Thiamine Diphosphate Supplementation as a Heat-Stress Mitigation Strategy for Hair Male and Female Lambs in Feedlot: Physiological Responses, Growth Performance, and Carcass Traits
Ulises Macías-Cruz, German Castillo Cristóbal, Leonel Avendaño-Reyes, María de los Ángeles López-Baca, José A. Roque-Jiménez, Miguel Mellado, César A. Meza-Herrera, Ricardo Vicente-Pérez, Marisol López-Romero, Nallely Rivero-Pérez

TL;DR
Adding thiamine diphosphate to feed helps female lambs handle heat stress better by improving energy use, but it doesn't work as well for males.
Contribution
Thiamine diphosphate improves energy efficiency in female lambs under heat stress without affecting meat quality or growth.
Findings
Thiamine diphosphate increased heat loss through the body surface in both male and female lambs.
Female lambs had reduced feed intake and improved carcass traits with thiamine supplementation.
Male lambs did not show significant improvements in growth or carcass traits with thiamine.
Abstract
High summer temperatures in desert regions decrease weight gain, feed efficiency, carcass weight, and meat tenderness in feedlot sheep, which is associated with insufficient availability of dietary energy by increasing energy requirements for thermoregulation. So, hair male and female lambs were fed thiamine diphosphate to evaluate its effect as glucogenic cofactor on physiological variables, feedlot growth, carcass traits and meat quality under hot desert conditions. Thiamine improved heat loss along the body surface, without changing rectal temperature and respiratory rate. Furthermore, it reduced feed intake in females but not in males. Regardless of gender, thiamine did not modify growth rate, carcass yield, fat deposition, and color and hardness in meat. Therefore, this heat stress mitigation strategy effectively increased dietary energy efficiency in ewe lambs but was ineffective…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
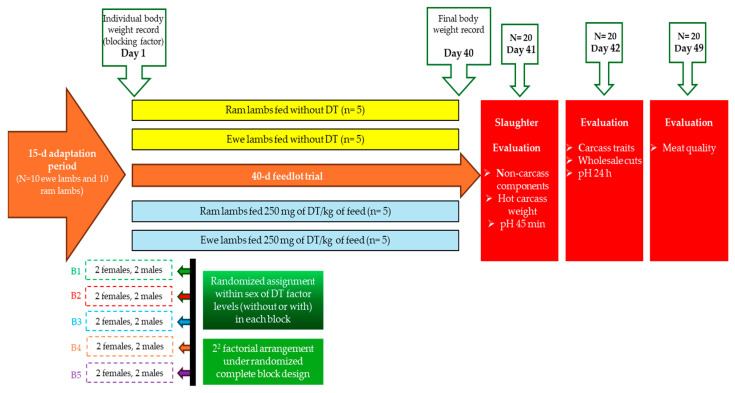 Figure 1
Figure 1 Figure 2
Figure 2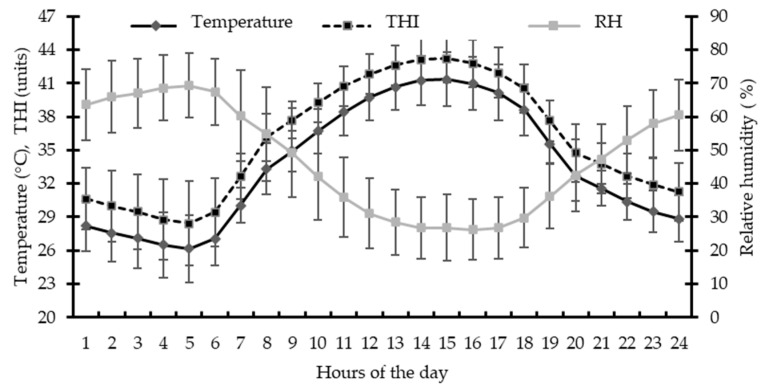 Figure 3
Figure 3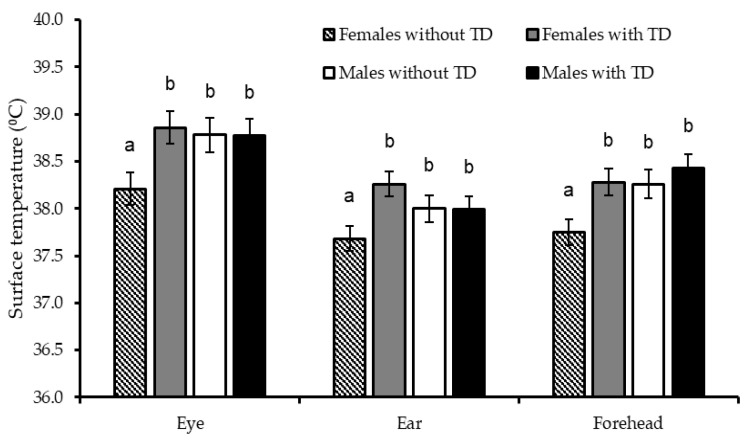 Figure 4
Figure 4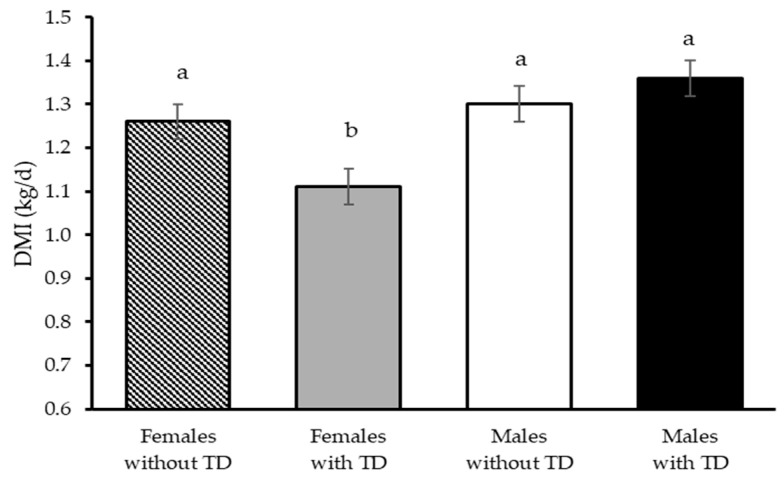 Figure 5
Figure 5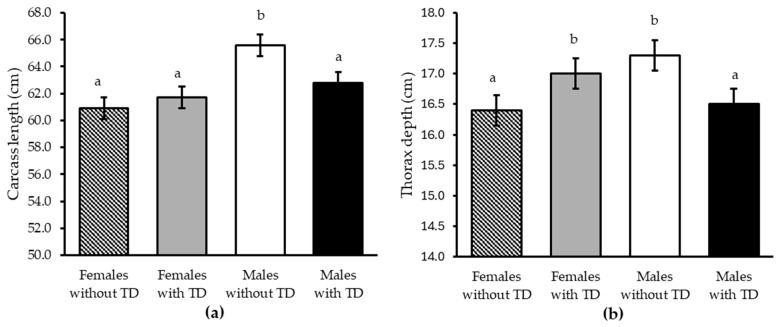 Figure 6
Figure 6- —Autonomous University of Baja California
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEffects of Environmental Stressors on Livestock · Meat and Animal Product Quality · Exercise and Physiological Responses
1. Introduction
Hair sheep are characterized by growing and producing meat under summer heat stress (HS) conditions in arid and desert regions [1]. Despite this behavior, the summer HS decreases the growth potential of fattening male and ewe lambs of these breeds by 20 to 30%, which negatively impacts muscle deposition, carcass weight, and meat quality [2,3]. These adverse effects of HS on growth and carcass traits are not ascribed to decreased feed intake, as in other sheep breeds, but rather to a redistribution of dietary energy that favors thermoregulation processes more than growth [4]. Given that these lambs have low availability of metabolizable energy (ME < 1.8 Mcal/kg dry matter) for growth, and their feed intake is maintained under HS, an alternative strategy for mitigating HS could be dietary supplementation of glucogenic cofactors.
