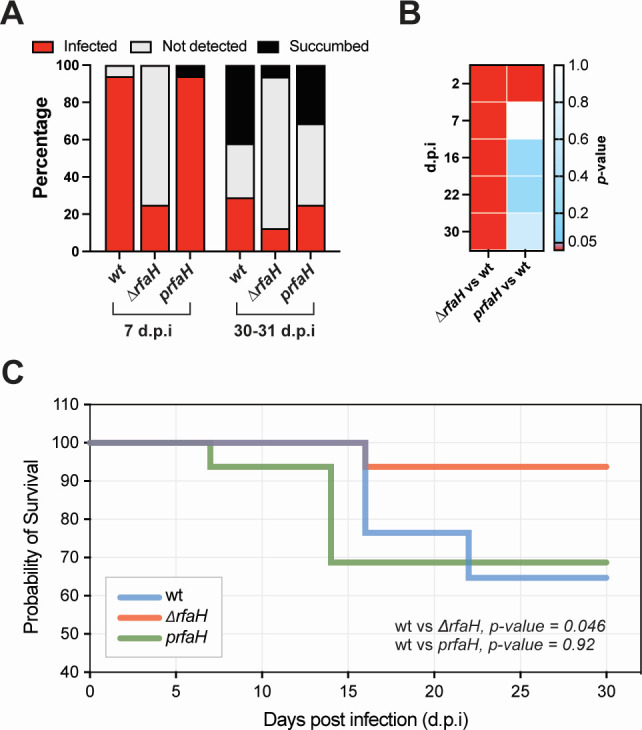 Fig 5
Fig 5| Locus | Gene | Description | Log2 fold change (26°C) | Log2 fold change (37°C) | ||||
|---|---|---|---|---|---|---|---|---|
| Δ |
| Δ | Δ |
| Δ | |||
| YPK_3937 |
| Transcriptional activator RfaH | −8.47 | — | — | −6.68 | — | — |
| YPK_0406 | Putative cytoplasmic protein | −3.10 | — | — | — | — | — | |
| YPK_4249 |
| 50S ribosomal protein L34 | −1.74 | — | — | — | — | — |
| YPK_0582 | Putative adhesin/hemolysin | −1.14 | — | — | — | — | — | |
| YPK_2421 |
| Flagellar basal body rod protein FlgF | −1.10 | — | — | — | — | — |
| YPK_1408 |
| Sulfate ABC transporter | −1.05 | — | — | — | −1.03 | — |
| YPK_1920 |
| Putative lipoprotein | 1.09 | — | — | — | — | — |
| piBX002 | Transposase | 1.96 | — | — | —- | — | — | |
| YPK_1388 |
| Nitrate reductase cytochrome c-type subunit | — | −1.64 | — | — | — | — |
| YPK_0379 | Glycosidase | — | −1.33 | — | — | — | — | |
| YPK_1134 |
| Urease accessory protein UreE | — | −1.30 | — | — | — | — |
| YPK_1133 |
| Urease subunit alpha, UreC | — | −1.28 | — | — | — | — |
| YPK_1137 |
| Urease accessory protein UreD | — | −1.13 | — | — | — | — |
| YPK_1135 |
| Urease accessory protein UreF | — | −1.09 | — | — | — | — |
| YPK_1136 |
| Urease accessory protein UreG | — | −1.04 | — | — | — | — |
| YPK_3760 |
| Putative transcriptional regulator Nlp | — | −1.28 | — | — | — | — |
| YPK_4113 |
| Glycerol kinase | — | −1.04 | — | — | — | — |
| YPK_RS05760 | Hypothetical protein | — | −1.06 | — | — | — | — | |
| YPK_3632 | Hypothetical protein | — | −1.04 | — | — | — | — | |
| piBX084 | Lipoprotein Virg | -— | 1.03 | — | — | — | — | |
| YPK_3390 |
| Cytochrome b561 | — | 1.03 | — | — | — | — |
| YPK_3948 |
| Carbon starvation protein CstA | — | 1.07 | — | — | — | — |
| YPK_0725 |
| Putative transcriptional regulator CadC | — | 1.19 | — | — | — | — |
| YPK_0617 | AntA/AntB antirepressor domain-containing protein | — | 1.33 | — | — | — | — | |
| YPK_1316 | Insertion element protein | — | 4.02 | — | — | — | — | |
| YPK_1642 | Hypothetical protein | — | 1.10 | — | — | — | — | |
| YPK_1153 | Hypothetical protein | — | 1.14 | — | — | — | — | |
| piBX059 | Hypothetical protein | — | 1.24 | — | — | — | — | |
| YPK_2209 | Hypothetical protein | — | 1.28 | — | — | — | — | |
| YPK_3090 | Hypothetical protein | — | 3.68 | — | — | — | — | |
| YPK_1734 | Hypothetical protein | — | 1.39 | — | — | — | — | |
| YPK_2076 | Hypothetical protein | — | 2.41 | — | — | — | — | |
| YPK_0840 | Hypothetical protein | — | 3.48 | — | — | — | — | |
| YPK_0192 | Hypothetical protein | — | — | −3.64 | 3.58 | 4.50 | 3.56 | |
| YPK_RS05880 | Hypothetical protein | — | — | −3.52 | — | — | — | |
| YPK_RS07695 | Hypothetical protein | — | — | −2.46 | — | — | — | |
| YPK_0193 | Hypothetical protein | — | — | −1.56 | — | — | — | |
| YPK_0848 | Transposase IS200 family protein | — | — | −1.23 | 1.40 | 1.07 | — | |
| YPK_4237 | Extracellular solute-binding protein | — | — | −1.04 | — | — | — | |
| YPK_1972 | Hypothetical protein | — | — | 1.20 | — | — | — | |
| YPK_3963 | Hypothetical protein | — | — | 1.54 | — | — | — | |
| YPK_RS20560 | Hypothetical protein | — | — | 1.95 | — | — | — | |
| YPK_1458 | Hypothetical protein | — | — | 2.38 | — | — | — | |
| YPK_RS05195 | Hypothetical protein | — | — | 2.46 | — | — | — | |
| YPK_2234 | Transposase IS200-family protein | — | — | 4.95 | — | — | — | |
| YPK_RS11665 | Holin | — | — | — | −3.40 | — | — | |
| YPK_2349 | Tail E family protein | — | — | — | −3.06 | — | — | |
| YPK_3401 | Monosaccharide-transporting ATPase | — | — | — | −1.16 | — | — | |
| YPK_2845 | Hypothetical protein | — | — | — | −4.23 | — | — | |
| YPK_0384 | Hypothetical protein | — | — | — | −4.10 | — | — | |
| YPK_2886 | Hypothetical protein | — | — | — | −3.02 | — | — | |
| YPK_0760 | Hypothetical protein | — | — | — | −2.01 | — | — | |
| YPK_2479 | Hypothetical protein | — | — | — | −1.41 | — | — | |
| YPK_2211 | Hypothetical protein | — | — | — | −1.23 | — | — | |
| YPK_4042 |
| Fimbrial protein | — | — | — | 1.46 | — | — |
| YPK_3140 |
| Phage transcriptional regulator AlpA | — | — | — | 1.18 | — | — |
| YPK_3284 | Type 11 methyltransferase | — | — | — | 1.00 | — | — | |
| YPK_3565 | Type VI secretion protein | — | — | — | 1.28 | — | — | |
| piBX060 | Uncharacterized protein pYV0051 | — | — | — | 1.60 | — | — | |
| YPK_1029 | Hypothetical protein | — | — | — | 1.01 | — | — | |
| YPK_1785 | Hypothetical protein | — | — | — | 1.12 | — | — | |
| YPK_0362 | Hypothetical protein | — | — | — | 1.22 | — | — | |
| YPK_1433 | Hypothetical protein | — | — | — | 1.40 | — | — | |
| YPK_RS18305 | Hypothetical protein | — | — | — | 1.00 | — | — | |
| YPK_2893 | Hypothetical protein | — | — | — | 3.34 | — | — | |
| YPK_0213 | Hypothetical protein | — | — | — | 3.66 | — | — | |
| YPK_0817 | Hypothetical protein | — | — | — | 4.50 | — | — | |
| YPK_0342 |
| Thiamine biosynthesis protein ThiH | — | — | — | — | −1.01 | — |
| YPK_1610 | Alcohol dehydrogenase zinc-binding | — | — | — | — | −1.02 | — | |
| YPK_2130 | Spermidine export protein Mdt | -— | — | — | — | −1.50 | — | |
| YPK_RS09520 | Hypothetical protein | — | — | — | — | 1.70 | — | |
| YPK_2612 | Hypothetical protein | — | — | — | — | 1.76 | — | |
| YPK_0953 | Hypothetical protein | — | — | — | — | 1.81 | — | |
| YPK_0705 |
| Flagellar biosynthetic protein FliQ | — | — | — | — | 2.37 | — |
| YPK_1228 | Hypothetical protein | — | — | — | — | 3.48 | — | |
| YPK_2533 | Transposase IS200 family protein | — | — | — | — | — | −5.66 | |
| YPK_0940 | Small terminase subunit | — | — | — | — | — | −3.61 | |
| YPK_2424 |
| Flagellar basal body rod protein FlgC | — | — | — | — | — | −2.45 |
| YPK_0477 | Putative phage-like protein | — | — | — | — | — | −2.38 | |
| YPK_2395 |
| Flagellar export protein FliJ | — | — | — | — | — | −2.05 |
| YPK_2400 |
| Flagellar motor switch protein FliN | — | — | — | — | — | −1.08 |
| YPK_3933 |
| Fatty oxidation complex, alpha subunit FadB | — | — | — | — | — | −1.00 |
| YPK_1605 |
| Inner membrane transporter RhtA | — | — | — | — | — | −1.37 |
