Self-resistance mechanism to acyldepsipeptide antibiotics in the Streptomyces producer
Dhana Thomy, Laura Reinhardt, Elisa Liebhart, Mirita Franz-Wachtel, Boris Maček, Peter Sass, Heike Brötz-Oesterhelt

TL;DR
This paper explains how Streptomyces bacteria protect themselves from their own antibiotic ADEP by using a unique ClpP protein.
Contribution
The study reveals a novel self-resistance mechanism involving ClpPADEP in Streptomyces producers of ADEP antibiotics.
Findings
ClpPADEP interferes with ClpP1P2 complex formation and inhibits ClpP1 proteolytic activity.
ClpPADEP forms functional complexes with ClpP2 and Clp-ATPases to maintain essential proteolytic functions.
ClpPADEP provides dual protection by preventing ADEP-induced degradation and preserving housekeeping Clp functions.
Abstract
Clp proteases are ubiquitous in bacteria and play an important role in regulatory proteolysis and in maintaining protein homeostasis within the bacterial cell. They consist of a tetradecameric, proteolytic ClpP core and associated AAA+ Clp-ATPases. The Clp system of Streptomyces is unusually complex, comprising up to five ClpP homologs (ClpP1–ClpP5) and three Clp-ATPases (ClpX, ClpC1, and ClpC2). Streptomycetes produce a plethora of secondary metabolites, including potent acyldepsipeptide (ADEP) antibiotics, which target ClpP. We have previously reported on the operation mode of the Streptomyces ClpP1P2 protease and identified a novel clpP gene (named clpPADEP) as a resistance determinant encoded near the ADEP biosynthesis gene cluster. However, the molecular function of ClpPADEP remains enigmatic. Here, we report on the molecular self-resistance mechanism to ADEP via ClpPADEP and its…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
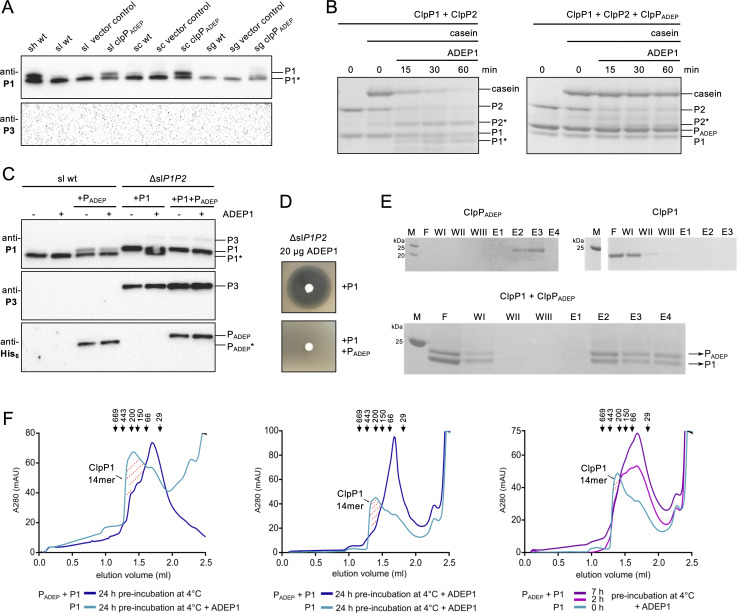 Fig 1
Fig 1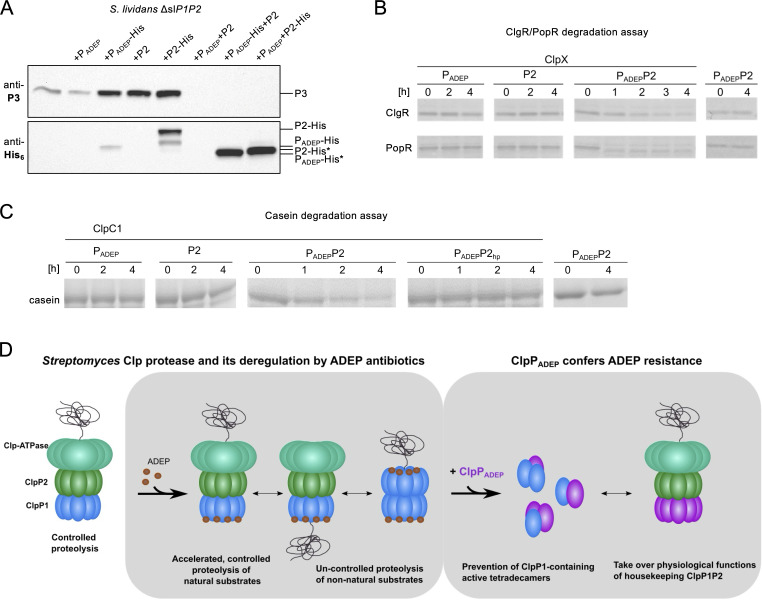 Fig 2
Fig 2- —Deutsche Forschungsgemeinschafthttp://dx.doi.org/10.13039/501100001659
- —Deutsche Forschungsgemeinschafthttp://dx.doi.org/10.13039/501100001659
- —Deutsche Forschungsgemeinschafthttp://dx.doi.org/10.13039/501100001659
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Microbial Natural Products and Biosynthesis · Genomics and Phylogenetic Studies
