Mechanisms of Arsenic Interaction in Bacillus subtilis and Related Species with Biotechnological Potential
Luz I. Valenzuela-García, María Teresa Alarcón-Herrera, Elizabeth Cisneros-Lozano, Mario Pedraza-Reyes, Víctor M. Ayala-García

TL;DR
This paper reviews how Bacillus subtilis and related bacteria resist arsenic toxicity through various molecular mechanisms, with potential applications in bioremediation.
Contribution
The paper provides a detailed analysis of arsenic resistance mechanisms in Bacillus species, focusing on the ars and ase operons and their biotechnological implications.
Findings
Bacillus subtilis uses biosorption, redox transformation, and efflux pumps to resist arsenic.
The ars operon includes genes like ArsC, ArsB, and ArsR, which are co-regulated in response to arsenic.
Environmental Bacillus species show evolutionary adaptations and potential for bioremediation.
Abstract
Arsenic (As) toxicity drives the evolution of resistance mechanisms in environmental microorganisms. Bacteria of the Bacillus genus are frequently identified in isolates from arsenic-contaminated sites, highlighting the importance of understanding the molecular mechanisms related to this bacterial genus. Bacillus subtilis, a soil microorganism and Gram-positive model paradigm, employs multiple strategies to counteract As toxicity, including biosorption, redox transformation, active efflux, and inducible genetic regulation. This review provides a comprehensive analysis of the physiological and molecular mechanisms involved in arsenic response in B. subtilis and related species, focusing on the ars and ase operons. The ars operon, located within the mobile SKIN element, encodes a reductase (ArsC), an Acr3-type efflux pump (ArsB), a carbon–arsenic lyase (ArsI/YqcK), and a transcriptional…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
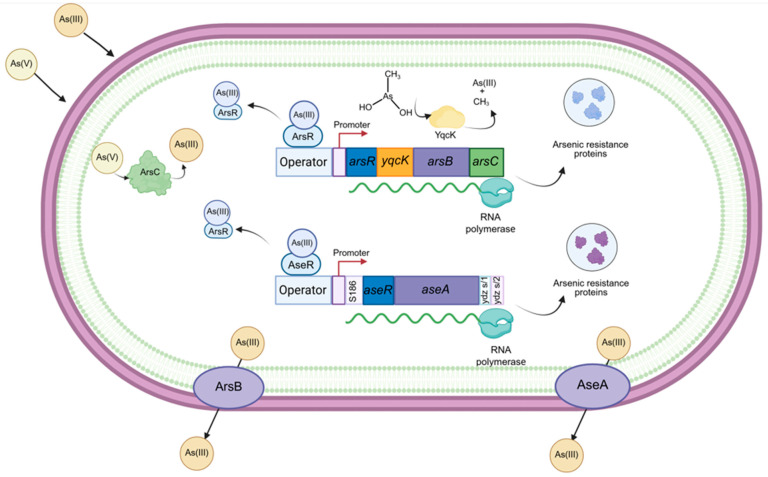 Figure 1
Figure 1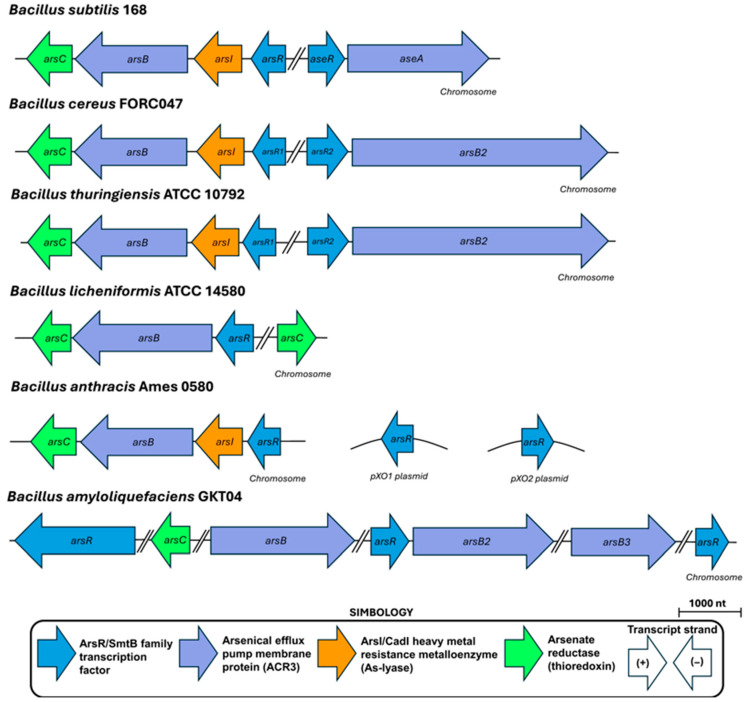 Figure 2
Figure 2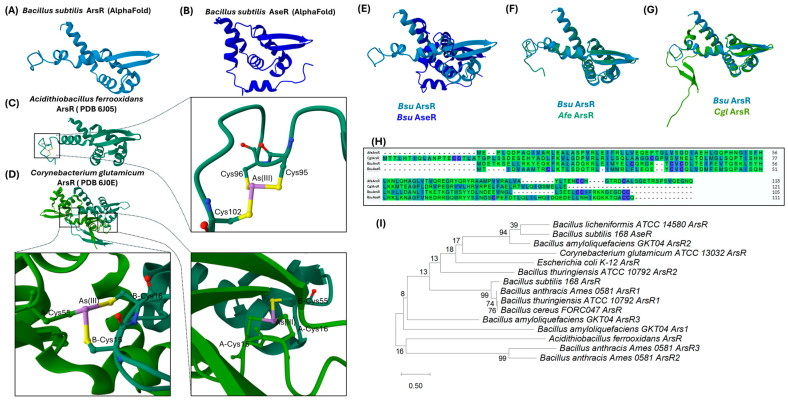 Figure 3
Figure 3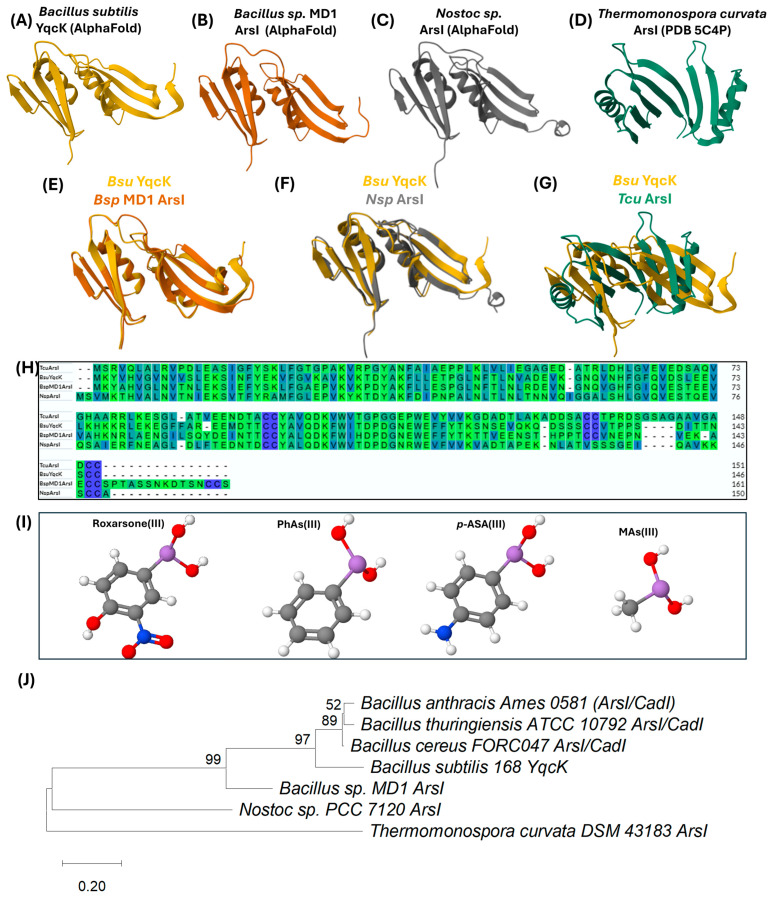 Figure 4
Figure 4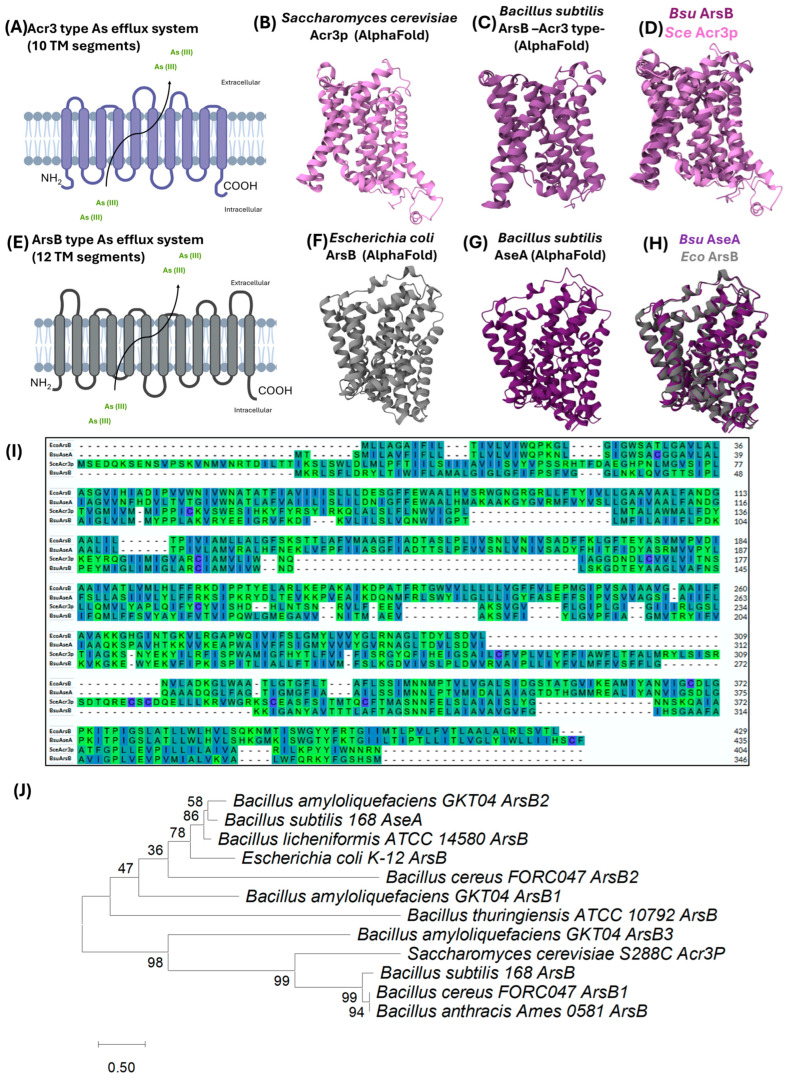 Figure 5
Figure 5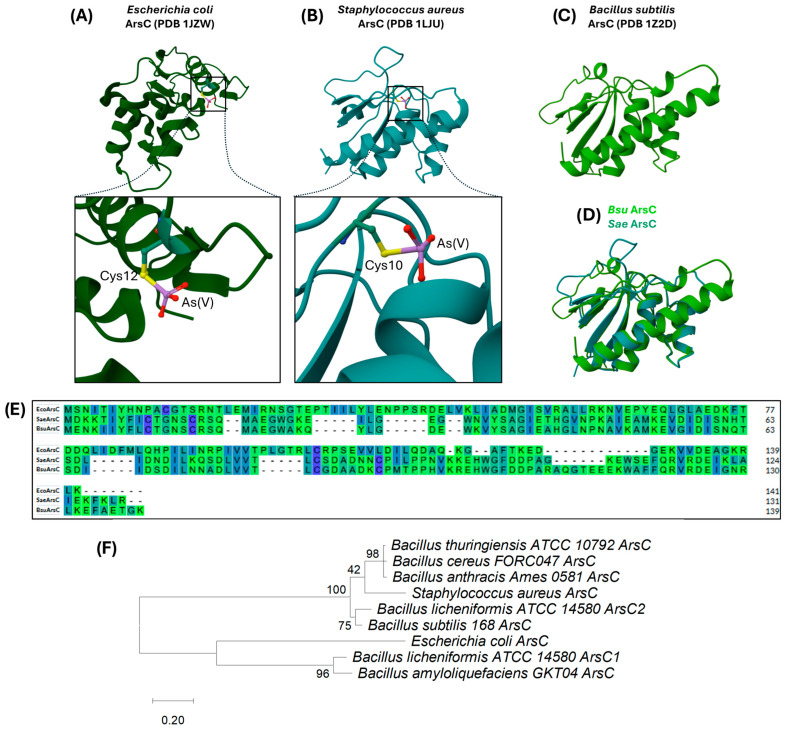 Figure 6
Figure 6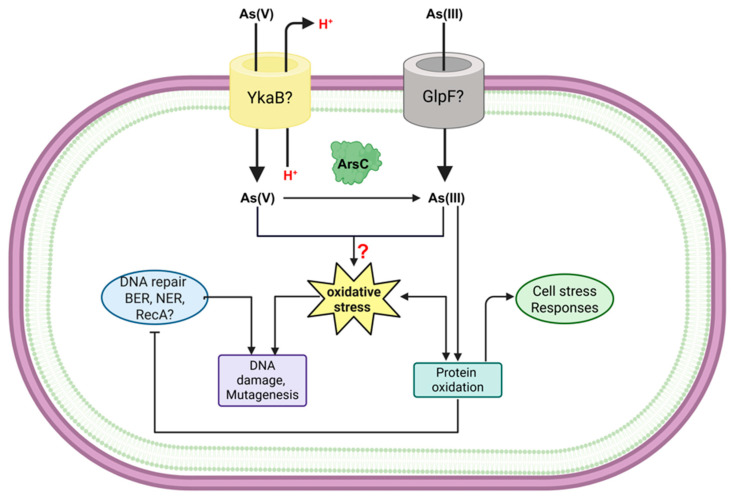 Figure 7
Figure 7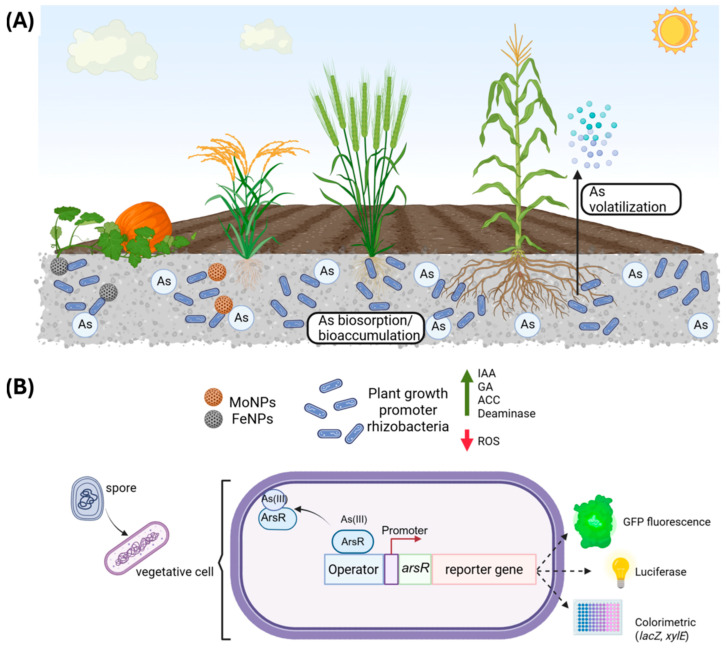 Figure 8
Figure 8| As Resistance Proteins | Possible Genes with Homology in | Similarity (%) | Description | Gene Function Evidence | References |
|---|---|---|---|---|---|
| ArsH |
| 25.41% | NADPH-dependent FMN oxidoreductase | Experimental | [ |
| ArsM |
| 28.70% | similar to N-methyltransferase | Putative | [ |
| ArsN | - | - | [ | ||
| ArsJ | - | - | [ | ||
| ArsP |
| 23.55% | membrane protein similar to permease | Putative | [ |
| ArsO |
| 27.48% | flavin-containing monooxygenase, facilitates cation export via CzcD | Experimental | [ |
| ArsT |
| 26.88% | ferredoxin/flavodoxin-NAD(P) oxidoreductase | Experimental | [ |
| ArsK | - | - | [ |
| Isolation Site Contaminated with Arsenic | Identified Species | References |
|---|---|---|
|
| ||
| West Bengal, India | [ | |
| West Bengal, India | [ | |
| Brahmaputra River basin, India | [ | |
| Groundwater wells of Hazrapara and Ghoshpara locality of Beldanga, Murshidabad (Distt.), West Bengal, India | [ | |
| Well water, Taif City, Kingdom of Saudi Arabia | [ | |
|
| ||
| Spring of water in Qorveh county, Kurdistan province, Iran |
| [ |
| Geothermal systems of Araró, | [ | |
| Surface and groundwater samples, Rautahat District of Nepal | [ | |
|
| ||
| Lake Oliveri–Tindari lake sediments, Italy | [ | |
| Sediments of the Orbetello Lagoon, Italy | [ | |
| Sediments of Mono Lake, California | [ | |
| Sediments of Mono Lake, California | [ | |
| Aquifer sediments of Datong Basin | [ | |
|
| ||
| Soil from Alkaline Crater Lake, Lonar, Maharastra, India |
| [ |
| Soil from Alps, Italy | [ | |
| Agricultural soil and mining origin soil, Guanajuato, Mexico | [ | |
| Soil from Beijing, China |
| [ |
| Soil of the Panki thermal power plant, Kanpur, Uttar Pradesh, India |
| [ |
| Soil of Miyazaki Prefecture, Japan | [ | |
| Soil of Unnao district of Uttar Pradesh (India) | [ | |
| Soil of Uttar Pradesh, India |
| [ |
| Soil of Shanxi Province in Northwest China | [ | |
| Soils from cattle dip sites | [ | |
| Golf course flooring, Florida, United States of America | [ | |
|
| ||
| Industrial effluent treatment plant, Vapi, India | [ | |
| Industrial wastewater from the chemical industry in Sheikhupura, Pakistan |
| [ |
| Wastewater from outskirts of Lahore, Pakistan | [ | |
| Sediment from an effluent drain from a glass-manufacturing plant | [ | |
| Tannery effluents of Savar, Bangladesh |
| [ |
|
| ||
| Sediment from mining site, Hokkaido, Japan | [ | |
| Ore sample, Bundugurang opencast uranium mine, |
| [ |
| Acid mine drainage site in Sabah, Malaysia |
| [ |
| Soil samples from a gold mining area, Paracatu, Minas Gerais, Brazil | [ | |
|
| ||
| Mongolian desert soil |
| [ |
|
| ||
| Rhizosphere of the plant | [ | |
| Rhizosphere of |
| [ |
| Rhizosphere of | [ | |
| Rhizospheric soil samples from the Baruipur district, West Bengal, India |
| [ |
|
| ||
| Surface of used polyethylene | [ | |