Thiamine diphosphate (TD) is a glucogenic cofactor derived from vitamin B1, which functions as a coenzyme for enzymes involved in the cell’s bioenergetic processes that lead to adenosine triphosphate (ATP) synthesis [5]. Thus, the enzymes pyruvate dehydrogenase, α-ketoglutarate dehydrogenase, transketolase, and branched-chain α-ketoacid dehydrogenase require TD as a cofactor in glycolysis and oxidative carbohydrate decarboxylation [6,7]. This response, in turn, stimulates greater ATP production and metabolizable energy by promoting better glucose utilization, as well as the availability of gluconeogenic substrates [8,9]. Based on the above, this glycogenic cofactor can enhance dietary energy efficiency mainly by aiding cells in converting ingested carbohydrates into energy through metabolic pathways (glycolysis) that optimize ATP production from glucose oxidation; likewise, if necessary, it also contributes to promoting the synthesis of gluconeogenesis precursors from fatty acids and short-chain amino acids from the diet [6,10]. In addition, it counteracts inflammatory processes and oxidative stress [11], two common problems triggered in sheep exposed to high environmental temperatures (Ta) [12]. In this way, TD in heat-stressed lambs has the potential to enhance not only dietary energy efficiency but also redox status, immune system function, and overall welfare.
To date, some studies have evaluated supplemental TD on the productive performance and carcass traits of fattening lambs, but only under thermoneutral conditions (12 to 30 °C), reporting inconsistent findings. Two experiments were conducted with Rambouillet wethers and female lambs using the same thiamine treatments (0, 50, 100, and 150 mg/head), and the difference between them was group or individual feeding [13]. Supplemental thiamine in group feeding, but not in individual feeding, was effective to increase weight gain, feed efficiency, dry matter intake (DMI), and carcass weight in a dose-dependent manner, while other carcass traits were unaffected (i.e., conformation, Longissimus thoracis [LT] muscle area, backfat thickness, and quality grade). In Afec-Assaf male lambs, subcutaneous injections of high-dose thiamine improved glucose metabolism and hepatic glycogen storage with no growth rate improvement [10]. Note that while TD feeding has not been investigated in heat-stressed fattening lambs of any breed, this vitamin has been shown to partially reverse the adverse effects of HS on milk production and quality, but not on DMI and body condition of lactating ewes [11]. This response advocates that TD can increase dietary energy efficiency and redirect this energy to improve productive variables of animals experiencing HS.