OBSERVATION
Acyldepsipeptide (ADEP) antibiotics are highly active against many Gram-positive species, including multi-resistant staphylococci, streptococci, and enterococci, with minimum inhibitory concentrations in the sub-µg/mL range. Accordingly, ADEP showed high efficacy in treating staphylococcal, enterococcal, and streptococcal infections in rodents (1, 2), and in combination with rifampicin, even eradicated persister cells in chronic biofilm infections (3). ADEP targets and deregulates ClpP, the core of the bacterial caseinolytic protease Clp (4, 5). The Clp protease is ubiquitous in bacterial cells, where it plays a crucial role in regulatory proteolysis and protein homeostasis (6, 7). Clp is a self-compartmentalizing serine protease composed of the proteolytic, tetradecameric core ClpP and cognate, hexameric AAA+ unfoldases (i.e., Clp-ATPases, such as ClpX and ClpC in Staphylococcus aureus), which recognize, unfold, and feed substrates of the Clp system into the barrel-shaped ClpP tetradecamer (8, 9). In Firmicutes, ADEP inhibits regulatory proteolysis of natural Clp protein substrates by abrogating the interaction of the proteolytic core ClpP with its partner AAA+ Clp-ATPases (5, 10–12) while activating the ClpP core for uncontrolled proteolysis via direct binding and conformational control (13–16).
Bacteria exposed to ADEP under laboratory conditions develop resistance, which entails spontaneous mutations in ClpP affecting its oligomerization, conformation, catalytic function, or interaction with partnering Clp-ATPases or ADEP (3, 17–20). However, these mutations also impair the function of the Clp protease in regulatory proteolysis and protein homeostasis. Since loss of ClpP function severely attenuates the ability of multiple pathogenic bacteria to colonize and infect rodents, it is tempting to speculate that such spontaneous loss-of-function mutations will less frequently occur in the host environment (4, 17). Due to the complexity of the essential Clp system in Streptomyces (21) and the multi-layered activities of ADEP, spontaneous loss-of-function mutations can also not explain the self-resistance of the ADEP producer Streptomyces hawaiiensis NRRL 15010.