| Sludge of a sewage treatment plant, Taiwan, China | [ | |
| Mangrove sediment, Matang Mangrove Forest, Perak, Malaysia | [ |
- —Secretaria de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI)
- —Consejo de Ciencia y Tecnología del Estado de Durango (COCYTED)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsArsenic contamination and mitigation · Chromium effects and bioremediation · Trace Elements in Health
1. Introduction
Bacillus subtilis is a Gram-positive model organism that has gained significant interest in biotechnological applications due to its robust physiological traits, including biofilm formation, spore production and its GRAS (Generally Recognized As Safe) designation that belongs to the Firmicutes phylum [1]. Notably, ~1000 of its 4100 genes (approximately 25%) are orthologues to those from the most extensively studied Gram-negative bacterium Escherichia coli; however, it has been estimated that there are about one billion years of evolutionary divergence between them, leading to notable differences in environmental adaptation [2]. Genes involved in essential functions for bacterial life, such as transcription and translation, metabolic functions, murein biosynthesis and cell division, general stress proteins, and membrane transport are shared by both prokaryotes [2]. Non-ortholog genes in B. subtilis include prophage-related genes and genes for secondary metabolism and antibiotic production, sporulation and competence, transport proteins, and interaction and adaptation to plant environments [3,4,5]. These genetic differences confer B. subtilis’ adaptive characteristics to diverse environments that are of biotechnological interest [2].
The Bacillus genus includes species involved in carbon and nitrogen cycling, human and livestock pathogens such as B. anthracis and B. cereus, and insect pathogens as B. thuringiensis; however, most species within this genus are non-pathogenic [6]. B. subtilis encompasses a set of subspecies and closely related species, collectively referred to as the Bacillus subtilis species complex [6]. Members of this complex share 99% or higher 16S gene sequence identity, making them difficult to distinguish phenotypically based on morphology, physiology, or biochemical characteristics, as these traits may vary significantly among strains. For instance, some B. subtilis strains can grow under anaerobic conditions or at temperatures above 56 °C, while others cannot, potentially leading to confusion with other species within the complex [6]. Species within this complex include Bacillus amyloliquefaciens, Bacillus atrophaeus, Bacillus axarquiensis, Bacillus mojavensis, Bacillus inaquosorum, Bacillus licheniformis, Bacillus malacitensis, Bacillus sonorensis, Bacillus tequilensis, Bacillus vallismortis, and Bacillus velezensis [6].
Among the B. subtilis strains widely used in research are those carrying auxotrophies for threonine (strain 23), nicotinic acid (strain 122), and tryptophan (strains 166, 167, and 168), generated via X-ray mutagenesis. However, the parental wild-type (WT) strain from which they were derived was lost, although all these mutants share the same genetic origin. Of these, strain 168 is the most used as a WT standard reference strain across multiple research groups and is classified as B. subtilis subsp. subtilis, whereas strain W23, which differs both genotypically and phenotypically from strain 168, is referred to as B. subtilis subsp. Spizizenii [7]. Genome sequencing of B. subtilis strain 168 revealed the presence of cryptic prophage elements such as PBSX, SPβ, and phage-like elements such as SKIN, integrated at different chromosomal loci. Therefore, it is feasible that horizontal gene transfer via bacteriophage infection has contributed to the acquisition of specific genes in this strain [2].
Throughout its evolutionary trajectory, Bacillus has developed genetic mechanisms to adapt to various environmental stressors, including the ability to utilize diverse carbon sources, secrete extracellular enzymes, synthesize antibiotics, and thrive in potentially toxic environments, particularly those contaminated with metals and metalloids. Arsenic is a toxic metalloid which has been found to be part of abundant crustal minerals existing for over 3 billion years [8]. To counteract As toxicity, microorganisms have developed efficient detoxification and tolerance mechanisms. This review focuses on the mechanisms employed by bacteria of the Bacillus genus to withstand and proliferate in environments polluted by arsenic, highlighting the main differences with other bacteria less distributed in natural environments such as E. coli.
2. Arsenic Toxicity and Cellular Interactions
2.1. Chemical Forms and Public Health Impact
Arsenic (As) is one of the most naturally abundant elements in the Earth’s crust and oceans, particularly in areas of high hydrothermal activity, mineral deposits, Cenozoic sediments, and carbon sources along the Pacific Ring of Fire [9]. Its ubiquity in soil environments explains the evolutionary selection for microbial genetic elements capable of mitigating its toxic effects, as elevated arsenic levels are detrimental to cellular function [10].