It is important to note that metabolic sexual dimorphism in fattening lambs could play an essential role in the effectiveness of TD to improve the availability of dietary energy for growth. The gonadal endocrine activity adjusts the energy and protein metabolism to prioritize the distribution of nutrients in different tissues according to gender [14]. While ovarian-synthesized estrogens favor the transformation of nutrients into adipose tissue rather than muscle tissue in ewe lambs, testicular testosterone in male lambs acts as a natural anabolic, prioritizing energy use for protein synthesis and lean muscle tissue deposition [15]. Thus, male lambs compared to female lambs under a feedlot systema show superiority in muscularity, growth performance, and carcass weight and size, but regularly lower carcass yield and body fat deposition, with contradictory gender effects on meat quality even though male lambs tend to accumulate more muscle glycogen [15,16]. Although HS partially or totally disrupts reproductive events in mammals by inactivating the endocrine reproductive axis with the activation of the hypothalamic–pituitary–adrenal axis [17], hair sheep appear to have reproductive resilience to HS, as both males and females can reproduce under high Ta of desert regions [18,19]. In this context, it was hypothesized that feeding TD to feedlot hair sheep improves productive performance, carcass traits, and meat quality without compromising physiological thermoregulatory capacity, especially noticeable in male lambs compared to females, under high summer Ta. Therefore, this study aimed to evaluate the effects of supplemental TD and gender on the physiological responses, feedlot growth, carcass traits, and meat quality of heat-stressed hair breed lambs during the summer in a desert region.
2. Materials and Methods
2.1. Study Site and Experimental Period
The study was conducted during the hottest summer months (July and August) at the Sheep Experimental Unit of the Institute of Agricultural Sciences, Autonomous University of Baja California (UABC), located in the Mexicali Valley, Baja California, Mexico (32.38° N and 115.28° W). The region presents a hot desert climate (BWh), with maximum temperatures exceeding 45 °C in the summer and minimum temperatures of 0 °C in winter; the average annual rainfall is 77.8 mm, mostly occurring in winter [20].
2.2. Animals and Pre-Experimental Handling
Twenty Dorper × Kathadin crossbred lambs (10 females and 10 males) were used and managed according to the Mexican Official Standards: NOM-051-ZOO-1995 [21] (establishes the humane treatment of animals during their mobilization), NOM-062-ZOO-1999 [22] (describes the management of production, care, use and health protection of laboratory animals), and NOM-033-SAG/ZOO-2014 [23] (specifies the methods for the humane slaughter of domestic and wild animals). Both protocol and field phase were also revised and authorized by the UABC Ethics Committee (Letter number: 047/2024-1).
All lambs received a single prophylactic handling, 30 d before starting the feedlot trial, as well as they were adapted to individual pens and to a basal experimental diet for 15 d. Prophylactic handling consisted of treating each animal with 3.0 mL of oral dewormer (Adbendal 10% Co^®^, Adler Pharma Laboratory, Jalisco, Mexico), 0.5 mL of A-D-E vitamins intramuscularly (Vigantol ADE Fuerte^®^, Elanco Animal Health Laboratory, Guadalajara, Jalisco, Mexico), and 2.5 mL of clostridium vaccine subcutaneously (Bovimune^®^ Clostri 10, LaPisa Laboratory, La Piedad, Michoacan, Mexico). The individual pens were built in an area of 2.0 m^2^, with cyclonic mesh walls and galvanized sheet shade installed at a height of 2.5 m on a dirt floor. Each pen was provided with a feed bunk and water trough. The experimental diet was formulated to meet the nutritional requirements for fattening lambs [24] by mixing the ingredients indicated in Table 1.
2.3. Experimental Design and Handling
On the first day of the 40-d feedlot test, lambs were about 4 months old and were weighed individually (BW = 31 ± 2.8 kg). Blocks of two male lambs and two ewe lambs of similar BW were formed and then lambs of each block were randomly assigned within each gender to one of two dietary strategies: basal diet without or with 250 mg of TD/kg of feed. Overall, lambs were distributed in four treatment combinations (n = 5) according to a 2 × 2 factorial arrangement under a randomized complete block design (Figure 1): (1) males without TD, (2) males with TD, (3) females without TD, and (4) females with TD. It is worth mentioning that TD was mixed directly into the diet and was offered during the 40-day feeding trial. Feed was offered ad libitum daily, with a 15% feed refusal, and was served at 06:00, 12:00, and 18:00 h. Drinking troughs were refilled simultaneously with fresh, clean water, while the sheep’s health status was visually monitored in the morning and afternoon, with no sick animals detected. Once the feeding trial was completed, lambs were fasted for 12 h and then slaughtered by exsanguination at a slaughterhouse located at the study site.