| YPK_0727 | Hypothetical protein | — | — | —— | — | −3.37 | ||
| YPK_1149 | Hypothetical protein | — | — | — | — | — | −2.07 | |
| YPK_3058 | Hypothetical protein | — | — | — | — | — | −2.06 | |
| YPK_3414 | Hypothetical protein | — | — | — | — | — | −1.44 | |
| YPK_1383 | Hypothetical protein | — | — | — | — | — | −1.25 | |
| YPK_0121 |
| Universal stress protein UspB | — | — | — | — | — | 1.05 |
| YPK_3991 | sn-glycerol-3-phosphate dehydrogenase subunit A | — | — | — | — | — | 1.05 | |
| YPK_3201 | Hypothetical protein | — | — | — | — | — | 1.09 | |
| piBX027 | Insertion element ISR1 | — | — | — | — | — | 1.11 | |
| YPK_RS17330 | Hypothetical protein | — | — | — | — | — | 1.13 | |
| YPK_3210 | 50S ribosomal protein L31 | — | — | — | — | — | 1.13 | |
| YPK_1385 |
| Ferredoxin-type protein NapF | — | — | — | — | — | 1.15 |
| YPK_1143 |
| CrcB protein | — | — | — | — | — | 1.49 |
| YPK_2133 | Hypothetical protein | — | — | — | — | — | 2.06 | |
| YPK_3916 |
| YscD/HrpQ, type III secretion apparatus protein | — | — | — | — | — | 2.17 |
| YPK_3177† |
| Ferric enterobactin transport protein FepE | −7.60 | −6.44 | −9.03 | −5.39 | −6.15 | −4.73 |
| YPK_3178† |
| Phosphomannomutase | −7.25 | −7.32 | −10.05 | −5.85 | −5.55 | −5.06 |
| YPK_3179† |
| NAD-dependent epimerase/dehydratase | −6.09 | −6.15 | −10.21 | −5.48 | −4.37 | −4.65 |
| YPK_3180† |
| Glycosyl transferase family 2 protein | −5.04 | −5.44 | −8.23 | −3.77 | −3.97 | −4.70 |
| YPK_3181† |
| Mannose-6-phosphate isomerase | −3.44 | −3.44 | −9.60 | −2.22 | −2.45 | −5.14 |
| YPK_3182† |
| NAD-dependent epimerase/dehydratase | −5.16 | −4.74 | −8.28 | −3.93 | −4.22 | −6.43 |
| YPK_3183† |
| GDP-mannose 4,6-dehydratase | −4.38 | −4.35 | −8.78 | −2.87 | −3.01 | −4.92 |
| YPK_3184† |
| Mannosyltransferase | −5.76 | −4.95 | — | −4.81 | −4.34 | — |
| YPK_3185† |
| O-antigen biosynthesis protein Wxy | −5.99 | −6.05 | — | −4.58 | −5.19 | — |
| YPK_3186† |
| LPS side chain defect: putative O-antigen transferase | −5.18 | −5.05 | — | −4.07 | −4.09 | — |
| YPK_3187† |
| Glycosyl transferase family 2 | −6.61 | −6.09 | —- | −6.02 | −5.05 | — |
| YPK_3188† |
| NAD-dependent epimerase/dehydratase | −4.14 | −3.86 | — | −3.66 | −3.77 | — |
| YPK_3189† |
| CDP-4-dehydro-6-deoxyglucose reductase | −2.69 | −2.38 | — | −1.95 | −2.04 | — |
| YPK_3190† |
| CDP-glucose 4,6-dehydratase | −2.81 | −2.53 | — | −2.43 | −1.97 | — |
| YPK_3191† |
| Glucose-1-phosphate cytidylyltransferase | −3.49 | −2.96 | — | −2.59 | −2.73 | — |
| YPK_3192† |
| Oxidoreductase FAD/NAD(P)-binding domain protein | −3.19 | −2.81 | — | −3.03 | −2.94 | — |
| YPK_0866 | Hypothetical protein | — | — | — | — | −1.88 | −1.09 | |
| YPK_0280 |
| Bacterioferritin-associated ferredoxin | — | — | — | — | −1.39 | −1.20 |
| YPK_1160 | Hypothetical protein | — | — | — | — | 1.01 | 1.19 | |
| YPK_1837 |
| Undecaprenyl phosphate-alpha-L-ara4N flippase | — | — | — | — | 1.24 | 1.21 |
| YPK_3798 | Hypothetical protein | — | — | — | — | 4.62 | 3.67 | |
| YPK_0142 | Hypothetical protein | — | — | — | 1.01 | 1.01 | — | |
| YPK_1770 | Hypothetical protein | — | — | — | 1.19 | 1.35 | —— | |
| YPK_1304 | Type VI secretion system effector, Hcp1 family | — | — | — | 1.72 | 1.23 | — | |
| YPK_3926 | Transposase IS3/IS911 family protein | — | — | — | −22.84 | — | −22.88 | |
| YPK_1861 | Hypothetical protein | — | — | — | −3.19 | - | −4.10 | |
| YPK_1389 |
| Cytochrome c-type protein NapC | −1.44 | −1.35 | −1.51 | — | — | — |
| YPK_1246 | Hypothetical protein | 1.01 | — | 1.29 | — | — | — | |
| YPK_3328 | Polar amino acid ABC transporter | 3.33 | — | 3.39 | 3.51 | 3.22 | — | |
| YPK_0631 | Hypothetical protein | — | — | — | 1.06 | 1.05 | 1.04 | |
- —Vetenskapsrådethttp://dx.doi.org/10.13039/501100004359
- —Vetenskapsrådethttp://dx.doi.org/10.13039/501100004359
- —Kempestiftelsernahttp://dx.doi.org/10.13039/501100007067
- —Umeå Universitethttp://dx.doi.org/10.13039/501100004885
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsYersinia bacterium, plague, ectoparasites research · Bacillus and Francisella bacterial research · Vibrio bacteria research studies
INTRODUCTION
The genus Yersiniae, in the family Enterobacteriaceae, consists of 11 species, of which Yersinia pseudotuberculosis, Y. enterocolitica, and Y. pestis are known to cause diseases in mammals (1–3). Y. pestis, transmitted by fleas, is the causative agent of plague (4), while both Y. pseudotuberculosis and Y. enterocolitica are enteric pathogens that are typically acquired by ingestion of contaminated food or water, leading primarily to self-limiting conditions such as adenitis, enteritis, diarrhea, and ileitis (2, 5, 6). High-dose Y. pseudotuberculosis infection in rodents, such as guinea pigs and mouse models, results in systemic infection due to translocation from the intestinal tract to the spleen and liver (7). The pathogenicity of all three Yersinia species is largely dependent on the type III secretion system and its related virulence substrate proteins, known as Yersinia outer proteins, which are encoded on a 70-kb virulence plasmid (8, 9). Low-dose Y. pseudotuberculosis oral infections cause chronic infection in mouse cecal tissue without signs of disease (10). This model of Y. pseudotuberculosis persistent infection offers insight into bacterial mechanisms of importance for the initiation and maintenance of persistent infections. We have previously explored the nature of Y. pseudotuberculosis persistent infection by profiling transcriptional changes from the early to the later persistent phase of infection (11). The study identified potentially significant key players during persistence, which primarily reflected the environmental conditions the pathogen encountered in the cecal lymphoid follicles. A set of global transcriptional regulators, such as the Crp/CsrA/RovA cascade, regulated gene expression, allowing Y. pseudotuberculosis to adapt to long-term residence in the host (11). Notably, expression of the gene encoding the transcriptional regulator RfaH was significantly enhanced during the persistent state of infection but was not investigated further (11).