As belongs to Group VA of the periodic table and possesses five valence electrons. Its most common oxidation states are +3 and +5. It can form inorganic compounds with elements such as oxygen, sulfur, iron, and chlorine, as well as organic compounds containing carbon and hydrogen. Both inorganic and organic forms of As can exist predominantly in the trivalent [As(III)] and pentavalent [As(V)] oxidation states, with the trivalent forms generally exhibiting significantly higher toxicity [11,12,13].
2.2. Mechanisms of Toxicity and Detoxification of Arsenic
As toxicity mechanisms vary according to its oxidation state and the biological system in question. As(V), being a molecular analog of phosphate, inhibits oxidative phosphorylation by disrupting energy metabolism. As(III), on the other hand, binds to sulfhydryl groups, impairing protein function and cellular respiration by targeting thiol groups in metabolic enzymes such as pyruvate dehydrogenase and 2-oxoglutarate dehydrogenase. Additionally, As (III) can induce the production of reactive oxygen species (ROS) and cause DNA damage and mutagenesis in mammalian cells [14,15].
Although arsenic is an element that naturally occurs in the Earth’s crust, human activities have led to the release of increasing concentrations of this metalloid to toxic levels in regions and ecosystems where it is not naturally found. The mobility of arsenic, its ability to translocate in plants and bioaccumulate in the food chain, has led to the presence of arsenic in human drinking water and food, increasing exposure doses in humans [16]. Arsenic exposure in humans is associated with health problems such as cancer and cardiovascular, neurological, cardiac, and dermatological disorders [17]. Arsenic detoxification in humans requires enzymatic transformation into the methylated and oxidized forms, monomethyl arsenate [MAs(V)] and dimethyl arsenate [DMAs(V)], which are excreted in the urine. Dimethyl arsenite [DMA(III)] is the precursor of more complex and less toxic organic compounds such as arsenosugars, arsenolipids, and arsenobetaine, which are abundant in aquatic environments but can also be degraded to simpler organic species or inorganic arsenic by microbial action.
Microorganisms have evolved various strategies to cope with As toxicity, including surface adsorption, redox transformation (reduction/oxidation), active efflux, volatilization, and DNA repair mechanisms [18,19,20,21,22]. Certain microorganisms can convert arsenic into various organic and inorganic forms, such as monomethylarsonic acid (MMA), dimethylarsinic acid (DMAA), trimethylarsine (TMA), trimethylarsine fatty alcohols (TMAsFOH), arsenic-containing hydrocarbons (AsHCs), arsenolipids, arsenosugars, and arsenobetaine (AB). Moreover, some microbes can degrade or transform naturally occurring or synthetic arsenical compounds such as phenylarsenicals and roxarsone, leading to the release of inorganic arsenic. Enzymes involved in these transformations include arsenic methyltransferases (ArsM), arsenate reductases (ArsC, ArrAB), arsenite oxidases (AioAB, ArxA), acetyltransferases (ArsN), and arsenic lyases (ArsI), among others [19,23].
While E. coli, the Gram-negative bacterial model to study As interactions, is an enterobacteria found in the intestines of animals, the Gram-positive model B. subtilis is found in soil, water, and plants [3,24]. Consequently, the molecular mechanisms underpinning As resistance may significantly differ between the two genera. Therefore, the following sections are aimed at describing mechanistic aspects regarding the processing of As conducted on B. subtilis and other Firmicutes.
2.3. The First Line of Defense: Extracellular Detoxification
The initial factor contributing to As cytotoxicity in Bacillus lies in its ability to penetrate the cell barriers, composed of both a lipid membrane and the bacterial cell wall. Thus, As can be immobilized at the cell surface by interacting biomolecules, or alternatively, transformed into a less toxic state by extracellular or periplasmic enzymes, thereby preventing its entry into the cytoplasm [25].
Arsenate [As(V)] is ionized at comparable pKa values: 2.2, 7.0, and 11.5 for arsenate and 2.1, 7.2, and 12.7 for phosphate [26]. Due to the molecular similarity between the phosphate group and arsenate, several studies suggest that arsenate may use phosphate transport systems as a route of cellular entry [19,27]. In contrast, arsenite [As(III)] tends to form compounds structurally analogous to hexoses, leading to the hypothesis that its uptake may occur via hexose permeases [21,27,28].
The entry of arsenic into microbial cells is a decisive factor in arsenic toxicity. Upon incorporation into the cell cytoplasm, arsenic can cause DNA damage, interact with the sulfhydryl (-SH) groups of proteins, inhibit the function of enzymes that depend on thiol groups, affect protein folding, and interfere with RNA and protein synthesis [29,30]. Therefore, preventing arsenic from entering cells is the first line of defense in bacteria.
2.3.1. Cell Surface Immobilization
The immobilization of As at the bacterial cell surface involves several processes, including biosorption, surfactant-mediated interaction, and precipitation, all of which serve to prevent arsenic from entering the cell. These mechanisms have been extensively studied due to their potential applications in the bioremediation of As and other heavy metals [31].
Biosorption is a physicochemical phenomenon involving passive interactions with biological materials to bind and concentrate contaminants [32]. During As biosorption, ions interact with cell wall or membrane components such as polysaccharides, proteins, and lipids that possess negatively charged functional groups. This is a passive process, not requiring energy or active cellular metabolism, and can even be carried out by non-viable (dead) cells. The critical factor is the presence of functional groups such as carboxyl, hydroxyl, amino, and phosphonate moieties [33]. Biosorption encompasses various interactions at the cell surface, including ion exchange, electrostatic or covalent complex formation with extracellular polymeric substances (EPSs), and physical adsorption onto the cell envelope.
The capacities of As biosorption have been evaluated in several species of the genus Bacillus. At pH 8, B. subtilis exhibited a biosorption capacity of 5.14 mg/g of biomass for As(III) and 2.35 mg/g of biomass for As(V) [34]. In contrast, B. cereus exhibited a maximum biosorption capacity for As(III) of 32.42 mg/g under optimal conditions, through a spontaneous, endothermic process that followed a Langmuir isotherm model [35]. Another strain of B. cereus isolated from a gold mine showed an even higher biosorption capacity of 153.41 mg/g, with hydroxyl, amide, and amine groups identified as the key functional moieties responsible for As binding [36]. Arsenic biosorption has also been found in the biomass of other B. cereus strains, including SZ2, W2, and MCC3402 [33,37,38]. Additional strains of the genus Bacillus have been tested for As biosorption, including Bacillus megaterium strains H3 and UM-123 [39,40], Bacillus licheniformis strains B3-15, DAS1, and NDSA24R [40,41,42,43], Bacillus thuringiensis strains WS3, A01 [38,44], B. amyloliquefaciens NBRISN13 [45], Bacillus salmalaya 139SI [46], Bacillus pacificus AKS1a [47], and Bacillus pumilus [40]. Overall, the biosorption capacities of these strains range from 0.005 mg/g per biomass in B. pumilus [40] to 153.4 mg/g of biomass in B. cereus SZ2 [36]. Furthermore, surface modification of B. subtilis cells by incubation with ferric iron (Fe[III]), which led to a deposition of ferric hydroxide onto the membrane, have shown to enhance As biosorption capacity by up to 9.51 mg/g for As(III) and 12.89 mg/g for As(V) at pH 8 [34]. Another Bacillus species, B. arsenicus MTCC4380, when immobilized on granular-activated carbon or MnFe_2_O_4_, was able to remove both As(III) and As(V) from wastewater through a spontaneous and exothermic process [48].
In species of the genus Bacillus, it has been reported that the biosorption capacity of arsenic is due to the presence of carboxylate, carboxyl, carbonyl, hydroxyl, amide, and amino functional groups on the surface of the biomass, as well as some glycosylated and/or phosphorylated proteins [21,35,36,47]. In addition, the multiple layers of the spores and their specific compositions are very important in the biosorption capacity of metals and metalloids [36]. A kinetic study using B. subtilis and Proteus mirabilis revealed that the rate-limiting step in As biosorption from contaminated soils is intraparticle diffusion [49].
Another strategy employed by microorganisms to cope with As toxicity is bioprecipitation. This mechanism reduces the concentration of soluble As in the surrounding environment by transforming it into insoluble species, thereby decreasing its ability to permeate biological membranes and its associated toxicity. Certain sulfate-reducing bacteria can precipitate As, generating arsenic sulfide under acidic conditions [50]. This phenomenon was also observed in bacteria isolated from an As-contaminated site, where a microbial consortium predominantly composed of species from the Caloramator, Clostridium, and Bacillus genera was able to remove 65% of soluble As within 150 days of incubation, forming crystalline arsenic sulfide (AsS) [51]. Extracellular polymeric substances (EPSs), such as some saccharides, can also act as reducing agents, and some quinones can act as mediators of electron transfer in the reduction of As(V) to As(III) [52].
2.3.2. Transformation of As by Extracellular Enzymes
The chemical transformation of As by enzymes located in the external surface of the cell membrane also constitutes a mechanism that prevents As from entering the cell. For instance, an arsenite oxidase located in the outer surface of the cell membrane of Alcaligenes faecalis has been shown to catalyze the oxidation of arsenite using cytochrome C as an electron acceptor in the presence of azurin [53]. Additional arsenite oxidases have been identified in some β-Proteobacteria isolated from As-rich environments, playing important roles in the respiratory chain, which allows these organisms to utilize arsenic as an energy source [54].
Some species of the Bacillus genus have been reported to use As compounds as terminal electron acceptors during anaerobic respiration. Notably, B. arsenicoselenatis, an isolate from the arsenic-rich Mono Lake, can reduce arsenate to arsenite under anaerobic conditions, using arsenate as a terminal electron acceptor [55]. Similarly, Bacillus sp. strain IIIJ3-1, isolated from As-contaminated water, displays the ability to reduce As(V) to As(III) under anaerobic conditions, putatively employing the arsenate reductase gene arrA, which will be discussed in subsequent sections [56]. Other species such as B. licheniformis A6 and B. flexus As-12 can transform arsenic by oxidizing As(III) to As(V), and the last can also reduce As(V) to As(III) through a cell-associated enzyme activity, although it is unknown whether this activity is carried out by intracellular enzymes or enzymes anchored to the cell surface [57,58,59].
2.4. Gateways to Toxicity: Cellular Uptake Pathways
Due to its structural analogy to phosphate, in E. coli, arsenate is known to be nonspecifically taken up via phosphate transporters such as the inorganic phosphate [P(i)] transporter (Pit) and phosphate-specific transport (Pst) systems. In contrast, arsenite can enter through aquaglyceroporins such as GlpF, which also mediates glycerol uptake [18,25]. Mutations in the Pit or Pst transporters in E. coli confer resistance to arsenate [60,61,62]. Some bacterial species, including Agrobacterium tumefaciens and Streptomyces, utilize the Pst system for arsenate influx; however, this system exhibits greater specificity for phosphate, thereby reducing arsenate uptake [63,64]. Regarding arsenite, some arsenic-resistant bacteria isolated from contaminated environments exhibit alterations in the GlpF transporter or entirely lack a homologous transporter, as observed in Thiomonas sp. 3As and Herminiimonas arsenicoxydans, leading to reduce As(III) influx [29,65].
In B. subtilis, the Pst system facilitates phosphate uptake but not arsenate, indicating that Pst is not the primary route for arsenate entry in this microorganism [66]. The genome of B. subtilis possesses the pit (ykaB) gene, which encodes a low-affinity phosphate:H^+^ symporter belonging to the Pit family [4]; however, its role in arsenate import remains uncharacterized.