Streptomycetes possess, in fact, one of the most complex Clp machineries reported in eubacteria so far. For example, the genome of the model organism Streptomyces lividans (sl) encodes at least three Clp-ATPases (ClpX, ClpC1, and ClpC2) and five ClpP homologs (ClpP1–ClpP5) (21, 22), and individual ClpP proteins show high homology among different Streptomyces strains (Fig. S1A through E). In contrast, most other bacterial genomes encode a single or two ClpP homologs, the latter often interacting to form ClpP1P2 hetero-tetradecamers. We have recently provided molecular insights into the operation mode of the ClpP1P2 complex in Streptomyces (18). We have shown that a ClpP1P2 heterotetradecamer forms the proteolytic core of the housekeeping Clp protease, with ClpP1 mainly accountable for the proteolytic activity of the ClpP1P2 complex, and ClpP2 for the recruitment of the Clp-ATPases. One of the natural substrates of ClpP1P2 is the transcriptional activator of the clpP3P4 operon, PopR. Its continuous degradation by ClpP1 (within the ClpP1P2 complex) ensures that the non-isofunctional ClpP3P4 core is expressed as a backup only in situations when ClpP1 or ClpP2 is not functioning (18, 23). Furthermore, while ClpP1 has been identified as a target for ADEP (18, 24), ClpP2 and ClpP3P4 were found to be ADEP-insensitive (18, 24), that is, ADEP binds to and deregulates ClpP1, but not ClpP2, and the interaction of ClpP2 with ClpX, ClpC1, or ClpC2 is not prevented by ADEP (18).
Recently, we identified a sixth clpP gene (named clpP_ADEP_) in the vicinity of the ADEP biosynthetic gene cluster in the ADEP producer S. hawaiiensis NRRL 15010 (25). Initial cell-based studies revealed that this accessory ClpP homolog acts as a resistance determinant against ADEP, conferring high-level ADEP resistance when heterologously expressed in ADEP-sensitive streptomycetes (25). However, the underlying molecular mechanism that leads to ADEP resistance remained unsolved. In this study, by combining cell-based and in vitro studies, we clarify the molecular basis of ClpP_ADEP_-mediated resistance, thereby revealing a novel mechanism of producer self-resistance.
Degradation of ClpP1 or expression of ADEP-insensitive ClpP3 is not part of the self-resistance mechanism of the ADEP producer
To elucidate the mechanism of ClpP_ADEP_-mediated resistance, we first tested the hypothesis that ClpP_ADEP_ may interact with the ADEP target ClpP1 to inhibit its function. ClpP1 inactivation would allow Streptomyces to survive via accumulation of the ClpP1P2 substrate PopR, an activator of the expression of the ADEP-insensitive ClpP3P4 (21). Such a survival strategy was previously reported for spontaneous, ADEP-resistant S. lividans mutants, which carried loss-of-function mutations in the clpP1 gene resulting in ClpP3P4 expression (24). Indeed, amino acid sequence alignments and phylogenetic analyses revealed that ClpP_ADEP_ shows the highest homology to ClpP1 (Fig. S1F), suggesting that an interaction between ClpP_ADEP_ and ClpP1 may be possible. Furthermore, we determined the transcript and protein levels of ClpP1, ClpP2, and ClpP_ADEP_ in the ADEP producer S. hawaiiensis. qPCR analysis revealed that clpP_ADEP_ was expressed already in the early exponential phase and transcript levels of clpP_ADEP_ and clpP1 closely paralleled each other throughout the different growth phases of the producer culture, while clpP2 transcript levels were slightly lower (Fig. S2). The protein level of ClpP_ADEP_, detected by quantitative proteomics, was considerable and reached approximately half the amount recorded for ClpP1 and ClpP2 (Fig. S3).
To investigate whether ClpP1 inactivation might occur by degradation, we analyzed cell extracts of the ADEP-producer S. hawaiiensis (sh), as well as of wild-type S. lividans (sl), Streptomyces coelicolor (sc), and Streptomyces griseus (sg), and ADEP-resistant mutants thereof that heterologously expressed ClpP_ADEP_. Western Blot analysis revealed the full-length ClpP1 protein and its naturally processed form ClpP1* in the extracts of all the strains (Fig. 1A). Thus, ClpP_ADEP_-mediated detoxification of ClpP1 does not occur by proteolytic degradation. Of note, Streptomyces ClpP proteins undergo processing reactions both in vitro and in living cells (18). As shown previously, ClpP1 processing requires the presence of ClpP2 and a Clp activator (e.g., either a Clp-ATPase interacting with ClpP2 or ADEP interacting with ClpP1), whereas ClpP2 processing depends exclusively on the presence of ClpP1, but not on a Clp-ATPase or ADEP. However, the processing of both ClpP1 and ClpP2 depends entirely on the integrity of the ClpP1 catalytic triad (18). In our current study, we noted that unprocessed ClpP1 occurred in higher amounts in extracts of cells that expressed ClpP_ADEP_, suggesting a ClpP_ADEP_-mediated inhibition of ClpP1 self-processing. For more details on the processing of ClpP proteins in Streptomyces, the reader is kindly referred to our recent publication by Reinhardt et al. (18).