2.4. Climatic and Physiological Variables
Relative humidity (RH), Ta and temperature–humidity index (THI) were recorded as climatic variables. The first two were measured with a thermohydrometer (Termotraker^®^, Culiacan, Sinaloa, Mexico) that was placed in the center of the experimental area at the height of the animals’ heads. This device was programmed to automatically record both Ta and RH every 20 min during the 40-d feedlot test. Upon completion, the data were downloaded to Excel^®^ to calculate the THI with the following formula [26]: THI = Ta − {(0.31 − [0.31 × RH]) (Ta − 14.4)}. For all climatic variables, average values per day and hour were calculated, and average daily maximum and minimum values were obtained.
Respiratory rate (RR), rectal temperature (RT), and surface thermography in different body regions were assessed at 06:00, 12:00, and 18:00 h on days 1, 10, 20, 30, and 40 of the feedlot trial. The number of intercostal movements per minute was quantified to determine RR, while a digital thermometer (Delta Track, Pleasanton, CA, USA) was inserted into the rectum to record RT. Finally, three thermal images per lamb were taken (front, side, and caudal; Figure 2) with an infrared thermographic camera (Fluke Ti401, Everett, WA, USA). These were downloaded to the computer to record the surface temperatures with the Smart View^®^ software (version 4.4., Fluke, WA, USA) in the following body regions: eye, ear, muzzle, forehead, nostrils, neck, shoulder, loin, belly, rib, flank, rump, leg, fore limb, hind limb, vulva, anal, rectal, perineum and testicles.
2.5. Feedlot Performance
All lambs were weighed individually on days 1 and 40 of the feedlot trial to record initial and final BW. The amount of feed offered and refused in each pen was weighed daily in the morning to calculate DMI. In addition, total weight gain (TWG), daily weight gain (ADG = TWG/fattening days), and feed efficiency (ADG/feed intake) were calculated.
2.6. Carcass Traits, Non-Carcass Components, and Wholesale Cut
All lambs were weighed before the slaughter to record slaughter BW. Once killed, lambs were bled, skinned, and eviscerated to record the individual weights of blood, head, skin, feet, testicles, heart, lung, liver, kidneys, spleen, full and empty gastrointestinal tract (GT), internal fats (omental, mesenteric, and kidney-pelvic-heart [KPH]), and hot carcass (HCW). While KPH fat weight was expressed as a percentage of the HCW, the weights of the other non-carcass components were expressed as a percentage of the empty live weight (EBW = slaughter BW − [full GT − empty GT]). Hot carcass weight was also expressed as a percentage of the EBW to calculate the carcass yield.
Subsequently, the carcasses were placed in a cold room at 4 °C to record cold carcass weight (CCW) and zoometric measurements (i.e., carcass length, thorax depth, chest circumference, and leg length and circumference). Then, carcasses were ribbed between the 12th and 13th to measure LT muscle area with a dot square grid of 64 mm^2^, as well as backfat thickness with a vernier. Finally, the right half carcass was weighed and dissected to measure the wholesale cut yields as described by Avendaño-Reyes et al. [27]. Briefly, the individual weights of neck, shoulder, loin, plain loin, rib, breast, and flank, leg, forequarter, and hindquarter were expressed as a percentage of the half carcass weight to calculate their yields.
2.7. Meat Quality
The carcass pH was recorded at 45 min and 24 h postmortem by inserting a penetration electrode into the loin (12th and 13th rib), connected to a portable pH meter (HI-98140, Hanna Instruments, Woonsocket, RI, USA). Subsequently, the LT muscle was dissected and vacuum-packed for aging during 7 d at a temperature between 0 and 4 °C. This muscle was unpacked and subjected to blooming for 30 min before evaluating the meat quality parameters: color, pH, cooking loss, Warner-Bratzler shear force (WBSF), and water-holding capacity (WHC).
The color was measured in triplicate at different points on the surface of each muscle using a portable colorimeter (CR-400, Konica Minolta Sensing, Inc., Osaka, Japan; D65 illuminator and 10° observer). Lightness (L*), redness (a*), yellowness (b*), Chroma (C*), and hue angle (h*) were color variables recorded after calculating an average of each one per muscle. Subsequently, 5 g of muscle and 25 mL of distilled water were homogenized (liquefied) for one minute, and then a pH meter for liquids was introduced into the mixture to measure pH (HI-2210, Hanna Instruments, Woonsocket, RI, USA). The pH was determined in duplicate to calculate an average for each muscle. For cooking loss and shear force, one steak was weighed, cooked until a core temperature of 71 °C using an electric grill, and finally chilled to room temperature for 20 min. The cooled steak was weighed again, and this weight was subtracted from the steak’s weight before cooking it; the difference in weight was expressed as a percentage of the uncooked steak’s weight to record cooking loss. In addition, the cooled steak was cut to obtain three cubes measuring 1.27 cm per side. These cubes were shared longitudinally with respect to the muscle fibers using a Warner-Bratzler shear force device (Salter 235, Manhattan, KS, USA). Finally, the WHC was determined following a standard methodology [28], which consists of weighing 3 g of meat and suspending it on porous fabric within 50 mL tubes es to be centrifuged and weighed again post-centrifugation. Thus, the WHC of the meat was calculated by expressing its post-centrifugation weight as a percentage of its initial weight. Both WBSF and WHC were measured in triplicate, so averages per muscle of each of them were recorded.