In E. coli, arsenite, in the form of arsenious acid As(OH)3, which mimics the structure of glycerol, can be transported via glycerol facilitators such as GlpF [67]. In B. subtilis, the glpF gene also encodes a glycerol uptake facilitator, and according to a previously unpublished report [68], a GlpF-deficient strain exhibits reduced sensitivity to arsenite, involving this protein in the transport of this ion. Thus, no current study has conclusively identified the arsenic uptake pathway in B. subtilis or other species of the genus.
Multiple reports have provided indirect evidence of As uptake in Bacillus species. For example, B. megaterium and B. pumilus strains isolated from an As-contaminated environment, analytical measurements showed 10% cellular arsenic internalization, with the remaining 90% bound to extracellular surface molecules [69]. Additional indirect evidence of As uptake was observed in B. licheniformis strains DAS-1 and DAS-2, where high phosphate concentrations in the medium (0.75 mM) exhibited a protective effect, and arsenate removal increased under phosphate-limiting conditions, supporting the involvement of phosphate transporters in arsenate uptake [42]. Intriguingly, in Bacillus sp. strain DJ-1, isolated from industrial effluent in Vapi, India, 80% of the accumulated arsenic was found compartmentalized within the cytoplasm. Moreover, whole-cell biosensors developed in B. subtilis have demonstrated induction of the ars operon in response to both As(III) and As(V), further confirming that this metalloid is internalized by the cell [70,71].
3. Molecular Arsenic Resistance Mechanisms in Bacillus subtilis
B. subtilis possesses two chromosomal operons that encode As resistance genes: the ars operon, which contributes approximately 50% of the bacterium’s As resistance, and the ase operon, accounting for about 40% of its tolerance to this metalloid [72,73] (Figure 1). This section provides a detailed description of the characteristics of the genes that comprise these two operons and their relationship with other As-resistance genes in other Bacillus species. It also describes the similarities in amino acid sequences and the structure–function relationships of proteins synthesized according to models generated by experimentation or AlphaFold simulation. Additionally, it includes phylogenetic analyses relating the functional characteristics of each gene product in the ars and ase operons.
3.1. The B. subtilis Ars Operon: A Phage-Derived Genetic Island
The ars operon in B. subtilis is located within a chromosomal region known as the sigK intervening (SKIN) element. This region is named as such because it comprises a 48 kb DNA fragment that disrupts the sigma factor gene sigK, creating two truncated genes: spoIVCB, which encodes the N-terminal region of Sigma K, and spoIIIC, which encodes the C-terminal region. Approximately 3 h after the initiation of sporulation, the site-specific recombinase spoIVCA mediates recombination between 5 bp direct repeats, allowing the reconstitution of an intact sigK gene. This process excises the skin element from the genome as a circular DNA molecule. Interestingly, this 42 kb element contains 60 ORFs, of which 32 encode proteins homologous to those found in the temperate phage ϕ105 and the defective phage PBSX, suggesting that it is a remnant from a phage DNA sequence [74,75,76].
The ars operon is encoded in the chromosomal negative strand (2,657,687 → 2,655,346) (genome version: NC_000964.3) and comprises four genes: arsR, arsI (yqcK), arsB, and arsC. It is inducible by arsenate, arsenite, and antimonite. A 2.4 kb transcript has been detected in the presence of arsenic, indicating that all four genes are co-transcribed in response to the metalloid. The transcription start site is located 34 nucleotides upstream of the arsR start codon. The promoter region includes −10 and −35 elements similar to those recognized by the sigma factor A (σ^A^) [72].
To establish the similarities and genome arrangement of As resistance genes in distinct Bacillus species, the genome databases of B. subtilis, B. cereus, B. thuringiensis, B. licheniformis, B. anthracis, and B. amyloliquefaciens were inspected. The accession numbers for these species can be found in Table S1 in the Supplementary Material of this review. The results of these analyses made it apparent that B. subtilis, B. cereus, and B. thuringiensis share similar composition and arrangements of their ars operons (Figure 2). In contrast, species like B. licheniformis, B. anthracis, and B. amyloliquefaciens possess ars operons that exhibit divergences in number and gene composition, in reference to B. subtilis (Figure 2). B. subtilis possesses two well-defined operons on its chromosome with As response genes: (i) the four-cistron containing ars operon encoded in the negative strand, composed of the transcriptional repressor (arsR), the putative arsenite lyase (arsI), the arsenite efflux pump (arsB), and the arsenate reductase (arsC); and (ii) the ase bicistronic operon, which is encoded in the positive strand and includes a transcriptional repressor (aseR) and an arsenite efflux pump (aseA). Similar genetic arrangements are found in B. cereus and B. thuringiensis. However, B. anthracis contains only the ars operon located in the negative strand. This operon is complemented by a pair of putative transcriptional repressors belonging to the ArsR/SmtB family encoded in the pXO1 and pXO2 plasmids. B. licheniformis has two regions with As response genes. The main operon is similar to the ars operon of B. subtilis and is located on the negative strand; however, it lacks an As-lyase gene and contains a second As(III) reductase on the positive strand. Interestingly, in B. amyloliquefaciens, the As resistance genes are distributed in different regions of its chromosome; furthermore, it possesses three ArsR/SmtB repressors, three As(III) efflux pumps, and one arsenate reductase (Figure 2). Of note, the genes encoded in the ars operon of this bacterium share homology with the elements found in the operons of plasmids R773 and R46 of the Gram-negative bacterium E. coli. Therefore, some Bacillus species exhibit a remarkable diversity in the genetic arrangements and strategies used by Bacillus species to cope with arsenic stress.
3.1.1. ArsR: The Metalloid-Sensing Repressor
In bacteria, the transcriptional regulator protein ArsR, functions as an As(III)-inducible repressor that binds to promoter/operator regions and negatively regulates expression of the entire ars operon [77]. In the absence of As, ars genes are transcribed at basal low levels, required to keep ArsR-dependent repression. Upon contact with arsenite (As[III]), ArsR loses its DNA-binding affinity, allowing RNA polymerase to increase the transcription rates of the ars genes. Structural and functional evidence from Acidithiobacillus ferrooxidans and Corynebacterium glutamicum has revealed that ArsR binds arsenite at a so-called type I binding site, employing three conserved cysteines to coordinate the metalloid [78,79] (Figure 3C,D).
In B. subtilis, the open reading frame of arsR consists of 318 bp and encodes a 105 amino acid-long protein with a predicted molecular mass of 12.1473 kDa. B. subtilis ArsR (BsArsR) shares 44% homology with the As repressor from Staphylococcus and 31–33% homology with those from E. coli encoded on plasmids R773 and R46, respectively [80].
BsArsR belongs to the ArsR/SmtB family of metalloregulatory proteins, which includes the repressor proteins SmtB, ArsR, and CdaC from diverse bacterial origins. The proteins that belong to the ArsR/SmtB family share structural and functional properties, acting as homodimer repressors that control the expression of resistance genes for Zn, Co, Cd, Pb, Bi, Ni, and metalloids like As and Sb [81]. Furthermore, in its active free-of-metalloid form, these repressors bind to Ars box, A:T rich sequence elements, existing in regulatory regions of the ars operons [82]. Upon binding of the metal or metalloid, the repressor protein destabilizes its interaction with DNA, thereby freeing the promoter region and allowing transcription of the genes under the repressor’s control [78,83].
Although B. subtils ArsR has not been crystallized, the predicted tridimensional structure derived from AlphaFold analyses (access ID: AF-P45949-F1-v4) [84,85] (Figure 3A) resembles the 3D structures of repressors from A.ferrooxidans (PDB: 6J05) [86] (Figure 3C) and C. glutamicum (PDB: 6J0E) [86] (Figure 3D), whose 3D structures have been determined through X-ray crystallography, thus evidencing a highly conserved structural similarity (Figure 3F,G). The characteristic structure of ArsR proteins consists of five alpha helices and two beta strands arranged in an α1–α2–α3–α4–β1–β2–α5 fold [86,87,88]. The dimer is formed via a quartet of alpha helices made up of the α2 and α5 helices from each subunit [81]. ArsR proteins contain a helix-turn-helix (HTH) DNA-binding motif, a region that enables the repressor to recognize and bind to the promoter elements of arsenic resistance operons. Regarding the metalloid binding site of ArsR, evidence postulates that it can be located in either the N- or C-terminal domains of each ArsR monomer; and in other cases, the binding site seems to be formed after dimer assembly (Figure 3C,D).
In the ArsR protein encoded in the E. coli plasmid R773, the coordination for As(III) binding relies on the residues Cys32, Cys34, and Cys37 located in the α3 helix of the N-terminal end (Type I site) [83,89]. With respect to A. ferrooxidans, the metalloid binding to ArsR is coordinated by the residues Cys95, Cys96, and Cys102, located in the C-terminal α5 helix (Figure 3C, enlarged) [88]. In contrast, the As-binding site of C. glutamicum’s ArsR is established by residues Cys15 and Cys16 existing in one of the monomers and Cys55 in the partner protein (Figure 3D, enlarged) [86]. In B. subtilis, the repressor ArsR contains two potential binding sites: one in the α3 helix (Cys38, Cys40, and Cys32) and another in the α5 helix (Cys93, Cys94, Cys104, Cys105) [68] (Figure 3H).
To further extend the structural conservation analysis of Bacillus ArsR proteins, a phylogenetic tree was constructed using the maximum likelihood method, with MEGA12 software (v. 12.0.14) set to a bootstrap value of 1000 replicates. This tree compared the amino acid sequences of ArsR proteins encoded by the genes indicated in Figure 2, as well as ArsR proteins from other bacteria. Figure 3I reveals high similarity among the ArsR repressors of B. subtilis, B. anthracis, B. thuringiensis, and B. cereus. These species are placed within the same clade, with a high bootstrap value. In contrast, ArsR from B. subtilis was found to significantly diverge from its counterparts in E. coli and C. glutamicum; and such divergence was even greater with ArsR from A. ferrooxidans. Therefore, despite significant differences, ArsR homologues diverging from typical Bacillus species seem to maintain their ability to bind arsenite and activate an As resistance response.
The expression of arsenic-responsive genes is controlled by a regulatory region composed of two cis-acting elements, a promoter dependent on the σ^A^ transcription factor, and an operator binding site for the ArsR homodimer. The ArsR operator does not directly overlap with the RNA polymerase binding site but is located slightly upstream of this promoter. In E. coli, the repressor dimer binds to an imperfect inverted repeat DNA sequence spanning from nucleotide −64 to −40 relative to the transcription start site of the ars operon [82]. In B. subtilis, the transcription start site of the ars operon was located 34 nucleotides upstream of the arsR start codon, and the putative RNA polymerase binding sites at positions −35 (TTGCAT) and −10 (TATAAT) were also identified in the gene’s promoter region [72]. The ArsR repressor protein binding site is suggested to be an 8 bp inverted repeat located 2 bp upstream of the −35 region [72].
3.1.2. ArsI: The Organoarsenical Lyase
The 441 bp long yqcK gene of B. subtilis, also known as arsI, codes for a protein with arsenic lyase activity (ArsI, from As-induced), capable of breaking the carbon–arsenic (C–As) bonds of organoarsenic compounds. ArsI is composed of 146 amino acids with a predicted molecular mass of ~16.5 kDa and is catalytically active in its monomeric form [75,90]. arsI genes are widely distributed among aerobic bacteria and play a fundamental role in degrading organoarsenical compounds in the environment [91]. ArsI activity directly influences the mobility and toxicity of arsenic in ecosystems [92]. ArsI belongs to a class of non-heme ferrous iron [Fe(II)]-dependent dioxygenases, which use molecular oxygen to catalyze the cleavage of the C–As bond [92]. This activity has been found to be essential for the detoxification of organoarsenical compounds and for B. subtilis’ adaptation to arsenic-contaminated environments, underscoring its role in the biogeochemical cycle of this element [91].