ClpPADEP disrupts toxic, ADEP-activated ClpP1 complexes. (A). Immunoblotting of cell extracts of S. hawaiiensis wild type (sh wt) as well as wild type or ClpPADEP-expressing mutants of S. lividans (sl), S. coelicolor (sc), and S. griseus (sg). Anti-Streptomyces ClpP1 (anti-P1) or ClpP3 (anti-P3) antibodies were used as indicated. ClpP3 expression was used as a readout for ClpP1 inactivation. Vector control, pSETermE (B). In vitro protease activity assays using heterologously expressed and isolated S. hawaiiensis ClpP proteins and casein as a model substrate. See Fig. S5 for replicate experiments. (C). Immunoblotting of cell extracts of ADEP-treated S. lividans wild type (sl wt) and S. lividans ΔP1P2 (ΔslP1P2) complemented with slclpP1 and/or clpPADEP as indicated. Strains were grown for 24 h and subsequently treated (+) with 5 µg/mL of ADEP1 for another 3 hours, upon which crude cell extracts were prepared and analyzed by immunoblotting using anti-P1, anti-P3, or anti-His6 antibodies. ClpP3 expression was used as a readout for ClpP1 inactivation. (D). ADEP disk diffusion bioassay of S. lividans ΔP1P2 complemented with either slclpP1 or both slclpP1 and clpPADEP. (E). Pull-down experiment applying pre-incubated mixtures of untagged shClpP1 and ClpPADEP-His6 to nickel-nitrilotriacetic acid agarose. SDS-PAGE of flow-through (F), wash steps (W I–III), and elution fractions (E 1–4), M, protein marker. See Fig. S6 for full-size SDS PAGE. (F). Size exclusion chromatography of purified S. hawaiiensis ClpP1 and ClpPADEP proteins in the absence or presence of ADEP1. For a size exclusion experiment demonstrating the conformational control that ADEP exerts on ClpP1, that is, inducing the active, extended conformation of ClpP1, see Fig. 3H in Reinhardt et al. (18). P1, unprocessed ClpP1; P1*, processed ClpP1; P2, unprocessed ClpP2; P2*, processed ClpP2; PADEP, ClpPADEP.*
Notably, in the same Western blot assay, we also used an anti-Streptomyces ClpP3 (anti-P3) antibody as a readout for ClpP1 inactivation and did not detect ClpP3 expression, which indicates that the PopR-degrading natural function of the ClpP1P2 complex was maintained in the ADEP producer. Likewise, ClpP3 was absent in ADEP-resistant S. lividans, S. coelicolor, and S. griseus that heterologously expressed ClpP_ADEP_ (Fig. 1A).
ClpPADEP confers ADEP resistance by disrupting ADEP-activated ClpP1 complexes
Next, we conducted an in vitro protease activity assay using heterologously expressed ClpP proteins and casein as a model substrate (Fig. 1B). While casein was efficiently degraded by ClpP1P2 in the presence of ADEP, the addition of ClpP_ADEP_ inhibited processing of ClpP1 as well as casein degradation by ADEP-activated ClpP1P2, indicating a direct effect of ClpP_ADEP_ on the function of ClpP1.
To further examine a potential effect of ClpP_ADEP_ on the function of ClpP1 in Streptomyces cells, we analyzed the effects of inducible expression of clpP_ADEP_ in S. lividans wild type and of slclpP1 and clpP_ADEP_ in a ΔP1P2 background, that is, in the presence or absence of ClpP2, respectively (Fig. 1C). Here, using an anti-P1 antibody, we confirmed that the expression of ClpP_ADEP_ slows down auto-processing of ClpP1 in S. lividans wild type, since bands of unprocessed ClpP1 could still be detected. In the S. lividans ΔP1P2 background, ClpP_ADEP_ entirely prevented ADEP-induced ClpP1 auto-processing, and ClpP_ADEP_ itself was only detected in its unprocessed form.