RfaH is a transcriptional regulator belonging to the NusG family of proteins (12). It enables the complete transcription of long operons by preventing Rho-dependent premature termination (13, 14). The specificity of RfaH for its target genes depends upon a 12-nucleotide conserved regulatory site called the operon polarity suppressor (ops), which is typically located at upstream regions of the operons regulated by RfaH (15, 16). RfaH prevents early transcriptional termination and enhances transcriptional elongation, ensuring, for example, lipopolysaccharide (LPS) biosynthesis, as shown for many bacteria (17). In Escherichia coli, RfaH acts as a specialized transcription-translation coupling factor that is recruited to RNA polymerase (RNAP) at the ops site, where it bridges RNAP and the lead ribosome via ribosomal protein S10, thereby accelerating transcription elongation rates and blocking Rho-dependent termination (18, 19). This coupling mechanism distinguishes RfaH from NusG, as the activity of the former is restricted to operons containing the ops element (18), and such coupling also contributes to regulatory mechanisms like transcription attenuation and riboswitch control (18, 20, 21). In addition, RfaH has been linked to various roles, including initiation of translation by interacting with the 30S ribosome (12), regulation of operons involved in capsular biosynthesis (22), hemin uptake components (23), and production of toxins like hemolysins and cytotoxic necrotizing factor (24). Indeed, survival of Vibrio vulnificus in serum is dependent on RfaH (25). RfaH has also been implicated in the pathogenesis of various pathogens, where its loss leads to attenuation in virulence of the Salmonella (23), E. coli (26), and Klebsiella (27). In Y. pseudotuberculosis, RfaH has been reported to contribute to resistance against antimicrobial chemokines and survival during mouse infections (28). However, the precise mechanisms by which RfaH enhances the expression of these components remain incompletely understood. Despite significant scientific advances in understanding the RfaH mechanism in various pathogens, knowledge about its molecular mechanisms in microbial pathogenesis, particularly Y. pseudotuberculosis, remains limited (29). In this study, we conducted phenotypic assays to dissect RfaH-dependent changes during bacterial growth and the establishment of infection in a mouse model. Furthermore, we extended our understanding of RfaH’s role in the pathogenesis of Y. pseudotuberculosis through transcriptomic analysis by performing transcriptomic profiling of wt, rfaH deletion mutants, and various RfaH-related mutant strains of Y. pseudotuberculosis at both environmental (26°C) and host body (37°C) temperatures to identify RfaH-regulated genes. We identified common and mutation-specific genes and pathways, shedding light on the potential molecular mechanisms of RfaH in persistent Y. pseudotuberculosis infections. Our findings provide a deeper understanding of RfaH’s role in Y. pseudotuberculosis virulence mechanisms and offer a valuable molecular reference for future studies to understand persistent infections caused by this pathogen.
RESULTS
RfaH expression is growth phase-dependent and can be induced by envelope stress
Given that the in vivo transcriptome profile of persistent Y. pseudotuberculosis resembles that of bacteria in the stationary phase (11), we investigated whether RfaH protein expression in Y. pseudotuberculosis is dependent on the bacterial growth phase. To do this, we generated an RfaH-LacZ translational fusion construct where the RfaH regulatory region, including the promoter region, ribosome binding site, and the coding sequence of the first 13 amino acids, was fused to the lacZ coding sequence without a start codon and measured the β-galactosidase activity of the construct. As a result, the expression of RfaH increased progressively over time and peaked during the late stationary phase (Fig. 1A), a pattern consistent with that observed in Salmonella enterica serovar Typhimurium (30, 31). This finding provided a starting point for exploring environmental cues that may influence RfaH production. Additionally, under in vitro conditions mimicking the host environments, we investigated the impact of specific environmental factors such as high osmolarity, bile salts, glucose, low pH, oxidative stress, temperature, and a combination of high osmolarity, bile salts, and glucose (BNG) on RfaH expression during both exponential and late stationary growth phases. To determine whether in vivo-relevant stress conditions can influence RfaH expression during exponential growth, when its baseline expression is relatively low, we exposed exponentially growing bacteria (OD₆₀₀ ≈ 0.4–0.5) to various stress conditions for 1 hour. High osmolarity, bile salts, and BNG significantly induced RfaH expression even at this early growth stage, with bile salts and BNG causing particularly dramatic increases (Fig. 1B). In contrast, low pH and 37°C slightly reduced RfaH expression, while glucose supplementation and oxidative stress had negligible effects (Fig. 1B). To test whether these conditions could further enhance RfaH levels during a phase when the expression is already elevated, we repeated the experiments in stationary-phase cultures. All tested conditions led to significant increases in RfaH expression, with bile salts and BNG again producing the most pronounced induction (Fig. 1C). Notably, unlike in the exponential phase, RfaH expression was not reduced at 37°C in stationary-phase cells. Thus, the host-associated stress conditions stimulate RfaH expression during both exponential and stationary growth phases, indicating that this regulatory protein may play a role in the pathogen’s adaptability to hostile environments in the host. The dramatic increase of RfaH expression under high osmolarity, bile salts, and BNG combinations, which are known to induce envelope stress (32, 33), suggests that RfaH expression might be induced by envelope stress.
*RfaH expressions increased during the stationary phase and under infection-relevant stress conditions. (A) The β-galactosidase activity of the RfaH and LacZ translational fusion construct reflected in the hourly increase in optical density at 600 nm (OD600) during bacterial growth at 26°C. (B and C) The β-galactosidase activity of RfaH and LacZ translational fusion construct in bacterial cultures at exponential phase (B) and stationary phase (C), after treatment with infection-relevant stressors: 125 mM NaCl, 0.5% bile salts, 0.4% glucose, pH 6, 10 mM H2O2, and a combination of 125 mM NaCl, 0.5% bile salts, and 0.4% glucose (BNG). Six independent biological replicates were used in panels A, B, and C. In panel A, dots represent the mean values of the six biological replicates, with error bars indicating standard deviations. In panels B and C, the bars represent the mean of six biological replicates (also shown as individual dots). Statistical significance was determined using an unpaired t-test, with Luria broth medium as the control. Asterisks indicate P-values as *<0.05, **<0.01, ***<0.0005, and ***<0.0001 and ns (not significant) as >0.05.