Organoarsenical compounds such as monosodium methylarsonate [MAs(V)] and roxarsone [Rox(V)] have been widely used as herbicides and animal growth promoters. MAs(V) is primarily applied as a post-emergent herbicide for selective weed control. Although considered less toxic than inorganic arsenic species, it can undergo environmental degradation to yield the more toxic As(III) [91,93]. Roxarsone, in turn, is commonly added to poultry and livestock feed due to its anticoccidial activity. It improves efficiency and accelerates growth. However, arsenic-rich manure from treated animals—often used as fertilizer—contains 70–90% water-soluble arsenic, which can leach into soil and water systems [94]. Degradation of these compounds by ArsI yields As(III), which can contaminate crops and drinking water sources [95].
ArsI catalyzes the demethylation of methylarsonite [MAs(III)] to As(III) and the dearomatization of aromatic arsenicals like Rox(III) [91]. Its mechanism involves Fe(II) binding to the active site, followed by interaction with the organoarsenic substrate. Molecular oxygen then binds to Fe(II) and is inserted into the C–As bond, leading to its cleavage and the release of As(III). Structural studies have identified Gln8, His65, and Glu117 in Thermomonospora curvata ArsI (corresponding to His5, His62, and Glu115 in Bacillus ArsI) as critical for metal binding. Additionally, Cys98 and Cys99 (Cys96 and Cys97 in Bacillus) form the substrate-binding site [93] (Figure 3H). A loop-gating mechanism has been proposed, in which a structural loop in ArsI regulates substrate and product access [92].
Mutation of key residues, such as Lys105 in T. curvata, to alanine, arginine, or glutamate leads to a loss of catalytic activity and decreased Fe(II) affinity, suggesting that Lys105 is involved in dioxygen activation [95]. Furthermore, chemical modification of cysteine, histidine, and glutamic/aspartic acid residues inhibits ArsI activity, confirming their catalytic importance. Mutation of Cys96 and Cys97 to serine abolishes substrate binding and enzymatic activity [93]. Tyr38, Gln103, and Lys105 also interact with Fe(II) through water molecules and are essential for catalysis [92].
Isothermal titration calorimetry (ITC) studies have shown that Fe(II) binds to ArsI with a dissociation constant (Kd) of approximately 4 µM. In contrast, As(III), the reaction product, does not bind significantly to ArsI. However, MAs(III) and phenylarsenite [PhAs(III)], a Rox(III) analog, bind the enzyme with Kd values of 0.3 µM and 0.6 nM, respectively, indicating a higher affinity of ArsI for Rox(III). Substrate binding results in the quenching of ArsI’s tryptophan fluorescence, enabling quantification of its affinity for different compounds using intrinsic protein fluorescence quenching techniques [93]. Rox(III) degradation by ArsI produces As(III) and 2-nitrohydroquinone, confirming that the enzyme acts as a dioxygenase that catalyzes the rupture of the C-As bond by incorporating one oxygen atom from O_2_ into the carbon and another into the arsenic [91,93]. A graphical representation of the organoarsenical substrates processed by ArsI from T. curvata is shown in Figure 4H.
Additionally, the cyanobacterium Nostoc sp. has been found to possess an ArsI enzyme (NsArsI) capable of demethylating MAs(III) to As(III). In this organism, NsArsI mainly exists in a trimeric state. Heterologous expression of NsarsI in Escherichia coli confers resistance to MAs(III), suggesting its potential application in environmental bioremediation [96]. A comparative analysis of the primary and tertiary structures (Figure 4E,H) of As-lyases from different bacteria revealed that YqcK from B. subtilis (access ID: AF-P45945-F1-v4) [84,85] (Figure 4A) exhibits a high degree of similarity with the ArsI enzyme from Bacillus sp. MD1 (access ID: AF-A0A059WI14-F1-v4) [84,85] (Figure 4B), an environmental strain isolated from a golf course in Florida (Figure 4E) [91]. Interestingly, high structural similarity was also observed between AlphaFold models of B. subtilis YqcK and Nostoc sp. (access ID: AF-A0A1Z4KSY1-F1-v4) [84,85] (Figure 4C,F) ArsI. In contrast, although the crystal structure of T. curvata ArsI has been used as a reference for predicting other As-lyase topologies [93], it only shows moderate structural homology with B. subtilis YqcK (Figure 4G).
In agreement with the comparative structural study of As-lyases described above, a phylogenetic analysis based on amino acid sequences revealed that YqcK from B. subtilis is more closely related to ArsI enzymes from Bacillus sp. MD1 (84% similarity) and Nostoc sp. (81% Similarity) than to ArsI from T. curvata (41% similarity). The latter is the only crystallized As-lyase recorded in the literature; consequently, the majority of inferences regarding As-lyase functions have been derived this protein. These findings highlight the need for structural studies of YqcK from B. subtilis and other ArsI enzymes from B. anthracis, B. thuringiensis, and B. cereus, as they are all phylogenetically related (Figure 4J). These studies will refine our understanding of the molecular mechanisms of resistance to organoarsenical compounds in the genus Bacillus.
3.1.3. ArsB: The Acr3-Type Efflux Pump
arsB, the third gene found in the ars operon of B. subtilis, is composed of 1041 bp. It encodes a 346 amino acid protein with a predicted molecular mass of ~38.1 kDa [90]; this protein has been involved in the efflux of arsenite from the cell, a highly conserved As resistance mechanism in bacteria. Although referred to as arsB (yqcL) in B. subtilis, this gene differs significantly from the arsB genes from E. coli and S. aureus, as it shares only 24% sequence identity with the latter [72,97]. Indeed, BsArsB is more closely related to membrane proteins that belong to the Arsenical Resistance-3 (ACR3) family, as is depicted in the phylogenetic tree of Figure 5J. Both arsB and acr3 encode membrane-bound proteins involved in arsenic resistance in prokaryotes but differ in transport mechanism, substrate specificity, and phylogenetic distribution [25].
While ArsB is restricted to prokaryotes, Acr3 has a broader distribution, being found in bacteria, archaea, fungi, and even in some plants. The prevalence of arsB and acr3 varies by taxonomic group: acr3 predominates in Proteobacteria, Firmicutes, Actinobacteria, and Asgardarchaeota, while arsB is more common in Alphaproteobacteria, Gammaproteobacteria, and Campylobacterota [25,98]. Of note, the product of the arsB/yqcL gene (ID access: AF-P45946-F1-v4) [84,85] located in the SKIN element of B. subtilis shows greater homology to the Acr3p transporter of Saccharomyces cerevisiae [84,85] (access ID: AF-Q06598-F1-v4) [84,85] than to the E. coli arsB transporter [84,85] (access ID: AF-P0AB93-F1-v4) [84,85] (Figure 5B–D,I,J).
The well-characterized E. coli ArsB transporter has been shown to contain 12 transmembrane helices (Figure 5E) and is coupled to a dimeric ATPase encoded by arsA. In contrast, Acr3-type transporters, despite their widespread presence across all domains of life, have not yet been crystallized [28]. Genetic studies using translational fusions to alkaline phosphatase and green fluorescent protein have shown that the B. subtilis Acr3 protein comprises 10 transmembrane helices, with both the N- and C-termini facing the cytoplasm [97] (Figure 5A).
The Acr3-type transporters are powered by a proton motive force (H^+^ gradient) to export arsenite; therefore, they function as antiporters. Their primary substrate is arsenite (As[III]) (Figure 5A,E), although they can also transport antimonite (Sb[III]) [99].
A recent study elucidated the transport mechanism of these antiporters in E. coli BL21 (ΔarsB) expressing Acr3Bs mutants. To this end, the authors employed AlphaFold2 structural modeling, evolutionary conservation analysis, conserved motif identification via ConSurf and WebLogo, Western blotting, and microscale thermophoresis (MST) to measure arsenite-binding affinity. Results from these analyses revealed that Acr3 antiporters of the BART superfamily adopt a folding pattern typical of the Na^+^/H^+^ antiporter NhaA [100]. The predicted structure of B. subtilis ArsB (Acr3Bs) includes 10 transmembrane helices (TM1–TM10) arranged in two topologically inverted repeats (TM1–TM5 and TM6–TM10). In the central domain, discontinuous helices TM4 and TM9 interact to form an X-shaped structure (Figure 5C). Three conserved motifs have been identified: Motif A (GxARCxA) in the TM4 loop, Motif C (NNFE) in TM9, and Motif B (LNSxxQ) in TM5; these form the active binding pocket. Motive A is located in the N-terminal of Acr3Bs; furthermore, it has been shown that the residues G115, R118, and C119 are required for the efficient transport of arsenite [100]. Residues N292 and E295 in Motif C form hydrogen bonds with arsenite, directly contributing to transport. Motif B may be involved in protein protonation/deprotonation; residues R118 (TM4) and E322 (TM10), in this motif, interact with N292 (TM9) to stabilize the X-helix structure [100].
Notably, although the ars operon can be induced by Sb (III), studies have shown that Acr3Bs expression in E. coli confers only marginal resistance to Sb (III), while it significantly enhances resistance to As (III) [72,100]. Results derived from a comparative amino acid alignment between ArsB from B. subtilis, Acr3p from S. cerevisiae, and ArsB proteins from E. coli and S. aureus show the conserved motifs already mentioned (Figure 5I).
3.1.4. ArsC: The Arsenate Reductase
arsC (yqcM), the fourth gene in the ars operon of B. subtilis, is 420 bp long and encodes an arsenate reductase of 139 amino acids with a molecular weight of 15.4 kDa [90], whose crystal structure has been resolved [101]. Arsenate taken up through phosphate transporters can be converted and detoxified in the cytoplasm initially by ArsC, which catalyzes the reduction of arsenate (As[V]) to arsenite (As[III]), a key step in arsenic detoxification. Once reduced, arsenite can be exported by efflux systems such as ArsB and Acr3 or methylated by enzymes like arsenite-S-adenosylmethionine methyltransferase (ArsM) in certain microorganisms [19,27,102,103].
It has been suggested that bacterial and archaeal arsC gene sequences reflect a common ancient origin for arsenate reductases, possibly emerging in the reducing atmosphere of early Earth. Initially, enzymes detoxifying As(III) may have been critical, while the post-Great Oxidation Event environment, richer in As(V), drove the evolution of arsenate-reducing enzymes [102]. The ArsC reaction mechanism involves the direct binding of arsenate at the enzyme’s active site, where it is reduced to arsenite. In E. coli, ArsC contains a single cysteine residue (Cys-12) in the active site, flanked by an arginine triad (Figure 6A enlargement). In contrast, in S. aureus, the enzyme has three additional cysteine residues, including Cys-10, Cys-15, and Cys-90 (Figure 6B enlargement), indicating structural and mechanistic diversity among bacterial arsenate reductases. Interestingly, the ArsC protein from B. subtilis exhibits significant sequence and structural similarities to the ArsC protein from S. aureus (Figure 6D,E). In fact, a phylogenetic analysis of several ArsC proteins of the Bacillus species, including B. subtilis, B. thuringiensis, B. cereus, B. anthracis, and B. licheniformis ArsC2, showed that they are all phylogenetically related to ArsC from S. aureus, as they are found in the same clade. However, they diverge from the ArsC from E. coli. In fact, E. coli ArsC is related to ArsC1 from B. licheniformis and ArsC from B. amyloliquefaciens, evidencing the genetic diversity among Bacillus species.