Notably, in S. lividans ΔP1P2, we detected ClpP3 when either ClpP1 or ClpP_ADEP_ alone or both were introduced, but ClpP2 was lacking. These results indicate that although ClpP_ADEP_ must interact with ClpP1 to affect its functions, no proteolytically active complex is formed between these two ClpP proteins that could substitute for the housekeeping functions of the ClpP1P2 complex. Visible proof is the induction of ClpP3 expression, as a readout for ClpP1 inactivation, and the failed processing of ClpP1 and ClpP_ADEP_.
Of note, using an ADEP bioassay, S. lividans ΔP1P2 was found to be sensitive to ADEP treatment when complemented with ClpP1 alone, showing that the antibacterial activity of ADEP does not require ClpP2, while S. lividans ΔP1P2 was ADEP-resistant when complemented with both ClpP1 and ClpP_ADEP_ (Fig. 1D).
To obtain more information about the nature of the interaction between ClpP1 and ClpP_ADEP_, we pre-incubated a mixture of untagged shClpP1 with ClpP_ADEP_-His_6_ followed by pull-down with nickel-nitrilotriacetic acid. Here, despite extensive washing with 20 mM imidazole, we observed co-elution of both shClpP1 and ClpP_ADEP_-His_6_ in a similar ratio, indicating a direct interaction of both homologs (Fig. 1E). In addition, we examined complex formation between shClpP1 and ClpP_ADEP_ in the absence or presence of ADEP by size exclusion chromatography (Fig. 1F). Pre-incubation of shClpP1 with ADEP induced the formation of tetradecameric shClpP1 complexes as reported previously (18). However, the presence of ClpP_ADEP_ prevented ADEP-induced tetradecamer formation of shClpP1, and only ClpP oligomers smaller than 70 kDa could form, representing ClpP dimers and/or trimers. Of note, ClpP_ADEP_ alone eluted mostly in lower oligomeric states (Fig. S4). Hence, the interplay of ClpP1 and ClpP_ADEP_ involves a strong, direct interaction of both homologs resulting in mixed ClpP1/ClpP_ADEP_ dimers and trimers, thereby preventing the formation of catalytically active, tetradecameric ClpP complexes of both homomeric and heteromeric nature, even in the presence of the tetradecamer-inducing activator ADEP.
ClpPADEP ensures natural substrate degradation by forming an ADEP-insensitive ClpPADEPP2-Clp-ATPase complex
Although our data indicated that ClpP_ADEP_ interferes with ClpP1 oligomerization and activity (Fig. 1B, E, and F), important cellular functions, which commonly rely on an operational ClpP1P2 complex, appeared to run unhindered in S. hawaiiensis. A clear indication was the lack of ClpP3 expression in ClpP_ADEP_-containing cells (Fig. 1A and C). Therefore, we next explored a putative interaction of ClpP_ADEP_ with ClpP2 in Streptomyces cells in the absence of slClpP1 by complementing S. lividans ΔP1P2 with clpP_ADEP_, slclpP2, or both. To allow detection of ClpP_ADEP_ or ClpP2 in immunoblots, proteins were expressed as either untagged or His_6_-tagged versions. Then, we used these strains in immunoblotting experiments to study ClpP3 expression (Fig. 2A). Once again, the absence of ClpP3 served as a proxy for a functional ClpP complex capable of degrading PopR, the transcriptional activator of the clpP3P4 operon (18). Interestingly, ClpP3 expression was observed when either ClpP_ADEP_ or ClpP2 was present alone, but ClpP3 expression was lost when both ClpP_ADEP_ and ClpP2 were co-expressed. Furthermore, ClpP homologs were only processed when ClpP_ADEP_ and ClpP2 were co-expressed, as indicated by anti-His_6_ antibody detection. These cell-based data implied that ClpP_ADEP_ and ClpP2 form a proteolytically active, heteromeric complex, which allows processing of both ClpP_ADEP_ and ClpP2 and finally enables the degradation of natural ClpP1P2 substrates, for example, PopR, as indicated by the repressed expression of ClpP3.