Lack of RfaH is associated with bacterial aggregation and reduced motility
To investigate the impact of rfaH deletion, we performed phenotypic assays and assessed bacterial morphology using both light and atomic force microscopy. Assays were conducted under conditions with high RfaH expression, as well as under a control condition at 26°C. The ΔrfaH strain aggregated after being subjected to NaCl, BNG, and temperature (37°C for 2 hours), suggesting that RfaH influences bacterial surface structures (Fig. 2A). However, the expression of rfaH in trans complemented the aggregation phenotype (Fig. 2A). To further assess motility, we performed soft agar motility assays at 26°C using both wt and ΔrfaH strains, following exposure to NaCl and BNG. This temperature was chosen since Y. pseudotuberculosis is non-motile at 37°C in vitro, and previous transcriptomic profiling showed that specifically during persistent infection, flagellar gene expression partially resumes and the global transcriptional profile resembles that of bacteria grown at 26°C (11). Under high osmolarity and BNG treatment, the ΔrfaH strain displayed reduced motility, and this defect was rescued by complementation of rfaH in trans (Fig. 2B). To determine whether the motility defects were due to structural changes in flagella, we visualized both untreated and stress-treated bacterial cells using atomic force microscopy. Despite the loss of motility, the ΔrfaH strain retained flagella in both NaCl and BNG-supplemented media (Fig. 2C). These findings suggest that the reduced motility is not due to flagella absence but may be due to the involvement of RfaH in regulating surface structures, potentially through its influence on cell aggregation or chemotaxis signaling pathways.
Loss of rfaH reduces motility despite the presence of flagella under stress conditions. (A) Phase-contrast microscopy images of wt, ΔrfaH, and ΔrfaH complemented with in trans rfaH expression (prfaH) strains under various conditions: control (26°C), 125 mM NaCl, a combination of 125 mM NaCl, 0.5% bile salts, and 0.4% glucose (BNG), and at 37°C. (B) Motility profile of wt, ΔrfaH, and prfaH strains under control, 125 mM NaCl, and BNG conditions at 26°C. (C) Atomic force microscopy images of the flagella from the wt and ΔrfaH strains under control, 125 mM NaCl, and BNG conditions at 26°C.
Defective O-antigen biosynthesis contributes to RfaH-dependent phenotypic changes
Given that RfaH is known to regulate LPS in Y. pseudotuberculosis (28), we hypothesized that the aggregation observed in the ΔrfaH mutant strain might be due to defective LPS. A likely explanation is that shortened LPS exposes underlying outer membrane adhesins such as autotransporters like Ag43 that are normally masked by intact LPS. Exposure to these adhesins has been shown to enhance bacterial auto-aggregation in other LPS-deficient strains (34–36). LPS comprises three distinct regions: lipid A, a core polysaccharide, and an O-antigen (Fig. 3A). Silver-stained SDS-PAGE analysis of LPS profiles from wt and ΔrfaH strains revealed that the ΔrfaH strain lacks a single O-antigen unit of outer core LPS, consisting of lipid A, core, and one O-repeat unit, at both 26°C and 37°C (Fig. 3B). The lower molecular weight of the outer core in the ΔrfaH strain was previously observed in Y. pseudotuberculosis IP32953 and Y. enterocolitica and was associated with the absence of the single O-antigen unit (28, 37). Further investigation into the composition of lipid A species in both wt and ΔrfaH strains using matrix-assisted laser desorption/ionization – time of flight (MALDI-TOF) analysis revealed no differences between the two strains at either temperature (Fig. 3C and D). This suggests that the lower molecular weight of the LPS produced by the ΔrfaH strain may result from insufficient O-antigen biosynthesis, as the operon responsible for O-antigen biosynthesis, which is known to be regulated by RfaH, has an ops sequence located in the upstream region of the operon (Fig. 3E).
RfaH-related phenotypic changes are indirectly influenced by defective O-antigen biosynthesis. (A) Composition of LPS in Y. pseudotuberculosis. (B) LPS profiles of wt and ΔrfaH mutant strains were analyzed at 26°C and 37°C via 15% SDS-PAGE with silver staining. (C and D) Representative MALDI-TOF mass spectra of lipid-A from wt and ΔrfaH (C) at 26°C and (D) at 37°C. (E) Schematic of the O-antigen biosynthesis operon, highlighting the ops sequence located upstream. Nucleotide substitutions introduced to generate the m-ops mutant are marked in red. The array of seven genes on the distal part of the operon is shown for the ΔO-aB deletion mutant. (F) LPS profiles of wt, ΔrfaH, and other O-antigen biosynthesis operon mutants and the ΔrfaH complementation strain were analyzed at 26°C via 15% SDS-PAGE with silver staining. (G) Phase-contrast microscopy images of multiple mutants with defective O-antigen biosynthesis.
To determine whether the ΔrfaH phenotypes are due to defective O-antigen biosynthesis, we constructed several mutant strains deficient in O-antigen biosynthesis. To do this, we deleted the gene encoding for CDP-6-deoxy-delta 3,4-glucoseen reductase (ΔddhD), an enzyme essential for CDP-ascarylose production during the biosynthesis of O-antigens (38). Moreover, we deleted a sizable DNA region, including seven genes (gmd, fcI, manC, wbyQ, gne, manB, and wzz), toward the end of the O-antigen operon to severely attenuate O-antigen biosynthesis (called ΔO-aB) (Fig. 3E). Additionally, we generated a mutant strain with a modified ops sequence located upstream of the O-antigen biosynthesis operon by introducing six nucleotide substitutions inside and in the flanking regions of the ops sequence (called m-ops) (Fig. 3E). LPS profiling showed that all three mutant strains exhibited a lower molecular weight of LPS outer core profiles similar to that of the ΔrfaH strain (Fig. 3F). The defective LPS profile of the m-ops strain provides evidence that RfaH positively regulates the O-antigen biosynthesis operon. Microscopy analysis of these O-antigen mutants under NaCl, BNG, and 37°C conditions revealed phenotypes mirroring those of the ΔrfaH strain. Following BNG treatment, these mutants exhibited similar aggregation phenotypes (Fig. 3G). Thus, the aggregation observed in the ΔrfaH strain is likely due to indirect effects from defective O-antigen biosynthesis.
The global effect of RfaH on the Y. pseudotuberculosis transcriptome
To investigate other potential genes or operons regulated by RfaH, we performed RNA-seq on ΔrfaH and wt strains grown at 26°C and 37°C. To distinguish genes whose expression regulation is affected specifically by defective O-antigen biosynthesis in the absence of RfaH, we included m-ops and ΔO-aB strains, both of which have defective O-antigen synthesis. Differential expression analysis revealed consistent and significant downregulation of all genes in the O-antigen biosynthesis operon (e.g., manB, gne, fcl, gmd, wzx, wzy) in both ΔrfaH and m-ops mutants compared to wt at both temperatures (Fig. 4A), consistent with the known role of RfaH in promoting efficient transcription elongation of long operons. In contrast, the ΔO-aB strain showed reduced expression only of the seven deleted genes (Fig. 4A) within the O-antigen operon, confirming the structural basis of the disruption.