3.2. The Ase Operon: A Chromosomal Arsenic Resistance System
B. subtilis possesses an additional arsenic resistance operon located in the genomic region 579,354 (+) → 581,741 (+) (Figure 2), which is distant from the ars operon embedded within the SKIN element and transcribed from the negative strand. The ase operon is composed of two genes: aseR and aseA; and two pseudogenes, ydzS/1, and *ydzS/*2 (Figure 2, pseudogenes omitted). The aseR gene encodes a regulatory protein of the operon (Figure 3B), and aseA encodes an efflux pump for As (III) (Figure 5G), while the function of ydzS/1 and ydzS/2 is currently unknown. This operon lacks an arsenate reductase, leading some authors to suggest that the ase operon could be more ancient in Bacillus, possibly arising during the conditions of a reducing atmosphere. In contrast, the ars operon is believed to have been integrated into the genome of B. subtilis via horizontal gene transfer during evolution into the sigK gene, as the SKIN element exhibits characteristics of a prophage [73,74]. However, it has been demonstrated that the ase operon contributes to the arsenic (III) resistance conferred by the ars operon [73].
3.2.1. AseR: A Second Layer of Regulation
The 336 bp long open reading frame of aseR codes for a 111-amino acid protein with a predicted molecular mass of 12.9 kDa [90]. AseR is a transcriptional repressor belonging to the ArsR-SmtB family, although it shares only approximately 27% homology with ArsR from B. subtilis [73,97] (Figure 3B,E,H). In fact, a structural comparison of the AseR (access ID: AF-P96677-F1-v4) [84,85] and ArsR (access ID: AF-P45949-F1-v4) [84,85] 3D models does not reveal the same degree of similarity as that exhibited between the ArsR model from B. subtilis and its homolog in A. ferrooxidans, which is more closely related to B. subtilis phylogenetically (Figure 3H). Despite the low bootstrap value, the phylogenetic analysis in Figure 3I indicates that AseR is a variant of ArsR from B. subtilis, and therefore, a more recent version of this transcriptional repressor; however, this cannot be unambiguously assessed due to the mobility of the SKIN element. The SKIN element contains the ars operon genes and is known to have originated from an ancestral template phage [74]. While both proteins regulate the expression of As resistance genes in B. subtilis, they differ in specificity and response mechanisms to As.
Like ArsR, AseR forms a homodimer that interacts with DNA through its helix-turn-helix (HTH) domain. Although no crystal structure of AseR is currently available, the DNA-binding domain has been mapped to helices α3 and α4 (Figure 3). The critical amino acids for metalloid binding—Cys33, Cys35, and Asp39—are also located in helix α3, forming a triangular geometry as that described for ArsR [78] (Figure 3). In AseR, arsenite binds to the thiol groups of cysteines in the α3 helix, inducing a conformational change that decreases DNA affinity. While in vitro assays showed that AseR can also bind zinc at the same sites, this does not trigger release from DNA, suggesting that the triangular geometry is key to specificity.
It has been shown that AseR binds to the inverted repeat sequence TATATAACGATTTGCTTATATA located in the operon promoter region of the ase operon. This inverted repeat is a conserved binding site for transcriptional regulators involved in arsenic resistance [78].
3.2.2. AseA: An ArsB-Type Efflux Pump
The aseA gene, 1308 bp long, is the second gene in the ase operon (Figure 2) whose transcription is regulated by the AseR repressor [78]. The protein coded is 435 amino acids long and possesses a predicted molecular mass of 47.1 kDa. AseA functions as an As(III) efflux pump contributing to As(III) but not As(V) resistance as it can only export arsenite outside the cells [73]. Deletion of the aseA gene in a genetic background lacking the ars operon increased B. subtilis sensitivity to arsenite; however, the single deletion of aseA from the WT parental strain had a minimal impact on such resistance due to the presence of the ars operon [73]. In B. subtilis strains lacking the SKIN element, the AseA protein may be solely responsible for arsenic efflux [104].
Evidence derived from comparative structural analyses based on AlphaFold simulations has revealed that AseA from B. subtilis (access ID: AF-A0A164VUH8-F1-v4) [84,85] shares significant similarity with ArsB from E. coli. Indeed, both proteins belong to the ArsB permease family of transporters, which are characterized by containing twelve transmembrane passages (Figure 5B). In contrast, ArsB from B. subtilis has been shown to belong to the ACR3 family; however, proteins from this family exhibit only ten transmembrane passages (Figure 5A). Of note, both AseA and ArsB are broadly distributed across Bacillus species (Figure 5J). A phylogenetic analysis confirmed that AseA from B. subtilis is included in the same clade as ArsB from E. coli and B. licheniformis, as well as ArsB2 from B. amyloliquefaciens. Interestingly, B. amyloliquefaciens, a common biocontrol agent, contains arsB, encoding a third enzyme that facilitates the expulsion of As(III), which is more related to the ArsB permease family (Figure 5J). In conjunction, these observations highlight the diversification of proteins produced by environmental bacteria of the genus Bacillus specialized in the expulsion of arsenite.
3.3. Beyond Core Operons: Additional Genetic Determinants
In addition to the genes described in the ars and ase operons, additional determinants of As resistance have been identified in bacteria of the genus Bacillus, including aioAB, aioXRS, arrAB, aioS, and arsD; however, arsH and arsM detected in other genera are absent from the Bacillus species.
The aioAB genes encode subunits of an arsenite oxidase, which can be found in some As resistance operons in bacteria in conjunction with aioR and aioS genes. The structure of the AioB subunit possesses a Rieske [2Fe-2S] domain that enables electron transfer. This enzyme can convert arsenite into the less toxic arsenate, allowing As(III) to serve as an electron donor for anaerobic respiration. In Bacillus sp. strain IIIJ3-1, the product of the aioB gene has been shown to oxidize arsenic under aerobic conditions with kinetic values of Km = 2.8 mM and Vmax = 0.2 mM h^−1^. However, despite its functional similarity, a phylogenetic analysis revealed a distant relationship of this gene to an oxidase from Listeria monocytogenes [56].
The aioXSRS operon, involved in arsenite oxidation, was detected in the isolate B. firmus L-148 [57]. The cistron aioR, codes for a regulator of the histidine kinase sensor, which is coded by the second cistron, aioS. The latter detects As (III) in the environment, autophosphorylates at a histidine residue, and transfers the phosphate group to AioR, which then binds the operon’s promoter region [57].
The arrAB operon from the microorganism B. selenitireducens has been involved in As resistance. Consistently, arrA and arrB code for the large and small subunits of a respiratory arsenate reductase, respectively. This dimeric enzyme uses arsenate as the final electron acceptor in the respiratory chain under anaerobic conditions, facilitating energy generation. The arrAB genes in B. selenitireducens are part of an operon that also includes arrD, a chaperone protein, and arrC, a membrane subunit involved in anchoring and electron transfer [105].
The arsD gene is commonly found in five-gene ars operons, adjacent to arsA, suggesting a co-evolutionary acquisition. arsD can be located on plasmids or in chromosomes in both bacteria and archaea. Notable examples from non-Bacillus species include E. coli (plasmid R773 arsRDABC operon), Klebsiella oxytoca (plasmid pMH12), Acidiphilium multivorum (plasmid pKW301), Salmonella typhimurium (plasmid R46), Halobacterium sp. NRC-1 (plasmid pNRC100 with arsADRC), and Sinorhizobium sp. As4 [106,107]. In Bacillus, arsD has been identified in some operons, including Bacillus sp. UWC [108], Bacillus sp. CDB3 [109], Bacillus sp. PVR YHB 1-1 [110], B. paralicheniformis ZAP17, and B. altitudinis ZAP62 [111]. ArsD plays a bifunctional role, as a transcriptional repressor and a metallochaperone, for heavy metal regulation and detoxification [112]. Firstly, it acts as a repressor regulating ars operon expression [113]. Furthermore, together with ArsR, ArsD modulates gene expression, where ArsR sets the basal level and ArsD potentiates expression of the ars operon. Of note, deletion of arsD results in the overexpression of arsA, arsB, and arsC [82]. Additionally, evidence has shown that ArsD facilitates the transfer of As(III) to the ArsA ATPase, a subunit of the ArsAB efflux pump. This interaction increases ArsA’s affinity for As(III), thus enhancing resistance to environmental arsenite [106].
Structural analysis revealed that ArsD has neighboring cysteine pairs (Cys12-Cys13, Cys112-Cys113, Cys119-Cys120), which are essential for metal binding. Although Cys12-Cys13 is the only conserved pair across all ArsD homologs, each pair may form a separate metalloid binding site. Residues Cys12, Cys13, and Cys18 are directly involved in the As(III) transfer to ArsA. This interaction occurs at cysteine-rich metalloid-binding sites. In silico models suggest that the α1 helix of ArsD and its metalloid binding site facilitate interaction with ArsA [112]. The transfer of As(III) requires ATP hydrolysis and a transient conformation of ArsA during its catalytic cycle [106].
Another gene found in the ars operons is arsN, which encodes a putative acetyltransferase, though its function is not fully understood. This gene has been identified in bacterial genomes of Shingomonas sp. SKA58, Rhizobium leguminosarum bv. viciae 3841, Rhodobacter sphaeroides ATCC17025, Burkholderia sp. 383, Burkholderia vietnamiensis G4, and Burkholderia multivorans ATCC 17616. Experimental evidence suggests that arsN confers arsenate resistance via a mechanism that requires other As detoxification components like arsC or arsD [114].
In the context of the recognition and degradation of organoarsenical compounds, as mentioned above, B. subtilis possesses the arsI gene (yqcK), encoding a putative carbon–arsenic lyase that could degrade organoarsenicals to inorganic arsenic [91]. However, genes like arsH and arsM, involved in organoarsenical degradation, are absent in this microorganism. ArsH, on the one hand, belongs to the FMN- and NADPH-dependent oxidoreductase superfamily that can operate over organoarsenicals, highlighting its role in redox reactions vital for survival in As-contaminated environments [115]. ArsH oxidizes trivalent organoarsenicals into less toxic pentavalent species. The arsH gene has been found in P. putida and plasmids of Serratia marcescens and Sinorhizobium sp., suggesting broad distribution and evolutionary relevance for adaptation to arsenic-rich environments [115]. Heterologous expression studies showed that introducing arsH from P. putida into E. coli grants resistance to MAs(III), phenylarsenite (PhAs(III)), and Rox(III). Interestingly, the yhdA gene of B. subtilis encodes an NADPH-dependent FMN oxidoreductase that shares 25.4% homology with ArsH (Table 1), and although its involvement in chromate detoxification has been described [116], it remains to be clarified whether it could have a role in the detoxification of organoarsenical species.
On the other hand, ArsM catalyzes the transfer of a methyl group from S-adenosylmethionine (SAM) to arsenite, producing monomethylarsonic acid (MMA), dimethylarsinic acid (DMA), and sometimes trimethylarsine (TMA). This methylation process is key to arsenic biotransformation, impacting both detoxification and environmental mobility of As [21]. ArsM’s catalytic mechanism involves the binding of As(III) and SAM to the protein, followed by thiol and methyl group transfers. Conserved cysteine residues at the active site are essential for As binding and maintaining its reduced state [21]. arsM genes have been identified in distinct bacteria, including Rhodopseudomonas palustris, Pseudomonas spp., Halobacterium sp. NRC-1, Streptomyces sp., Amycolatopsis mediterranei, Sphaerobacter thermophilus, Geobacillus kaustophilus, Rubrivivax benzoatilyticus, and Conexibacter woesei [117]. arsM has not been detected in Bacillus species, though some studies suggest that these bacteria can volatilize As, hinting at alternative enzymes or mechanisms [21,117]. Although no enzymes capable of methylating arsenic (As) have been identified in B. subtilis, genome analysis revealed an open reading frame, ydaC, which encodes a putative methyltransferase that shares nearly 30% homology with ArsM (Table 1). This opens the possibility of further researching the functions of uncharacterized proteins that could be involved in arsenic resistance.