ClpPADEP ensures natural substrate degradation by the formation of an ADEP-insensitive ClpPADEPP2-Clp-ATPase complex. (A) Immunoblotting of cell extracts of S. lividans ΔP1P2 complemented with either clpPADEP, slclpP2, or both, as indicated. Strains were grown for 24 h and crude cell extracts were prepared and analyzed by immunoblotting using anti-P3 and anti-His6 antibodies. ClpP3 expression was used as a readout for ClpP1 inactivation. Of note, in S. lividans ΔP1P2, a double band is detected for ClpP2-His using anti-His6 antibodies as previously observed (18). (B) In vitro protease activity assays using heterologously expressed and isolated S. hawaiiensis proteins ClpPADEP and ClpP2, the Clp-ATPase protein ClpX, as well as the natural ClpXP1P2 substrates ClgR and PopR. See Fig. S7 for replicate experiments. (C) In vitro protease activity assays using the isolated S. hawaiiensis proteins ClpPADEP and ClpP2, the Clp-ATPase protein ClpC1, as well as the model protein substrate β-casein. In addition, a ClpP2 hydrophobic pocket mutant (ClpP2hp) was used to analyze Clp-ATPase binding. See Fig. S8 for replicate experiments. (D) Model of ADEP mode of action and mode of resistance in Streptomyces. For further details on the mode of action, see (18). ClpPADEP prevents the formation of toxic ClpP1-containing tetradecamers and forms a new proteolytic core with ClpP2 to take over essential natural functions. P2, unprocessed ClpP2; P2, processed ClpP2; PADEP, unprocessed ClpPADEP; PADEP*, processed ClpPADEP.*
To corroborate the results of our cell-based experiments, we performed in vitro protease activity assays using the isolated S. hawaiiensis proteins ClpP_ADEP_ and ClpP2, the cognate Clp-ATPase ClpX, as well as the natural ClpXP1P2 substrates ClgR and PopR. When testing different combinations of ClpP_ADEP_, shClpP2, and ClpX to attempt the digestion of ClgR and PopR (Fig. 2B), we found that neither ClpP2 nor ClpP_ADEP_ alone was capable of protein degradation with ClpX. However, when ClpP_ADEP_, ClpP2, and ClpX were combined, ClgR and PopR were digested, thus validating that ClpP_ADEP_ and ClpP2 jointly substitute for the ClpP1P2 core in natural protein degradation. Likewise, when we used the Clp-ATPase ClpC1 in in vitro degradation assays with the model protein substrate ß-casein, proteolysis occurred only when ClpP_ADEP_, ClpP2, and ClpC1 were combined (Fig. 2C). Of note, when ClpP2 was mutated in its Clp-ATPase interaction site, the hydrophobic pocket, and thereby rendered incapable of Clp-ATPase binding (18), the degradation of ß-casein was prevented. Our data, therefore, consistently show that ClpP_ADEP_ and ClpP2 form a proteolytically active heteromeric Clp protease core that interacts with either ClpX or ClpC1, with ClpP2 representing the interaction partner for the Clp-ATPases.
In summary, our data unravel the molecular basis of a novel mechanism of producer self-resistance. Here, the mode of action of the ClpP-deregulating ADEP antibiotic is matched by a self-resistance mechanism conferred by ClpP_ADEP_, which is far more complex than simple target mutation (Fig. 2D). The presence of an extra, ADEP-resistant ClpP_ADEP_ protein protects the cell from the detrimental consequences of ADEP by (i) preventing tetradecamer formation of the ADEP target ClpP1 and consequently inhibiting its catalytic activity and (ii) substituting for ClpP1 in a complex with ClpP2 and a Clp-ATPase to preserve the housekeeping functions of the Clp protease in regulatory proteolysis and protein homeostasis, even in the presence of ADEP antibiotics. The combination of these diverse abilities in a single protein is remarkable, as it requires direct interaction with two different ClpP partners, on the one hand, preventing and, on the other hand, newly generating catalytic function.