2.8. Statistical Analysis
All data were subjected to analysis of variance using procedures from the SAS statistical package (SAS Institute Inc., Cary, NC, USA; version 9.4). Productive performance, carcass characteristics, non-carcass components, wholesale cut yields, and meat quality were analyzed with the GLM procedure, where the models included fixed effects of block, TD supplementation, gender, and the TD × gender interaction; additionally, animal was included in the models as a random effect. Physiological variables were analyzed with the MIXED procedure using the same model, but the hour of the day was included as a repeated measurement factor over time. In this model, different variance-covariance structures were tested to select the best fit based on the AIC and BIC values closest to zero. Means were compared with the PDIFF option of SAS, declaring differences at p ≤ 0.05.
3. Results
3.1. Environmental Conditions
The average Ta and RH recorded during the 40-d feeding period were 33.6 ± 1.4 °C (25.9–41.8 °C) and 47.41 ± 8.50% (23.8–72.3%), respectively, leading to an average THI of 35.89 ± 1.69 units (28.0–43.55 units). Daytime Ta remained above 30 °C and lasted until 22:00 h, while the THI did not drop to a thermal comfort level (<22.2 units) at any time of day (Figure 3).
3.2. Effects of Thiamine Diphosphate
No interaction between TD, gender, and/or hour of the day (p > 0.05) was detected for RT and RR; likewise, these physiological variables were not affected (p ≥ 0.78) by the supplemental TD alone (Table 2 and Figure 4). Some body surface temperatures were only affected (p < 0.05) by the TD × gender interaction; particularly, TD in lambs, but not in male lambs, increased eye, ear, and forehead temperatures by more than 0.50 °C. Regardless of gender, TD increased (p ≤ 0.05) surface temperatures of neck (+0.45 °C), shoulder (+0.43 °C), loin (+0.38 °C), rump (+0.42 °C), forelimb (+0.44 °C), anal (+0.47 °C), vulva (+0.71 °C), perineum (+0.80 °C), and testes (+1.62 °C), as well as, tended to increase them in rib (p = 0.09; +0.37 °C), flank (p = 0.07; +0.41 °C) and hind limb (p = 0.09; +0.33 °C). Surface temperatures in the rest of the body regions were not affected (p ≥ 0.11) by TD feeding.
In feedlot performance, the TD × gender interaction affected (p = 0.02) only DMI; TD decreased intake by 12% in ewe lambs with no effect in male lambs (Table 3 and Figure 5). Regardless of gender, there were no changes (p ≥ 0.28) in weight gain, feed efficiency, and final BW due to TD.
In carcass characteristics, both TD alone or interacting with gender did not affect (p ≥ 0.15) carcass weight and dressing percentage, LT area, chest circumference, leg length and circumference, back fat thickness, and KPH and omental fat deposition (Table 4). Nevertheless, a 23% reduction (p = 0.03) in mesenteric fat accumulation was observed with TD feeding regardless of gender. In addition, there was a TD × gender interaction effect (p ≤ 0.05) on some carcass zoometric measurements where TD in males decreased carcass length and thorax depth by 4.3 and 4.6%, respectively; conversely, in females, it led to an increase in thorax depth by 3.6% without affecting their carcass length (Figure 6).
The TD × gender interaction did not affect either the non-carcass component percentages (Table 5), wholesale cut yields (Table 6), or meat quality (Table 7). Similarly, supplemental TD alone did not affect (p ≥ 0.14) all these variables.
3.3. Effects of Gender
Male lambs, compared to ewe lambs, recorded 11.5 bpm less (p < 0.01) and had higher (p ≤ 0.04) surface temperature on the nostrils (+0.36 °C), ribs (+0.47 °C) and anal region (+0.39 °C), likewise tended (p = 0.08) to have higher temperature on the loin, belly, rump and forelimbs (Table 2). There was no effect (p ≥ 0.12) of gender on RT and the rest of body surface temperatures. Overall, male lambs had 31, 33, and 25% more (p < 0.01) ADG, TWG, and feed efficiency than ewe lambs, respectively (Table 3). This response resulted in male lambs weighing 3.70 kg more (p < 0.01) than ewe lambs at the end of the fattening trial. In carcass (Table 4), the gender did not affect (p ≥ 0.11) HCW, CCW, LT muscle area, and most morphometric measurements (i.e., chest circumference, leg length and circumference). Ewe lambs recorded 6.8% less (p = 0.04) EBW but higher (p = 0.01) carcass yield (47.66 vs. 49.74%) than male lambs. In addition, higher (p < 0.01) backfat thickness, KPH fat, and mesenteric fat were observed in lambs than in male lambs; females tended (p = 0.10) also to accumulate more omental fat.