A key question about organoarsenicals is, how do they enter and exit the cell? Membrane proteins such as ArsJ, ArsP, and ArsK have been identified in some bacteria, with roles in exporting organic As compounds, while uptake mechanisms remain less characterized. Aquaglyceroporins and sugar permeases may help import certain organoarsenical species [21]. The ArsJ permease in P. aeruginosa DK2 might export pentavalent organoarsenicals like 1-arseno-3-phosphoglycerate [103,118], while ArsP and ArsK proteins can export trivalent organoarsenicals such as methylarsenite (MAsIII) and roxarsone (RoxIII). arsP genes have been detected in Campylobacter jejuni, Cupriavidus metallidurans, and Shewanella putrefaciens [119]. arsK genes are conserved in Agrobacterium, Rhizobium, Ensifer, Mesorhizobium, Pseudochrobactrum, and Sinorhizobium [103,115,120]. So far, none of these genes (arsP, arsJ, or arsK) have been detected in arsenic resistance operons of Bacillus species, leaving the question of whether specific permeases for organic arsenic compounds exist in this genus open. One possibility is that the ycgR gene of B. subtilis could function as a transporter of organic species of As. Its product is 23% similar to ArsP, and it has been identified as a membrane protein (Table 1). However, its function remains to be characterized experimentally.
Some genes like arsN, arsO, and arsT have been found in As resistance operons, but their functions remain poorly understood. The arsN gene encodes a putative acetyltransferase and has been found in Shingomonas sp. SKA58, Rhizobium leguminosarum bv. viciae 3841, Rhodobacter sphaeroides ATCC17025, Burkholderia sp. 383, Burkholderia vietnamiensis G4, and Burkholderia multivorans ATCC 17616. Experimental evidence suggests that arsN confers arsenate resistance via a mechanism that requires other As detoxification components such as arsC or arsD [114]. The arsO gene encodes a putative flavin-binding monooxygenase, whereas arsT encodes a thioredoxin reductase [103,121]. The arsT gene has been identified in the strain Bacillus sp. CDB3, which contains an eight-gene operon (arsRYCDATorf7orf8) [122]. Although no homolog has been found for arsN in B. subtilis, it is known experimentally that CzcO is involved in cation transport and is 27% similar to ArsO. YumC, meanwhile, has been characterized as a ferredo-xin/flavodoxin-NAD(P) oxidoreductase and bears a modest similarity to ArsT. Table 1 lists Ars resistance homologs from B. subtilis and their putative functions.
3.4. Overcoming Genetic Damage: DNA Repair and Stress Responses
It has been widely reported that As can cause DNA damage through distinct mechanisms, including oxidative damage, single- and double-strand breaks, and interference with repair systems. The type of DNA damage is dependent on the As species [68,105].
As has been reported to inhibit DNA repair in E. coli by affecting one or more steps in the post-replication repair pathway. This is based on the observation of decreased survival of WT or nucleotide excision repair-deficient E. coli cells exposed to UV light and subsequently to arsenite, whereas the addition of arsenite after UV irradiation of a recA-deficient strain did not show any further decrease in survival [126].
DNA repair is an important factor contributing to arsenic resistance in both eucaryotes and prokaryotes. Upregulation of DNA repair-related gene transcripts has been observed in arsenic-resistant bacteria such as Thiomonas sp. and Herminiimonas arsenicoxydans and in microbial communities from arsenic-contaminated environments [105].
In B. subtilis, DNA repair mechanisms and the mutagenic effects of some agents, including metals like chromium, have been studied [127,128]. However, studies on the Bacillus response to arsenic exposure have primarily focused on resistance mechanisms such as efflux, reduction, and oxidation, rather than the involvement of DNA repair enzymes. Only one study has highlighted the importance of superoxide dismutase in counteracting oxidative stress caused by As [129]. This lack of information presents an opportunity for future research into DNA repair systems in Bacillus species (Figure 7).
4. Bacillus Species Resistant to Arsenic: From Environmental Strains to Biotechnological Tools
4.1. Environmental Isolates of Arsenic-Resistant Bacillus
The prevalence of arsenic-resistant Bacillus species is notable in various environments and geographic regions, both contaminated and uncontaminated, suggesting that this bacterial genus possesses well-conserved resistance mechanisms and significant adaptive strategies. A wide variety of Bacillus species have been identified in environmental isolates from different sites, including aquifers, sediments, soil, hot springs, industrial effluents, mines, desert ecosystems, mining tailings, hyperalkaline lakes, the rhizosphere, and in association with plants (Table 2). While several reviews have focused on these isolates and their potential for bioremediation [31,111,130,131], this section focuses on the resistance mechanisms described in these isolates without ignoring the biotechnological potential of the particular species to which these isolates belong.
The mechanisms responsible for As resistance in the isolates listed in Table 2 include detoxification through efflux pumps, reduction, oxidation, and immobilization, all of which are encoded by ars, ase, aiox, and arr operon genes found either on plasmids or on the bacterial chromosome [132]. Reduction of arsenate to arsenite by the arsC gene has been associated with resistance mechanisms in some isolates, such as B. cereus AG27, capable of withstanding arsenate at 40 mM and arsenite at 35 mM. This high resistance was attributed to the presence of a 5.14 Kbp plasmid containing the arsC gene, which encodes an arsenate reductase highly similar to that of B. subtilis [133].
In Bacillus firmus L-148, induction of the arsC gene was also observed in the presence of 10 mM As(III), although arsenic in this isolate was shown to induce the expression of resistance genes at different levels: arsA > arsB > aioS > arsR > arsD > arsC. Additional determinants, including ars mem (arsenical membrane protein), arsC rel (As(V) reductase-related protein), acr-3-2 (As(III) efflux pump), and ars trans (arsenic transporter), were detected in this study [57]. Further induced factors in this isolate included stress response and stage IV sporulation proteins, as well as superoxide dismutase [57].
Reduction of arsenate to arsenite, by arsenate reductases, appears to be a common resistance mechanism among several isolates, as a key step preceding the expulsion of As(III) by efflux pumps. This mechanism has been described in B. licheniformis DAS-2, B. firmus L-148, B. simplex, and B. megaterium, where the presence of the arsB gene was identified [41,42,57,134,135]. Gu et al. 2018 found that some Bacillus sp. isolates carrying the gene for the Acr3 efflux pump exhibited higher arsenic tolerance (minimum inhibitory concentration, MIC, of 42 mM NaAsO_2_) than B. megaterium isolates carrying the arsB efflux pump gene (MIC of 24 mM NaAsO_2_) [136].
In some isolates, like Bacillus sp. IIIJ3-1, in addition to As(III) efflux systems (arsB and acr3), the presence of the arrA gene (coding for a respiratory As(V) reductase) and the aioB gene (coding for an As(III) oxidase) was also detected. This isolate demonstrated arsenic sequestration and cytosolic arsenate reduction capabilities, although no arsC gene was detected, suggesting either an alternative As(V) reduction mechanism or sequence diversification of the arsC gene.
In some Bacillus species, such as B. paralicheniformis ZAP17 and B. altitudinis ZAP62, the arsA and arsD genes have also been identified as part of the arsRBCDA operon. The arsA gene encodes an ATPase that powers the arsenic pump, while the arsD gene encodes a metallochaperone that binds As(III) and delivers it to the ArsA subunit of the efflux pump [111].
An alternative mechanism to counteract the toxic effects of As in some Bacillus isolates involves the production of siderophores or, in the case of As, “arsenophores” that are ligands capable of chelating arsenic salts [137]. None of the Bacillus isolates described in Table 2 were found to possess genes encoding arsenic methyltransferases. However, the Bacillus sp. MD1 isolate, recovered from a golf course, was found to carry the arsI gene, responsible for As demethylation. It is well-known that arsenical herbicides are commonly used on golf courses, so the presence of this enzyme in the isolate may be related to the ability to demethylate As, thereby detoxifying it into its inorganic form [91].
As noted above, the presence of As resistance genes in Bacillus isolates has been reported both on the genome and on plasmids. Bacteria like Bacillus sp. AG24, Bacillus sp. AGM13, B. cereus AG24 [135], and B. cereus AG27 [133] carry these determinants in plasmids, while in most arsenic-resistant Bacillus bacteria, the genes are chromosomally encoded. Table 2 lists different environmental strains of Bacillus and the locations where they were found.
4.2. Harnessing Bacillus for Bioremediation and Sustainable Agriculture
The mechanisms described in this review unveil the strong potential that the microorganism of the Bacillus genus possesses for the development of bioremediation, phytoremediation, and environmental monitoring strategies in arsenic-contaminated sites.
One of the advantages of this genus lies in its ability to generate spores, which are differentiated cell forms with characteristics for enduring adverse environments that are inhospitable to other organisms. Additionally, Bacillus has been documented to promote plant growth through the production of volatile compounds; meanwhile, associations such as the Food and Drug Administration (FDA) have classified bacteria such as B.sutilis, B. licheniformis, B.clausii, and B. coagulans as safe given that they do not produce toxins or are pathogenic, and can therefore be used as probiotics or for enzyme production [164,165,166].
The main uses of Bacillus strains in biotechnology also include immobilizing enzymes on spore surfaces, improving stability and reuse, producing biofuels such as butanediol and ethanol, biological compounds such as vitamins and antibiotics, and even developing vaccines that use B. subtilis as an antigen presenter on the cell or spore surface. All of this highlights the importance of understanding the ability of Bacillus strains that may encounter the metalloid in both industrial and agricultural water to detoxify arsenic and their role in mitigating toxicity in agricultural food production. Some studies have addressed the possibility of using Bacillus strains for As removal in water or soil, and others have addressed their contribution to reducing toxicity to the microenvironment through various interactions with microorganisms or plants.
The biosorption, immobilization, and capture capacities of various Bacillus species, whether through living cells, inactive biomass, or modified biomass, can be applied to immobilize As in soils, remove it from aqueous solutions and effluents, and mitigate toxicity in ecosystems, thereby contributing to the proliferation of other species. A clear example is the non-living biomass of B. cereus Sz2, which has a high As(III) biosorption capacity of 153.41 mg/g and can be reused at a rate of up to 94% after desorption at an acidic pH of 1. This makes it ideal for arsenic removal in mine effluents [36]. Similarly, the B. salmalaya 139SI strain can be used as a biosorbent for As(V) in aqueous solutions, as it reaches a maximum removal efficiency of 92% and a recovery rate of 93% after ten desorption cycles [46]. B. thuringiensis WS3 biomass has also been used to remove As(III) from wastewater via passive adsorption of metal ions through functional groups in the cell wall, such as amino, phosphate, hydroxyl, and carboxylate [38]. A mixture of bacteria, including B. amyloliquefaciens, B. cereus, B. velezensis, and Bacillus sp., which was immobilized with loaded biocarbon, demonstrated the ability to immobilize arsenic, lead, and cadmium in soils through complexation, ion exchange, oxidation, and precipitation mechanisms [167].
The ability of certain Bacillus species to control the mobility of arsenic in soil is applied in environmental cleanup efforts [168]. Moreover, plant growth-promoting rhizobacteria (PGPR) not only lessen metal toxicity in plants but also prevent arsenic from reaching the consumable portions of crops, which is crucial for ensuring food safety [168].
Some strains of this genus, isolated from arsenic-contaminated agricultural soils, including Bacillus sp. ZH16, helped reduce arsenic translocation and promoted wheat plant growth when combined with biogenic molybdenum nanoparticles (MoNPs) while triggering the production of indole-3-acetic acid (IAA), phosphate solubilization, and ACC deaminase activity [169]. In an arsenic-contaminated agricultural field in Durgapur, India, a native B. cereus (PMM6) isolate exhibited resistance to arsenate (75 mM) and arsenite (25 mM), presumably through mechanisms of biosorption and bioaccumulation of these As forms [37]. This strain also displayed plant growth-promoting traits such as indole-3-acetic acid production, ACC deaminase activity, phosphate solubilization, and siderophore production, facilitating rice growth under arsenic stress [37,169], reducing arsenic mobilization, and thus preventing the generation of ROS.