One might wonder why this rather complex resistance mechanism evolved in the ADEP producer instead of a mere out-of-function point mutation in the ADEP target ClpP1. A plausible explanation could be a diverging substrate spectrum of the non-isofunctional pairs ClpP1P2 and ClpP3P4, probably mediated or enhanced by a distinct interaction preference for the Clp-ATPases. The full substrate spectrum of those Streptomyces proteases is elusive, but it was shown that an S. lividans clpP1 mutant had a cell cycle defect (21). It presented a “bald” phenotype by arrest in the basal mycelial state and loss of sporulation capacity. Expressing clpP3 and clpP4 in this strain did not rescue the bald phenotype (21). Vice versa, a clpP3 mutant was not affected in its cell cycle but produced more of the blue antibacterial pigment actinorhodin (21).
Introducing the phylogenetically distinct ClpP_ADEP_ to modulate the multi-layered Clp system in Streptomyces seems to have been overall worthwhile, but still a challenge for nature, as suggested by the fact that S. hawaiiensis NRRL 15010 is the only known ADEP producer known to date.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brötz-Oesterhelt H, Beyer D, Kroll H-P, Endermann R, Ladel C, Schroeder W, Hinzen B, Raddatz S, Paulsen H, Henninger K, Bandow JE, Sahl H-G, Labischinski H. 2005. Dysregulation of bacterial proteolytic machinery by a new class of antibiotics. Nat Med 11:1082–1087. doi:10.1038/nm 130616200071 · doi ↗ · pubmed ↗
- 2Brown Gandt A, Griffith EC, Lister IM, Billings LL, Han A, Tangallapally R, Zhao Y, Singh AP, Lee RE, La Fleur MD. 2018. In vivo and in vitro effects of a Clp P-activating antibiotic against vancomycin-resistant enterococci. Antimicrob Agents Chemother 62:e 00424-18. doi:10.1128/AAC.00424-1829784838 PMC 6105829 · doi ↗ · pubmed ↗
- 3Conlon BP, Nakayasu ES, Fleck LE, La Fleur MD, Isabella VM, Coleman K, Leonard SN, Smith RD, Adkins JN, Lewis K. 2013. Activated Clp P kills persisters and eradicates a chronic biofilm infection. Nature 503:365–370. doi:10.1038/nature 1279024226776 PMC 4031760 · doi ↗ · pubmed ↗
- 4Brötz-Oesterhelt H, Vorbach A. 2021. Reprogramming of the caseinolytic protease by ADEP antibiotics: molecular mechanism, cellular consequences, therapeutic potential. Front Mol Biosci 8:690902. doi:10.3389/fmolb.2021.69090234109219 PMC 8182300 · doi ↗ · pubmed ↗
- 5Brötz-Oesterhelt H, Sass P. 2014. Bacterial caseinolytic proteases as novel targets for antibacterial treatment. Int J Med Microbiol 304:23–30. doi:10.1016/j.ijmm.2013.09.00124119566 · doi ↗ · pubmed ↗
- 6Olivares AO, Baker TA, Sauer RT. 2016. Mechanistic insights into bacterial AAA+ proteases and protein-remodelling machines. Nat Rev Microbiol 14:33–44. doi:10.1038/nrmicro.2015.426639779 PMC 5458636 · doi ↗ · pubmed ↗
- 7Illigmann A, Thoma Y, Pan S, Reinhardt L, Brötz-Oesterhelt H. 2021. Contribution of the Clp protease to bacterial survival and mitochondrial homoeostasis. Microb Physiol 31:260–279. doi:10.1159/00051771834438398 · doi ↗ · pubmed ↗
- 8Gottesman S, Roche E, Zhou Y, Sauer RT. 1998. The Clp XP and Clp AP proteases degrade proteins with carboxy-terminal peptide tails added by the Ssr A-tagging system. Genes Dev 12:1338–1347. doi:10.1101/gad.12.9.13389573050 PMC 316764 · doi ↗ · pubmed ↗