Differential gene expression profile of ΔrfaH, m-ops, ΔO-aB mutants, and wild-type strains with validation of the gene expression results by quantitative real-time PCR (qRT-PCR). (A) Heatmap highlighting low expression profiles of O-antigen biosynthesis operon genes in ΔrfaH, m-ops, and ΔO-aB versus the wt strain at 26°C and 37°C. (B and C) Venn diagrams highlighting differential gene expression patterns in ΔrfaH, m-ops, and ΔO-aB versus the wt strain at (B) 26°C and (C) 37°C. (D) Bar plot comparing RNA-seq and qRT-PCR expression levels of six randomly selected genes. The x-axis represents the genes, and the y-axis represents the log2 fold change from both RNA-seq and qRT-PCR. (E) Linear regression analysis of expression level between RNA-seq and qRT-PCR data. The x-axis represents the log2 fold change of RNA-seq, and the y-axis indicates the log2 fold change of qRT-PCR.
Notably, analysis of uniquely differentially expressed genes (DEGs) revealed that ΔrfaH regulated a distinct set of genes not altered in the m-ops or ΔO-aB mutants. At 26°C, eight DEGs were exclusively regulated in ΔrfaH (Fig. 4B), including downregulated genes encoding a thiosulfate transporter (cysP), ribosomal protein (rpmH), a putative adhesin/hemolysin, and flagellar rod protein (flgF) (Table 1), suggesting additional roles for RfaH in motility, translation, and membrane physiology beyond O-antigen regulation. In contrast, genes encoding a putative lipoprotein (ynbE) and a virulence plasmid located transposase (piBX002) were upregulated, possibly due to the loss of RfaH-mediated transcriptional repression of stress-related genes or mobile elements (Table 1). At 37°C, the transcriptomic impact of ΔrfaH was even broader, with 23 ΔrfaH-specific DEGs (Fig. 4C). These included downregulated genes involved in membrane transport (e.g., holin, monosaccharide-transporting ATPase) and phage-related or tail-associated proteins, along with upregulated genes linked to phage activity (e.g., alpA), fimbriae, and type VI secretion system components. The increased number of ΔrfaH-specific DEGs at host temperature suggests additional regulatory roles for RfaH under infection-relevant conditions, potentially modulating envelope structure, mobility, and stress adaptation (Table 1). Notably, no *ops-*like motifs were detected in the regulatory part of any of these genes or operons.
For the m-ops mutant, 25 DEGs were exclusively regulated at 26°C, including strong downregulation of the urease operon (ureC, ureD, ureE, ureF, ureG), which remained unaffected in both ΔrfaH and ΔO-aB mutants (Table 1). These findings suggest that the 5′ untranslated region of the O-antigen biosynthesis operon, including the ops sequence, a known RfaH recruitment signal, may influence additional regulatory pathways beyond O-antigen biosynthesis. In the ΔO-aB strain, 35 exclusive DEGs (Table 1) were identified across both temperatures, consistent with the complete removal of key O-antigen biosynthetic genes. These changes likely reflect secondary responses to the total loss of LPS side chain structure, potentially involving envelope stress, metabolic shifts, or activation of mobile genetic elements. Eight DEGs were shared by all three mutants and corresponded to core O-antigen biosynthesis genes, all consistently downregulated relative to wt, highlighting their dependency on both structural and transcriptional integrity of the operon.
To validate the RNA-seq findings, we performed quantitative real-time PCR (qRT-PCR) on six representative genes. Expression patterns were consistent between qRT-PCR and RNA-seq data, with a strong correlation (R² = 0.8807), thereby confirming the robustness of our transcriptomic analysis (Fig. 4D and E).
RfaH is required for full in vivo virulence of Y. pseudotuberculosis
To assess the role of RfaH in Y. pseudotuberculosis virulence in a mouse model, we performed low-dose oral infection in FVB/N mice using the ΔrfaH mutant and wt strains. Thereafter, we followed Y. pseudotuberculosis colonization with the presence or absence of a bioluminescent signal produced by a bioluminescent reporter system introduced in the virulence plasmid and monitored with In Vivo Imaging System (IVIS).
While up to 94% of the mice were infected with the wt strain, only 19% of the mice were infected with the ΔrfaH mutant strain (Fig. S1A). Furthermore, 36% of wt-infected mice developed acute infection and succumbed to the infection by 16 days post-infection (d.p.i.), whereas none of the ΔrfaH-infected mice showed symptoms of acute infection (Fig. S1A). Interestingly, one ΔrfaH-infected mouse developed symptoms of acute infection, despite the absence of or undetectable bioluminescent signal. While this suggests the possibility of transient colonization or IVIS sensitivity limitations, we cannot rule out unrelated causes of illness in this individual animal. Moreover, the ability of the ΔrfaH strain to establish persistent infection was reduced, with only two mice maintaining infection, compared to five mice infected with the wild-type strain up to 30 d.p.i. (Fig. S1A). This reduction is likely influenced by the requirement for RfaH during the initial stages of infection, with a notable number of mice remaining uninfected. Notably, the attenuation observed in the ΔrfaH mutant in the mouse oral infection model could be complemented with the expression of rfaH in trans (Fig. S1A). Infection with the prfaH complementation strain resulted in four mice developing persistent infection, indicating that RfaH is required to establish persistent infection in mice. Additionally, we monitored the presence of prfaH during infection by continually screening for antibiotic resistance, encoded on the plasmid that encodes rfaH, on Y. pseudotuberculosis that was shed through feces (Fig. S1B). Our results confirmed that prfaH was still present even after 31 d.p.i., and with a high bacterial load after 10 d.p.i. (Fig. S1B). Overall, the infection profile of ΔrfaH-infected mice showed attenuation in virulence capacity at both the early (7 d.p.i.) and persistent (30–31 d.p.i.) phases, whereas this attenuation was complemented in mice infected with the prfaH strain (Fig. 5A). Moreover, these differences were statistically significant at all monitored time points (Fig. 5B). Further analysis of the survival of mice infected with different strains showed significantly higher survival of mice infected with ΔrfaH compared to mice infected with strains expressing this regulator (Fig. 5C).
Loss of RfaH attenuates virulence in low-dose oral infection of FVB/N mice and can be complemented by trans expression of rfaH. (A) Infection profile of FVB/N mice infected with a low dose of wt (n = 17), ΔrfaH (n = 16), and prfaH (n = 16) Y. pseudotuberculosis at early (7 d.p.i.) and persistent (30–31 d.p.i.) infection phases. Mice were orally infected with overnight bacterial cultures, and the infection process was monitored daily post-infection for wt, ΔrfaH mutant, and prfaH strains. (B) Heatmap showing differences (by P-values) in the profiles of mice that were infected, not detected, or succumbed, comparing wt versus ΔrfaH and prfaH strains separately at different time points. P-values were calculated using Fisher’s exact test. (C) Survival rates of mice infected with wt, ΔrfaH, and prfaH strains. Statistical significance was determined using the log-rank (Mantel-Cox) test.
DISCUSSION
In the present study, we found that the RfaH expression reaches the highest level at the stationary phase under various infection-related in vitro conditions. This correlates with the observed upregulation of RfaH during persistent infection, where the transcriptome of persistently colonized Y. pseudotuberculosis is more similar to bacteria in the stationary phase than to that in growing bacteria (11). Similarly, Rojas et al. (30) reported that the expression of RfaH increased up to twofold when the growth of Salmonella reached the late exponential phase and remained high throughout the stationary phase. Moreover, we also observed RfaH upregulation under multiple stress conditions, including NaCl, bile salts, low pH, oxidative stress, temperature, and BNG, implying a possible role of RfaH in stress adaptation and survival in harsh conditions. However, the rfaH-lacZ translational fusion utilized in this study reports regulation up to translation initiation but does not account for potential post-initiation effects such as differential protein stability or regulation during elongation.