It has also been reported that inoculation with B. subtilis S4 combined with iron nanoparticles helped mitigate arsenic-induced toxicity in Cucurbita moschata seeds by enhancing peroxidase (POD) and superoxide dismutase (SOD) activity [170], reducing levels of hydrogen peroxide, malonaldehyde, and electrolyte leakage. In other report, inoculation with B. amyloliquefaciens (NBRISN13) in combination with feldspar was found to reduce arsenite translocation and decrease As content in rice grains (52–72%). This treatment alleviated oxidative stress via modulation of enzymatic activity and phytohormone production, enhancing plant growth and yield [152,171]. B. amyloliquefaciens SN13 also reduced arsenic toxicity in rice by modulating the expression of carbohydrate metabolism and arsenic stress-related genes [45].
A consortium formed by B. licheniformis (NDSA24R), Priestia endophytica NDAS01F, and Priestia flexa NDAS28R isolated from Nadia, West Bengal, reduced As concentration in rice plants by lowering bioconcentration and translocation factors. This consortium induced systemic acquired resistance, stimulated sulfur metabolism and cell wall synthesis, and reduced oxidative stress in treated plants [43].
Another study evaluated the combination of B. faecalis and enriched composted biochar (ECB) to alleviate arsenic stress in maize plants. Results showed that this combination enhanced maize growth under arsenic exposure by regulating antioxidant production and reducing arsenic uptake by roots and leaves [172]. B. pacificus (AKS1), an arsenic-tolerant strain isolated from arsenic-contaminated groundwater in West Bengal, India, displays promising traits for environmental bioremediation. This species tolerates multiple metals and arsenic, growing in 20 mM arsenate and 10 mM arsenite. Genome analysis revealed the presence of As resistance genes, including arsC, arsB, and arsR. It also exhibited arsenic adsorption capacity and production of plant growth-promoting metabolites such as indole acetic acid (IAA), gibberellic acid (GA), and proline, along with nitrogen fixation capabilities [47].
4.3. Engineering Biosensors: Precision Detection of Arsenic
Some studies have employed genetic engineering to enhance the bioremediation potential of B. subtilis or to develop As biosensors. One notable work demonstrated the feasibility of heterologous expression in B. subtilis 168 of the arsenite-S-adenosylmethionine methyltransferase enzyme (CmarsM) from the thermophilic alga Cyanidioschyzon merolae, enabling the conversion of inorganic arsenic into dimethylarsinate and trimethylarsine. This modified strain was tested in arsenic-contaminated organic manure compost, resulting in significantly increased arsenic volatilization [173].
Understanding the ars operon has also been crucial in the development of As biosensors, leveraging the operon’s specific response to arsenic. Since the ars operon is regulated by the Ars repressor protein, genetic engineering allows the fusion of the promoter region (including the ArsR binding site) with a reporter gene. As(III) leads to repressor dissociation, allowing reporter expression. Biosensors in B. subtilis have been created by fusing the ars promoter to reporter genes such as β-galactosidase (lacZ) [70], luciferase, catecol-2,3-dioxygenase (xylE) [174,175], and GFP [71].
Furthermore, B. subtilis spores have been exploited to develop spore-based biosensors for easy portability, storage, transport, and field use. These spores germinate in the presence of samples and produce vegetative cells capable of responding to arsenic within a relatively short time, outperforming biosensors based on other bacteria. For instance, the spore-based biosensor using plasmid pMutin3-ars-lacZ retained detection ability after at least six months of storage at room temperature and could detect As concentrations as low as 1 × 10^−7^ M in freshwater and serum samples [71,175,176]. These spores can germinate in the presence of the sample and generate vegetative cells capable of responding to As in a relatively short time compared to biosensors of other bacteria. The As detection system based on B. subtilis spores with the pMutin3-ars-lacZ plasmid stored at room temperature maintained their arsenic detection capability after at least 6 months [70], being able to sense As concentrations from 1 × 10^−7^ M in freshwater and serum samples [176]. The spores of the pAD123-ars-gfpmut3a biosensor were able to germinate and emit fluorescent signals in less than 4 h in the presence of water artificially contaminated with arsenic from a concentration of 100 µM As(III) [71]. For the biosensor with the xylE reporter gene, it was observed that the sensitivity of the spore-based biosensor was low at an arsenic concentration of 1 mM but was strong and visible to the naked eye at a concentration of 5 mM As [175]. Although improvements to this biosensor allowed for decreased response times and minimum detection concentrations of As of up to 0.54 µg/L after 2 h of the addition of the enzymatic substrate catechol.
The use of biosensor spores has allowed the incorporation into a portable centrifugal microfluidic platform system in which quantitative response was achieved in an incubation time of 2.5 to 3 h, using small sample volumes and reagents [177]. Figure 8 summarizes several key biotechnological applications addressed in this section.
5. Concluding Remarks
B. subtilis and related species possess a multiplicity of strategies to detect and counteract the noxious effects of arsenic. While these bacteria rely on strategies of biosorption and transformation that prevent this metalloid from penetrating cells, they can deploy specific mechanisms to cope with its damaging effects [68,72,73]. Accordingly, evolution has equipped these species with genetic determinants that are specifically induced by As to generate sophisticated efflux pumps and reductases. This review has summarized the current knowledge of the complex, multi-faceted defense mechanisms these bacteria use. These strategies include immobilization and redox transformation, as well as highly regulated intracellular detoxification via the ars and ase operons. The core finding is that Bacillus does not deploy a single mechanism, but rather a coordinated arsenal. The initial line of defense involves biosorption and potential extracellular enzymatic transformation, which limits cellular uptake [31]. Once internalized, the ars operon—with its phage-derived, co-regulated components (arsR, yqcK, arsB/Acr3, and arsC)—and the chromosomally encoded ase operon (aseR and aseA) work together to reduce, sequester, and actively efflux the toxic metalloid. Our comparative genomic analysis highlights the diversity in genetic arrangements of these operons across different Bacillus species, underscoring an evolutionary trajectory shaped by vertical inheritance and horizontal gene transfer, particularly from mobile genetic elements like the SKIN prophage [74].
It is important to note that, despite the significant focus on expulsion and enzymatic detoxification, arsenic’s insidious effects extend to genetic material itself. Exposure to arsenic induces oxidative stress and direct DNA damage, posing a fundamental threat to genomic integrity. While the specific DNA repair pathways activated by Bacillus in response to arsenic-induced lesions remain largely unknown, the bacterium’s survival in highly contaminated environments implies their existence. Future research must prioritize elucidating these repair mechanisms, as enhancing genomic stability could be key to engineering hyper-resistant strains for bioremediation.
Environmental isolates of Bacillus further demonstrate the adaptability and biotechnological potential of these systems, showcasing diverse resistance strategies across contaminated ecosystems [111,131]. The in-depth understanding of resistance mechanisms employed by Bacillus has made it possible to exploit its biotechnological potential in bioremediation and sustainable agriculture [37,168,169,170,171]. Their ability to sporulate, their safety profile in numerous species, and their metabolic versatility make them ideal for practical applications. They are used as efficient biosorbents to remove As from wastewater, as inoculants to immobilize As in soil, and importantly, as plant growth-promoting bacteria (PGPR) to mitigate As toxicity in crops such as rice and wheat. Several Bacillus strains reduce the translocation of the metalloid to the edible parts of plants, thereby ensuring food safety, and improve plant growth by producing phytohormones, solubilizing phosphate, and exhibiting ACC deaminase activity. Therefore, arsenic-resistant Bacillus species are a powerful and versatile tool for developing bioremediation strategies and ensuring agricultural productivity in contaminated soils. Beyond bioremediation, B. subtilis emerges as an ideal platform for the development of next-generation As biosensors [70,71,176,177]. Its ability to form ultra-stable spores allows for the creation of portable, long-lasting, field-ready devices capable of detecting trace concentrations of As(III) in water and soil. Genetic engineering of the ars operon, fused to reporters such as GFP or luciferase, makes these bacteria sensitive, economical, and ecologically sustainable tools for real-time environmental and clinical monitoring.
Key challenges remain, such as elucidating the full diversity of As uptake pathways in bacteria from the genus Bacillus and understanding the ecological implications of horizontal gene transfer in As resistance dissemination. Future research should explore the integration of these microbial systems into bioremediation technologies, leveraging genetic engineering to enhance As sequestration, volatilization, or biosensor development. Additionally, the role of Bacillus in plant-associated As mitigation warrants deeper investigation to optimize sustainable agricultural practices in contaminated regions.
Addressing the global challenge of arsenic contamination requires a concerted and interdisciplinary strategy. Translating the knowledge of Bacillus molecular biology into practical applications necessitates synergistic collaboration among microbiologists, genetic engineers, environmental scientists, materials science experts, agronomists, artificial intelligence developers, electronic engineers, and data scientists. The integration of genomics, synthetic biology, and advanced materials engineering is essential for designing next-generation bioremediation strategies and high-precision biosensors. Consequently, the soil bacterium B. subtilis, with its sophisticated and evolutionarily refined arsenic resistance machinery, represents a powerful and versatile platform for developing safer and more sustainable environmental solutions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Errington J. van der Aart L.T. Microbe Profile: Bacillus subtilis: Model Organism for Cellular Development, and Industrial Workhorse Microbiology 202016642542710.1099/mic.0.00092232391747 PMC 7376258 · doi ↗ · pubmed ↗
- 2Kunst F. Ogasawara N. Moszer I. Albertini A.M. Alloni G. Azevedo V. Bertero M.G. Bessières P. Bolotin A. Borchert S. The Complete Genome Sequence of the Gram-Positive Bacterium Bacillus subtilis Nature 199739024925610.1038/367869384377 · doi ↗ · pubmed ↗
- 3Earl A.M. Losick R. Kolter R. Ecology and Genomics of Bacillus subtilis Trends Microbiol.20081626927510.1016/j.tim.2008.03.00418467096 PMC 2819312 · doi ↗ · pubmed ↗
- 4Saier M.H.Jr. Goldman S.R. Maile R.R. Moreno M.S. Weyler W. Yang N. Paulsen I.T. Transport Capabilities Encoded within the Bacillus subtilis Genome J. Mol. Microbiol. Biotechnol.20024376711763970 · pubmed ↗
- 5Alcaraz L.D. Moreno-Hagelsieb G. Eguiarte L.E. Souza V. Herrera-Estrella L. Olmedo G. Understanding the Evolutionary Relationships and Major Traits of Bacillus through Comparative Genomics BMC Genom.20101133210.1186/1471-2164-11-332PMC 289056420504335 · doi ↗ · pubmed ↗
- 6Rooney A.P. Price N.P.J. Ehrhardt C. Sewzey J.L. Bannan J.D. Phylogeny and Molecular Taxonomy of the Bacillus subtilis Species Complex and Description of Bacillus subtilis subsp. inaquosorum subsp. nov.Int. J. Syst. Evol. Microbiol.2009592429243610.1099/ijs.0.009126-019622642 · doi ↗ · pubmed ↗
- 7Zeigler D.R. Prágai Z. Rodriguez S. Chevreux B. Muffler A. Albert T. Bai R. Wyss M. Perkins J.B. The Origins of 168, W 23, and Other Bacillus subtilis Legacy Strains J. Bacteriol.20081906983699510.1128/JB.00722-0818723616 PMC 2580678 · doi ↗ · pubmed ↗
- 8Vaughan D.J. Sulfide Mineralogy and Geochemistry: Introduction and Overview Rev. Miner. Geochem.2006611510.2138/rmg.2006.61.1 · doi ↗