On the other hand, no effect of gender (p ≥ 0.14) was observed on the weights (expressed as % of EBW) of blood, skin, lungs, liver, heart, spleen, and empty gastrointestinal tract (Table 5). Male lambs versus ewe lambs presented higher (p ≤ 0.04) head (5.19 vs. 6.10%) and kidney (0.29 vs. 0.32%) weights and also tended to have higher (p = 0.08) feet weights. In wholesale cuts (Table 6), the shoulder yield was higher (p < 0.01) by 2.29%, while the forequarter and neck yields tended to be higher (p = 0.10) in male lambs than in ewe lambs. Conversely, the rib (p = 0.10), hindquarter (p = 0.10), and plain loin (p = 0.06) yields tended to be higher in ewe lambs. The rest of the cut yields were similar (p ≥ 0.21) between genders. Finally, the pH in male meat was lower (p = 0.04) at 45 min (6.36 vs. 6.49) and higher (p = 0.01) at 24 h postmortem (5.82 vs. 5.94) compared to that recorded in female meat (Table 7). In meat aged for 7 d, the gender only affected the a* and C* values, with a* being higher (p = 0.05) and C* tending to be higher (p = 0.09) in male meat than in female meat. The remaining color variables, as well as the WHC, cooking loss, and shear force, did not vary (p ≥ 0.11) due to gender.
4. Discussion
4.1. Environmental Conditions
Sheep of this study were exposed to a natural hot environment (Ta = ~34 °C), characteristic of the summer season in desert regions. Despite the good adaptation by hair sheep to warm environments, these breeds present imminent signs of thermal discomfort at Ta = 32 °C, and this becomes very noticeable at Ta ≥ 35 °C [29,30], since heat losses by latent means increase from 55% at 24 °C to 82 and 96% at Ta of 32 and 36 °C, respectively [30]. These climatic conditions provided an HS environment for lambs as the average Ta exceeded the upper limit (i.e., 30 °C) of the thermoneutral zone for hair breed sheep [1]. Based on the average THI, lambs experienced extremely severe HS (THI > 25.6 vs. 35.9 units) [26]. It is noteworthy that lambs were continuously exposed throughout the day to this type of HS, since THI fluctuated between 29 (early morning) and 43 units (afternoon). Therefore, HS mitigation strategies are needed in intensive sheep meat production of this area.
4.2. Effects of Thiamine Diphosphate
TD is a metabolic enzyme cofactor that, in addition to promoting glucose production and energy efficiency, has antioxidant and anti-inflammatory action in mammals [5,6]. To date, this compound has been investigated for improving energy efficiency in sheep raised under thermoneutral conditions, while no reports have been found in hot environments. Healthy sheep maintain a core temperature between 38.3 and 39.9 °C in a thermoneutral environment [26], and HS conditions can raise it due to inefficient thermoregulatory ability [3,31]. Here, the fattening lambs, irrespective of gender, experienced mild hyperthermia [32] as their average RT exceeded the upper limit of the normal range by 0.2 °C (39.9 vs. 40.1 °C) [26], and feeding TD did not prevent it. Despite this response, TD-fed lambs exhibited different physiological thermoregulation strategies, given that they dissipated more body heat load through the skin than control lambs. These findings might be related to the fact that TD improves both the thermogenic capacity of subcutaneous fat tissue adipocytes [33] and peripheral vasodilation [34,35].
Supplemental TD in ewe lambs, but not in male lambs, increased body heat losses through the eyes, forehead, and ears, which indicates a gender-dependent effect of TD on head thermoregulation in heat-stressed hair lambs. Male lambs secrete testosterone, which has a vasodilator function in the brain according to studies with murine models [36]. Thiamine, for its part, plays an essential role in maintaining optimal brain function in ruminants by supplying it with glucose and ensuring adequate blood circulation [35,37]. As a result, this glucogenic cofactor aids in improving blood flow, endothelial function, and vasodilatation in the head [35,38], which explains the positive impact of TD on the heat losses across eyes, ears, and forehead in ewe lambs. In male lambs, this beneficial effect of TD was not observed because testosterone likely activated a thermoregulatory mechanism in the head, like that of TD. Thus, control male lambs achieved similar temperatures in those head regions as TD-fed male and female lambs.
Overall, the results partially support the original hypothesis of this research because the thermoregulatory capacity of the lambs was not compromised by TD supplementation. However, no benefit was observed in growth rate, economically important carcass characteristics, or meat quality with the inclusion of this feed additive during the fattening period. In line with this responses, different studies in sheep housed in a thermoneutral environment did not find changes in growth traits and feed efficiency with oral or injected thiamine [10,13,39]. In fattening calves, the dietary inclusion of this glucogenic cofactor also did not improve productive performance [40,41], carcass characteristics, or meat quality [40]. Therefore, the available evidence shows that the effect of TD on energy metabolism in ruminants is not strong enough to promote growth and affect meat quality. In fact, Kalyesubula et al. [10] mention that TD in sheep could function more as a redistributor of body energy than as a provider of energy substrate for muscle mass formation.