Another signature of bacteria in the persistent infection stage was the upregulation of genes involved in envelope perturbation (11). In this study, LPS profiling of strains mutated in different RfaH-dependent features highlighted uniformly defective LPS profiles, similar to the ΔrfaH strain. These observations are consistent with RfaH acting as a positive regulator of the O-antigen biosynthesis operon. Defective LPS biosynthesis impacted multiple bacterial surface processes, including clumping phenotypes of the various mutant strains, which were comparable to the ΔrfaH strains. In Salmonella, RfaH-regulated LPS has been reported to play a key role in the interaction between cells and the environment, contributing to virulence (30). Similarly, regulation of LPS has been reported as a common strategy for bacteria to change their surface and adapt to new environments, such as increased temperature and exposure to different chemicals (39). These results suggest that LPS-defective bacteria will have challenges adapting during infection. Previous studies in E. coli have shown that intestinal colonization was impacted by the regulatory action of RfaH on LPS-core production (40). Surprisingly, the LPS defect might also be responsible for the aggregation phenotype of ΔrfaH, as shown in this study. The reason for this is unclear, but it might be clustering per se or that the LPS defect affects the organization of membrane components in a way that hinders flagella function or the reduced expression of the flgF gene, which encodes the basal body rod protein that transmits motor torque from the hook to the filament, facilitating bacterial movement (41). In Aeromonas hydrophila, Qin et al. (42) reported that FlgF substituted FlgC in the formation of bacterial flagellar rod while preserving the presence of flagella. However, disruption of the flgC led to a 42% reduction of bacterial motility, chemotaxis, and adhesion (42). Consistent with these reports, downregulation of flgF in the ΔrfaH mutant at 26°C may contribute to reduced motility or adhesion, even though flagella are still present (Fig. 2C). Previous studies have reported that deletion of the rfaH gene attenuates bacterial virulence in several ways, including reduction of capsules and intact LPS, hemin receptor, and alpha-hemolysin during infection (29). The E. coli A192PP ΔrfaH mutant was highly susceptible, with no colonies detected after 30 min of incubation in 22% normal human serum or heat-inactivated serum (56°C for 30 min) (43). In this study, our transcriptome analysis highlighted that deletion of rfaH exclusively affected the expression of a specific set of genes at 26°C and 37°C, compared to the wt strain (Table 1). These genes might shed light on the global regulatory function of RfaH in Y. pseudotuberculosis. Genes encoding cytoplasmic protein, flgF, and hemolysin were downregulated in the ΔrfaH strain compared to the wt strain at 26°C. Similarly, genes encoding holin and monosaccharide-transporting ATPase were downregulated at 37°C (Table 1). In E. coli, the alpha-hemolysin expression has been reported to be under the control of RfaH (44). A study on Y. enterocolitica reported a decrease in the amount of Mg^2+^ transport ATPase proteins and phosphate ABC transporter in the YeO3-ΔrfaH strain (37). As suggested by Nagy et al. (23), this might be an adjustment in response to decreased LPS production due to the RfaH deletion, resulting in reduced demand for sugars and energy. In the Y. pseudotuberculosis ΔrfaH strain, the expression of genes encoding lipoprotein and transposase was enhanced at 26°C, while those encoding type VI secretion protein, transcriptional regulator AlpA, type 11 methyltransferase, and fimbrial protein were upregulated at 37°C (Table 1). A previous study of Acinetobacter baumannii reported that an LPS-deficient strain displayed increased expression of genes encoding cell envelope and membrane biogenesis, like lipoproteins and lipoprotein transport system (45). It is therefore possible that alteration of the lipoprotein content of the outer membrane is a response to LPS loss. Fimbrial proteins and type II methyltransferases have been reported to play key roles in the virulence and defense of several bacteria, including E. coli (46), Salmonella (47), Avibacterium paragallinarum (48), Metamycoplasma hominis (49), and Xanthomonas axonopodis pv. Glycines (50). Similarly, an AlpA positively regulated a programmed cell death pathway linked to the virulence of Pseudomonas aeruginosa (51), while the type VI secretion system has been identified within pathogenicity islands of Salmonella (52), as well as in the survival of Vibrio anguillarum and V. cholerae under stress conditions (53, 54). Interestingly, the high number of hypothetical genes among those regulated by RfaH suggests that much remains unknown about this transcriptional regulator. However, further analysis of these genes could provide valuable insights into potential mechanisms of rfaH regulation in infection. Notably, the stronger transcriptional impact observed at 37°C suggests that RfaH plays a more extensive regulatory role under host-like conditions. In contrast, gene expression changes at 26°C likely reflect environmental adaptation or priming for host colonization, consistent with the observed similarity between 26°C-grown bacteria and the transcriptome of persistent-phase Y. pseudotuberculosis.
The loss of RfaH severely attenuated the virulence of Y. pseudotuberculosis upon infection, as only two mice infected with the ΔrfaH strain could keep the infection up to 30 d.p.i. as compared to five mice infected with the wt strain (Fig. S1A). These highlight the significant role of RfaH in both the initial phase of infection and in the adaptation and maintenance of the pathogen at later stages. Notably, O-antigen-defective strains of Y. pseudotuberculosis and Y. enterocolitica showed attenuation in both mouse and rabbit infection models (55–57), indicating that an alteration in the cell surface might contribute to the attenuation observed in the rfaH deletion mutant. Similarly, the inactivation of RfaH dramatically decreased the virulence of the uropathogenic E. coli strain (29). However, a previous study reported that deleting the rfaH gene in Y. pseudotuberculosis did not reduce its virulence in BALB/c mice during acute infection (28). However, that study used a different Y. pseudotuberculosis strain (IP32953) and a different mouse model (BALB/c) for infection. We have previously demonstrated that FVB/N mice are more suitable for the establishment of persistent infection by the YPIII strain than BALB/c mice. One difference between the mouse strains used in the two studies is their major histocompatibility complex class I haplotype: BALB/c mice possess H-2^d^, while FVB/N mice have H-2^q^. The susceptibility of FVB/N mice to persistent murine encephalomyelitis virus infection and the increased resistance conferred by an H-2^b^ transgene highlight the significant role of major histocompatibility complex (MHC) haplotype variation in determining host susceptibility to infectious agents (58). This suggests that different outcomes of rfaH deletions could simply be due to different mouse models used in different infections. Moreover, although Y. pseudotuberculosis YPIII and IP32953 strains have the same O-antigen biosynthesis operon structure and upstream ops sequence, IP32953 is known to be more virulent and exhibit greater colonization capacity in mice than YPIII (59). This suggests that strain-specific factors, beyond rfaH and O-antigen production, significantly influence virulence in mouse models of Y. pseudotuberculosis infection.
Mutations in the ops region can impair RfaH binding, leading to premature transcription termination or improper transcription of downstream genes, thereby reducing their expression. Interestingly, our RNA-seq analysis revealed that the ops mutation affected the transcriptional regulation of several genes that are not regulated by RfaH. This finding underscores a possible distinct regulatory role for this ops element, independent of RfaH, in the expression of specific genes. Notably, analysis of read mappings of the O-antigen biosynthesis operon’s 5′ untranslated region, surrounding the ops region, revealed a unique mapping profile in the m-ops strain. The relatively low coverage of the GGGGGG sequence downstream of the ops sequence in the wt and ΔrfaH strains, compared to the flanking regions, was absent in the m-ops strain (Fig. S2). This observation suggests the presence of a potential RNA motif that may have been disrupted by a single nucleotide substitution in the m-ops strain, possibly indicating the action of an RNase that processes the 5′ UTR of the O-antigen biosynthesis operon. Consequently, a possible cleavage of this region may result in an approximately 136 nt long 5′ UTR, which may function as a regulatory non-coding RNA precisely modulating the expression of genes uniquely regulated in the m-ops strain. In E. coli, the ops sequence leads to the formation of a DNA hairpin in the non-template strand (60). This hairpin is critical for RfaH recruitment, thereby stabilizing the paused complex and enabling subsequent transcriptional activation (61). Mutations within the ops sequence, like in our m-ops strain, might not only disrupt RfaH binding but also destabilize the formation of this hairpin, thereby abolishing RNAP pausing. Thus, the low read coverage downstream of ops sequence in the m-ops strain can be alternatively explained by the loss of this structured pause element, rather than (or in addition to) the action of a hypothetical non-coding RNA. However, all these hypotheses are beyond the scope of the current study, and they require further investigation to elucidate the novel regulatory role of the 5′ UTR in the O-antigen biosynthesis operon.