Unexpectedly, TD reduced DMI only in ewe lambs without affecting growth rate, feed efficiency, carcass weight, or LT area. Thiamine can increase dietary energy efficiency by functioning as a pyruvate dehydrogenase coenzyme in the glycolysis pathway for the acetyl CoA formation, a substrate that produces ATP energy [6,7]. It is also a cofactor of catabolic enzymes abundant in some organs, muscles, and fatty tissue, which degrade these tissues to release gluconeogenic precursors (i.e., short-chain branched-chain amino acids and non-esterified fatty acids) that promote the uptake of hepatic glycogen and body energy availability [8,39]. Given that TD did not affect fat deposition (except mesenteric fat), organ weights, and wholesale cut yields in the present study, it is speculated that this glucogenic cofactor improved dietary energy efficiency in ewe lambs. Consequently, they met their energy requirements for growth with lower DMI. Similarly, castrated and ewe lambs had a reduction in feed intake when TD was supplemented at doses greater than 100 mg/d/animal [13], while in intact bulls, does not [40].
In contrast to these findings, it had been hypothesized that hair-breed male lambs would have a better productive response to TD supplementation than ewe lambs under the climatic conditions of the present study. There is no clear explanation as to why the opposite happened, considering that the metabolic sexual dimorphism effect was expected to prevail in these hair sheep as their gonads maintain endocrine activity during the hot summer season in arid regions [18,19]. Perhaps the improved dietary energy efficiency observed in TD-fed ewe lambs could be linked to the fact that females were more thermotolerant than males. In this regard, a sexually dimorphic stress response has been reported in Brahman cattle, a bovine breed recognized for its high tolerance to HS, similar to hair sheep [42]. Compared to bulls, Brahman heifers demonstrated greater stress resilience, as they recorded higher circulating cortisol concentrations and pro-inflammatory activity but did not experience immunosuppression. In addition, further research is needed to clarify whether the optimal dose of TD is dependent on gender to exert beneficial effects on dietary energy efficiency in heat-stressed sheep.
4.3. Effects of Gender
The high summer Ta recorded in the present study compromised the thermoregulatory capacity in lambs of both genders, leading them to experience mild hyperthermia as their RT were 0.25 °C above the normal range [1]. Gender did not affect RT, suggesting that this genetic factor does not influence the thermotolerance of hair breed lambs fattened in desert climate. Although the physiological thermoregulation strategy used by these sheep was gender dependent. Female lambs dissipated excess body heat load by making more use of evaporative mechanisms (e.g., higher RR) than sensible losses. In contrast, the opposite occurred in male lambs who favored greater heat radiation across the skin in varies body regions. This finding is consistent with results observed in heat stressed sheep from other breeds, where females required a greater increase in RR than males to maintain normothermia [43]. The difference between gender in the activation of physiological thermoregulation mechanisms may be attributed to several things: (1) sexual dimorphisms in physiological variables; (2) females tend to deposit more subcutaneous fat, which reduces their ability to transfer heat through the skin [44]; and (3) testosterone synthesized by males promotes peripheral vasodilation in response to elevated Ta, facilitating the redistribution of blood flow to peripheral tissues to dissipate heat load through the skin [45].
As expected, given some previous antecedents [46,47,48,49], male lambs showed superior growth and feed efficiency despite the high Ta recorded. It is noteworthy that male lambs weighed 3.7 kg more at slaughter than ewe lambs; however, this advantage was not reflected in carcass weight, and, in fact, they had a 2.1% lower carcass yield. These results could be explained by differences in body composition between genders [50]. So, the difference in slaughter weight in favor of male sheep was due to a greater BW associated with the full gastrointestinal tract, head, and some offal (i.e., testicles, feet, and kidney). Overall, carcass weight and yield results showed that HS reduces the natural efficiency of hair-breed male lambs, but not in ewe lambs, for gaining BW associated with better carcass mass deposition. This finding is not surprising, considering that HS can minimize testicular endocrine function in male lambs, leading to reduction in circulating concentrations of testosterone [51]. This steroidal hormone represents a natural anabolic for male sheep, as it stimulates the deposition of lean tissue and improves growth and carcass weight [52].
Furthermore, ewe lambs deposited markedly more internal and dorsal fat than male lambs, which is consistent with other studies with hair breed sheep under thermoneutral conditions [47,48,49,50]. The elevated lipogenic capacity exhibited by ewes is attributable to their ovarian estrogen production [16]. It is worth noting that steroid hormones play an essential role in the development of secondary sexual characteristics in sheep, which is directly reflected in palpable differences in the body conformation of males and females [52]. Testosterone in male lambs promotes longer bodies with a wider rib cage and more muscular chest and neck, while estrogen in ewe lambs produces wider hips with a narrower chest and a thinner neck [16]. This could explain the results of wholesale cut yields where male lambs had greater yields in forequarter, neck, and shoulder; conversely, female lambs tended to have better yields in hindquarter and plain loin.
Finally, it has been documented that sheep meat quality can vary according to gender, as male lambs reach physiological maturity earlier and have greater muscle glycogen reserves [15]. However, research results on sheep in this regard are still inconsistent under thermoneutral conditions [53,54], and no reports were found on hair breed sheep under HS conditions. Here, the gender modified the pH of the LT muscle in the first 24 h postmortem; particularly, the male muscle had a higher ultimate pH than the female muscle, which was above the normal range (5.4–5.8) [55]. Based on this muscle pH, one might expect a deterioration in meat quality because high ultimate pH is associated with the presence of dark, firm, and dry meat [56]. But the male meat only had a slightly redder tone with no difference in lightness, shear force, WHC, and cooking loss compared to the female meat. In fact, the average values of these quality variables were within the reference range for sheep meat (WBSF < 49 N, L* = 34 to 44, a* > 14, cooking loss = 15 and 24% and WHC ≥ 80%) [55]. Perhaps the male meat quality did not decrease because the evaluation was made on aged meat. Aging sheep meat for 7 d is a commonly used method to simulate shelf life, but it also has a positive impact by improving meat quality attributes in terms of pH, tenderness, WHC, and sensory profile [57]. This response suggests that future experiments could be conducted using unaged meat to elucidate the pure effect of gender on the meat quality of hair breed sheep subjected to HS.
5. Conclusions
Overall, regardless of gender, thiamine diphosphate supplementation did not prevent hyperthermia or improve growth rate, carcass characteristics, or meat quality of hair breed lambs finished in feedlots under heat stress conditions in desert regions. However, this glucogenic cofactor was effective in decreasing feed intake in ewe lambs, but not in male lambs, without compromising weight gain, carcass weight, Longissimus thoracis muscle area, and meat quality. Furthermore, thiamine diphosphate enhanced sensible heat losses in male and female lambs. Therefore, thiamine diphosphate supplementation proved to be a suitable heat stress mitigation strategy in hair ewe lambs, as it increased dietary energy efficiency.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Vicente Pérez R. Macías Cruz U. Avendaño Reyes L. Correa Calderón A. López Baca M.D.l.Á. Lara Rivera A.L. Impacto Del Estrés Por Calor En La Producción de Ovinos de Pelo. Revisión Rev. Mex. Cienc. Pecu.20201120522210.22319/rmcp.v 11i 1.4923 · doi ↗
- 2Macías-Cruz U. Avendaño-Reyes L. Álvarez-Valenzuela F.D. Torrentera-Olivera N.G. Meza-Herrera C. Mellado-Bosque M. Correa-Calderón A. Growth and Carcass Characteristics of Ewe Lambs Treated with Zilpaterol Hydrochloride during Spring and Summer Rev. Mex. Cienc. Pecu.20134112 Available online: https://cienciaspecuarias.inifap.gob.mx/index.php/Pecuarias/article/view/2822/2367(accessed on 10 August 2025)
- 3Macías-Cruz U. Saavedra O.R. Correa-Calderón A. Mellado M. Torrentera N.G. Chay-Canul A. López-Baca M.A. Avendaño-Reyes L. Feedlot Growth, Carcass Characteristics and Meat Quality of Hair Breed Male Lambs Exposed to Seasonal Heat Stress (Winter vs. Summer) in an Arid Climate Meat Sci.202016910820210.1016/j.meatsci.2020.10820232505111 · doi ↗ · pubmed ↗
- 4Nicolás-López P. Macías-Cruz U. Mellado M. Correa-Calderón A. Meza-Herrera C.A. Avendaño-Reyes L. Growth Performance and Changes in Physiological, Metabolic and Hematological Parameters Due to Outdoor Heat Stress in Hair Breed Male Lambs Finished in Feedlot Int. J. Biometeorol.2021651451145910.1007/s 00484-021-02116-x 33742264 · doi ↗ · pubmed ↗
- 5Serra M. Mollace R. Ritorto G. Ussia S. Altomare C. Tavernese A. PreianòM. Palma E. Muscoli C. Mollace V. A Systematic Review of Thiamine Supplementation in Improving Diabetes and Its Related Cardiovascular Dysfunction Int. J. Mol. Sci.202526393210.3390/ijms 2609393240362174 PMC 12072100 · doi ↗ · pubmed ↗
- 6Manzetti S. Zhang J. van der Spoel D. Thiamin Function, Metabolism, Uptake, and Transport Biochemistry 20145382183510.1021/bi 401618 y 24460461 · doi ↗ · pubmed ↗
- 7Hrubša M. Siatka T. NejmanováI. VopršalováM. KrčmováL.K. MatoušováK. JavorskáL. MacákováK. Mercolini L. Remião F. Biological Properties of Vitamins of the B-Complex, Part 1: Vitamins B 1, B 2, B 3, and B 5Nutrients 20221448410.3390/nu 1403048435276844 PMC 8839250 · doi ↗ · pubmed ↗
- 8Mann G. Mora S. Madu G. Adegoke O.A.J. Branched-Chain Amino Acids: Catabolism in Skeletal Muscle and Implications for Muscle and Whole-Body Metabolism Front. Physiol.20211270282610.3389/fphys.2021.70282634354601 PMC 8329528 · doi ↗ · pubmed ↗