MATERIALS AND METHODS
Strains and growth conditions
The wt strain in this study was the kanamycin-resistant bioluminescent Y. pseudotuberculosis YPIII/pIBX strain (Table S1). The YPIII strain is a well-studied model for Y. pseudotuberculosis pathogenesis (10, 11). The strains were cultured overnight at 26°C and 220 rpm in Luria broth (LB) supplemented with 50 µg/mL kanamycin. The samples were co-cultured in the morning to a starting OD of 0.05 and grown in LB at 26°C until the exponential phase before being aliquoted for several in vitro growth and total RNA purification experiments at 26 and 37°C. For stress conditions, the LB was supplemented with 125 mM NaCl, 0.5% bile, 0.2% glucose, and a combination of all three (LB-BNG).
Mutant and other strain constructions
To generate an in-frame deletion of a single gene or large genomic region of DNA, a 200-nucleotide flanking region in both ends of the region of interest was PCR amplified and ligated together with SalI and BglI (New England Biolabs, Inc.) into pDM4 (62) using the In-Fusion HD Cloning Kit (Clontech Laboratories, Inc.) according to the manufacturer’s instructions. The same strategy was employed for multiple nucleotide substitutions by using degenerate PCR primers at the PCR amplification step. The plasmids were transformed into E. coli DH5αλpir, and positive clones were selected on chloramphenicol (50 µg/mL) containing agar plates, confirmed by colony PCR. Confirmed plasmid constructs were transformed into E. coli S17-1λpir conjugation strain for conjugal mating with Y. pseudotuberculosis YPIII/pIBX. Positive allelic exchange was selected as described previously (62). The mutant strains of ΔrfaH, m-ops, and ΔO-aB were finally confirmed with sequencing (Table S1). For the complementation of rfaH, the gene and its promoter sequence were cloned into the BamHI and HindIII sites of pMMB66HE, resulting in prfaH. The promoter region of rfaH and a part of the downstream region encoding the first 13 amino acids were fused to promoterless lacZ in pFU61, where ColE1 had been exchanged with the low-copy replicon pSC101.
Mouse infection and bioluminescent imaging
Bacteria were subcultured on LB agar plates supplemented with kanamycin (50 µg/mL). For infection, the bacteria were grown overnight in LB at 26°C, and concentrations were estimated by absorbance at OD_600_. Cultures were resuspended to 10^7^ colony forming units (CFUs)/mL in sterilized tap water supplemented with 150 mM NaCl. Viable count and drinking volume determined the infection dose. Eight-week-old female FVB/N (Taconic Farms, Inc.) mice were deprived of food and water for 16 hours before oral infection with ~10^7^ CFUs of wt Y. pseudotuberculosis YPIII (pIBX, harboring the luxCDBAE operon in the virulence plasmid pYV) or the corresponding ΔrfaH mutant strain, which were supplied in their drinking water for 6 hours. Mice were inspected frequently for signs of infection and to ensure that infected mice showing prominent clinical signs were euthanized promptly to prevent suffering. The infections were monitored up to 31 d.p.i. by an in vivo imaging system, IVIS Spectrum, which was based on detecting the light produced by luciferase activity encoded on the luxCDBAE operon (63). Before imaging, the mice were anesthetized using the XGI-8 gas anesthesia system (Caliper LifeSciences, Inc.), which allowed control throughout anesthesia. The oxygen mixed with 2.5% IsoFloVet (Orion Pharma, Abbott Laboratories Ltd., Great Britain) was used for the initial anesthesia, and 0.5% isoflurane in oxygen was used during imaging.
β-Galactosidase assays
The in vivo transcriptome profile of persistent Y. pseudotuberculosis has been reported to resemble that of stationary-phase bacteria (11). Thus, we created a reporter vector with the rfaH promoter region, ribosome binding site, and first 13 amino acids fused to the lacZ coding sequence gene to screen for induction of RfaH expression by measuring β-galactosidase activity. The reporter vector was expressed in trans in wt Y. pseudotuberculosis, and the β-galactosidase activity was measured during growth in LB at 26°C. The expression level of RfaH was calculated by measuring the β-galactosidase activity of the strain with the RfaH-LacZ fusion. The β-galactosidase activity in this study was measured according to the method described by Zhang et al. (64). β-Galactosidase activity samples were taken every hour for the growth phase-dependent period, and absorbance at OD_600_ was recorded. On the other hand, we measured the RfaH activity after adding 125 mM NaCl, 0.5% bile salts, low pH with adding acetic acid, H_2_O_2_, temperature (37°C), and a combination of bile, NaCl, and glucose (BNG). The treatments were performed on bacterial cultures in LB at exponential (OD_600_: 0.4–0.5) and stationary (OD_600_: 2.2–2.4) phases and lasted for 1 hour, whereafter the activity of β-galactosidase was measured. All the experiments were repeated with six biological replicates, and enzyme activities (Miller units) were normalized according to cell density (OD_600_).
Phenotypic analysis
The wt and ΔrfaH Y. pseudotuberculosis strains were grown as described in the strain and growth conditions section. To visualize bacterial morphologies, the bacterial samples were mounted on slides and visualized under a phase contrast microscope after 2 hours of exposure to stress conditions and when the bacterial culture reached OD_600_ = 0.8.
Motility assay
Bacteria from overnight cultures were inoculated into LB and grown to OD_600_ = 0.8. Subsequently, they were treated with and without NaCl or BNG at 26°C for 2 hours. A 5 µL aliquot of each culture was spotted on LB with 0.25% agar. Plates were incubated at 26°C under aerobic conditions for 48 hours. The images of bacteria on the plates were monitored using a ChemiDoc XRS System (Bio-Rad).
Visualization of flagella by atomic force microscopy
The wt and ΔrfaH of Y. pseudotuberculosis YPIII-pIBX strains were grown overnight, diluted 25 times with LB media in the morning, and then cultured for 2 hours at 26°C to an OD_600_ = 0.8. Immediately after the bacterial cultures reached the desired OD_600_ value, the growth medium was supplemented with 125 mM NaCl or 0.5% bile, 125 mM NaCl, and 0.2% glucose. Following a 2-hour treatment, the samples were centrifuged for 4 min at 1,500 rpm, washed once with 2 mM MgCl_2_, and resuspended in 50 μL–200 μL of the same solution. Ten microliters of each sample was placed on freshly cleaved ruby red mica (Goodfellow Cambridge Ltd., Cambridge), incubated for 5 min at room temperature, and blotted dry before being placed into a desiccator for at least 2 hours. Images were collected by a Nanoscope V atomic force microscope (Bruker software) using ScanAsyst in the air with ScanAsyst cantilevers at a scan rate of approximately 0.9–1 Hz. The resulting images were flattened and/or plane-fitted in both axes using Bruker software and presented in amplitude (error) mode.
LPS analysis
Diluted overnight cultures were grown at 26°C until OD_600_ = 0.8. Thereafter, 1 mL cultures were centrifuged, and pellets were resuspended in 100 µL lysis buffer (100 mM Tris-HCl, pH 6.8, 1.5% SDS, 1.5% β-mercaptoethanol) and boiled for 10 min. After that, samples were subjected to an overnight treatment with 80 µg proteinase K, followed by SDS-PAGE gel electrophoresis. The gels were stained with Pierce Silver Staining Kit (Life Technologies, Inc.) according to the manufacturer’s instructions.
MALDIxin test
The bacterial pellet was resuspended in 200 µL of distilled water, washed three times with double-distilled water, and resuspended in 100 µL of double-distilled water. A 50 µL aliquot was then submitted to mild-acid hydrolysis by adding 50 µL of 2% acetic acid in double-distilled water and heating for 1 hour at 100°C. The hydrolyzed cells were spun, the supernatant was discarded, and the pellet was suspended in 25 µL of double-distilled water. An aliquot of 0.4 µL of the bacterial solution was loaded onto the target and immediately overlaid with 0.8 µL of a super-2,5-dihydroxybenzoic acid matrix (Sigma-Aldrich, Gillingham, UK) used at a final concentration of 10 mg/mL in chloroform/methanol (90:10, vol/vol). The bacterial solution and matrix were mixed directly on the target by pipetting, and the mix was dried gently under a stream of air (<1 min). Using the reflectron mode, MALDI-TOF MS analysis was performed on a 4800 Proteomics Analyzer (Applied Biosystems, Foster City, CA, USA). Samples were analyzed by operating at 20 kV in the negative-ion mode using an extraction delay set at 20 ns. MS data were analyzed using Data Explorer version 4.9 (Applied Biosystems)
RNA extraction, RNA-seq library construction, and Illumina sequencing
Total bacterial RNAs for RNA-seq and qRT-PCR analysis were isolated from bacterial cultures grown as described above. Three biological replicates were used to represent each sample at 26°C (control) and 37°C, resulting in 24 samples in total and six pairwise comparisons. The cultures were homogenized with 0.1 mm-sized glass beads with a Mini-Beadbeater (Biospec Products, Inc.). TRIzol Reagent (Life Technologies, Inc.) was used to isolate total RNA according to the manufacturer’s instructions, followed by DNase I treatment (Roche). The NanoDrop 1000 (NanoDrop Technologies, Wilmington, DE, USA) and Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) were used to assess RNA concentration and integrity, respectively, of the 24 samples. In addition, the integrity of the RNA samples was further tested in agarose gel before being used for subsequent analysis.
For the library preparation, the bacterial mRNAs were enriched by depleting ribosomal RNA using a Ribo-Zero rRNA removal kit (Illumina). Following depletion, 2.5 µg of total RNA from each sample in duplicate was utilized as the initial material for creating cDNA libraries. The strand-specific cDNA libraries of Y. pseudotuberculosis were constructed using the ScriptSeq Complete Bacteria Kit (Illumina) according to the manufacturer’s instructions, while the library concentrations were measured with a Qubit 2.0 Fluorometer (Life Technologies, Inc.). Finally, 10 pmol libraries were used within Illumina Reagent Kit v.3 (150 cycles) and sequenced with the MiSeq System (Illumina, Inc.), and adapters were trimmed by MiSeq internal software.
Reads processing, mapping, and gene expression quantification
In this study, the reference genome of Y. pseudotuberculosis YPIII (NC_010465 for the chromosome and NC_006153 for the pYV plasmid) was used. The CLC-Bio Genomic Workbench (QIAGEN) was then utilized for quality, ambiguity, length trimming, alignment with the reference genome, and normalization of reads per kilobase per million mapped reads. The CLC-Bio Genomic Workbench was used with default parameters for quality, ambiguity, and length trimming. The rRNA and tRNA annotations were deleted from the genome annotation files before mapping to avoid any bias related to rRNA depletion during library preparation. The Q20 percentages were more than 95%, while the Q30 base percentage, which is an indicator of the overall reproducibility and quality of the assay, was greater than 90%. Moreover, the GC contents of all the reads were above 45% for all 24 samples. Differential expression analysis was performed using the DESeq2 R package to identify DEGs between the controls and treated samples. To estimate the expression level, the DESeq2 program was used to normalize the number of counts of each sample gene using the base means. The difference was calculated, and the statistical significance was determined using the negative binomial distribution test (65, 66). Genes with a standard fold change of less than or equal to 1 (≥1 or ≤−1) and a P-value of ≤0.05 between control and treated samples were considered differentially expressed.
Validation of DEGs by qRT-PCR
To validate the reliability and repeatability of the RNA-seq data, six DEGs were randomly selected for verification by qRT-PCR. Notably, the qRT-PCR performed for ureE, wzz, and manB genes was in the m-ops background and compared to the wt at 26°C. Similarly, fliQ and YPK_1228 were also from the m-ops background and compared to the wt, but at 37°C. The qRT-PCR for the yscD gene was in the ΔO-aB background and compared to the wt at 37°C. The bacterial strains were grown, and the total RNA extracted was used as templates for cDNA synthesis with the RevertAid H Minus First Strand cDNA Synthesis Kit (Fermentas). The gene-specific primers (Table S2) were designed using Primer Premier 5.0 software (Premier Biosoft International, Palo Alto, CA, USA). The qRT-PCRs were performed in triplicate for each condition using KAPA SYBR FAST qPCR Master Mix (KAPA Biosystems) and a Real-Time PCR Detection System (Bio-Rad). The stable reference gene YPK_0340 (rpoB) was selected as an internal control to normalize the expression data. The relative expression levels of the six DEGs were calculated according to the 2−ΔΔCT (cycle threshold) method.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Koornhof HJ, Smego Jr. RA, Nicol M. 1999. Yersiniosis II: the pathogenesis of Yersinia infections. Eur J Clin Microbiol Infect Dis 18:87–112. doi:10.1007/s 10096005023710219574 · doi ↗ · pubmed ↗
- 2Martínez-Chavarría LC, Vadyvaloo V. 2015. Yersinia pestis and Yersinia pseudotuberculosis infection: a regulatory RNA perspective. Front Microbiol 6:956. doi:10.3389/fmicb.2015.0095626441890 PMC 4585118 · doi ↗ · pubmed ↗
- 3Smego RA, Frean J, Koornhof HJ. 1999. Yersiniosis I: microbiological and clinicoepidemiological aspects of plague and non-plague Yersinia infections. Eur J Clin Microbiol Infect Dis 18:1–15. doi:10.1007/s 10096005021910192708 · doi ↗ · pubmed ↗
- 4Cornelis GR. 2000. Molecular and cell biology aspects of plague. Proc Natl Acad Sci USA 97:8778–8783. doi:10.1073/pnas.97.16.877810922034 PMC 34011 · doi ↗ · pubmed ↗
- 5Bottone EJ. 1999. Yersinia enterocolitica: overview and epidemiologic correlates. Microbes Infect 1:323–333. doi:10.1016/s 1286-4579(99)80028-810602666 · doi ↗ · pubmed ↗
- 6Paff JR, Triplett DA, Saari TN. 1976. Clinical and laboratory aspects of Yersinia pseudotuberculosis infections, with a report of two cases. Am J Clin Pathol 66:101–110. doi:10.1093/ajcp/66.1.101779444 · doi ↗ · pubmed ↗
- 7Fisher ML, Castillo C, Mecsas J. 2007. Intranasal inoculation of mice with Yersinia pseudotuberculosis causes a lethal lung infection that is dependent on Yersinia outer proteins and Pho P. Infect Immun 75:429–442. doi:10.1128/IAI.01287-0617074849 PMC 1828392 · doi ↗ · pubmed ↗
- 8Galán JE, Wolf-Watz H. 2006. Protein delivery into eukaryotic cells by type III secretion machines. Nature 444:567–573. doi:10.1038/nature 0527217136086 · doi ↗ · pubmed ↗