Preserved reptile scales retain microscopic features, revealing a new instance of convergent evolution
Calvin A. Riiska, Gordon W. Schuett, Joseph R. Mendelson III, Jennifer M. Rieser

TL;DR
This study shows that preserved reptile scales retain microscopic features, revealing a new example of convergent evolution in viper microstructures.
Contribution
The study confirms a third independent evolution of isotropic microtexture in sidewinding vipers using preserved museum specimens.
Findings
Museum-preserved reptile scales retain microtextures similar to those in shed skins.
Bitis peringueyi shows a new instance of convergent evolution with micro-pits and spacing specific to sidewinding.
This expands sample availability for evolutionary studies using preserved specimens.
Abstract
Small-scale structures on biological surfaces can profoundly impact how animals move, appear and interact with their environments. Such textures may be especially important for limbless reptiles, such as snakes and legless lizards, because their skin serves as the primary interface with the world around them. Here, we examine ventral microstructures of several limbless reptiles, which are hypothesized to be highly specialized to aid locomotion via frictional interactions. Inspired by prior studies that investigated potential links between microtextures, phylogeny, habitat and locomotion—but that were limited by their reliance on shed skins—we characterized the structures present on preserved museum specimens and found that they are quantitatively similar to those found on shed skins. Using this result, we confirmed a previously hypothesized—but untested due to the lack of shed…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
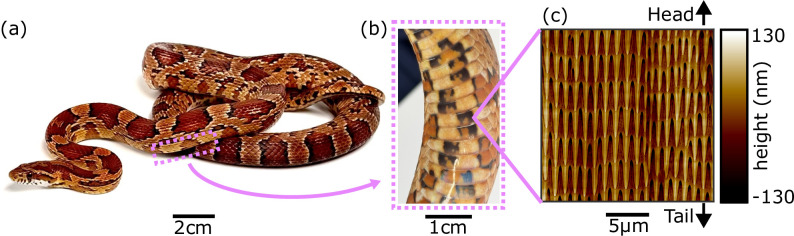 Figure 1
Figure 1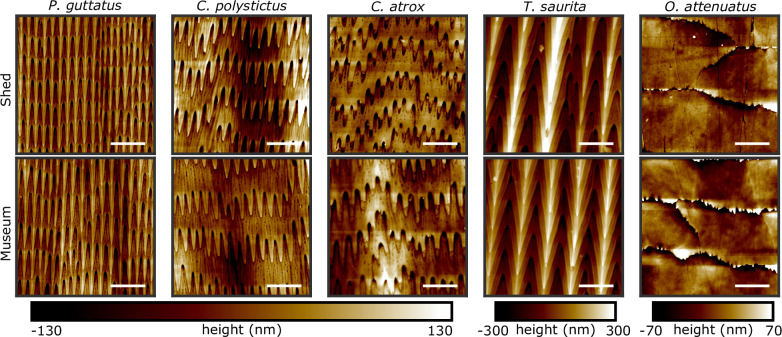 Figure 2
Figure 2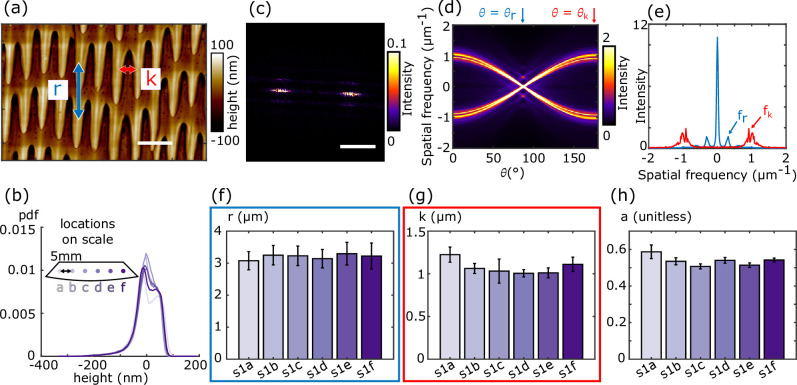 Figure 3
Figure 3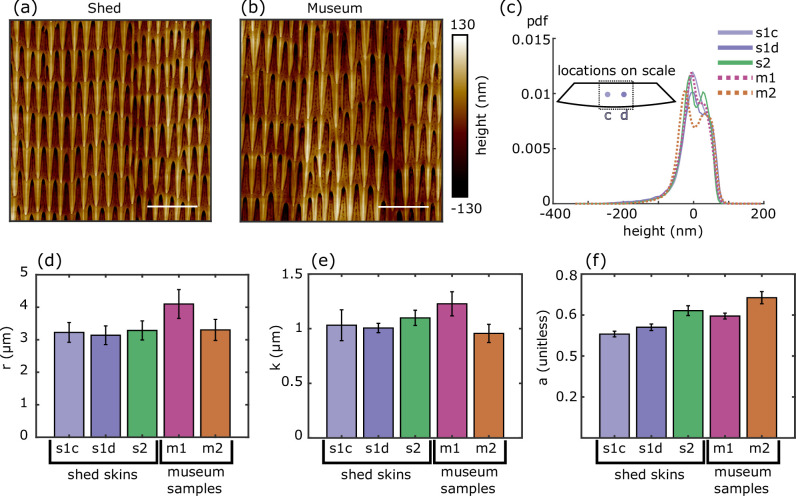 Figure 4
Figure 4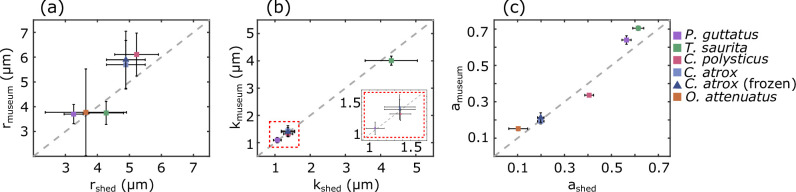 Figure 5
Figure 5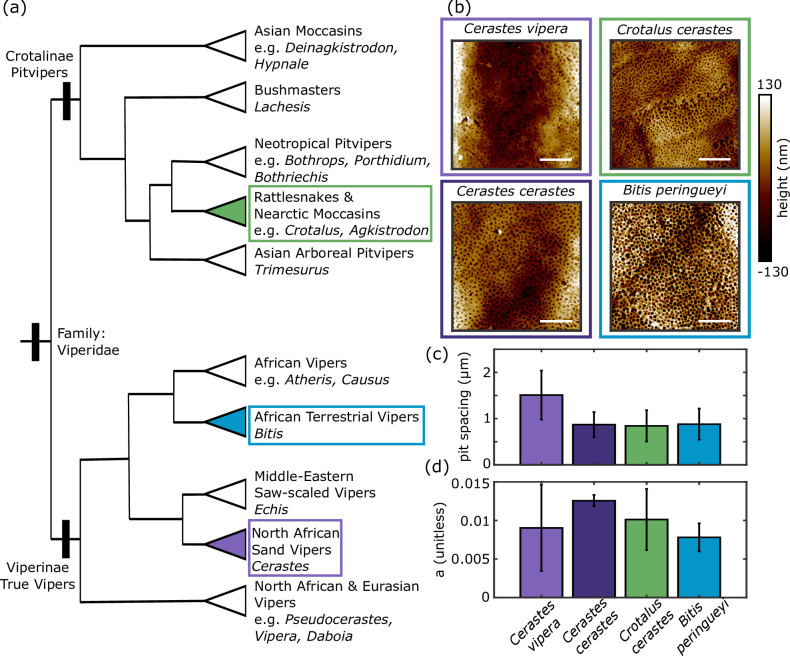 Figure 6
Figure 6- —NSF Physics of Living Systems Student Research Network
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAmphibian and Reptile Biology · Adhesion, Friction, and Surface Interactions · Paleontology and Evolutionary Biology
Introduction
The diverse textures present on animal skins or exoskeletons often play crucial roles in mediating interactions with the environment. Remarkably, microtextures with features on the micrometre or nanometre scale, often requiring the use of sub-diffraction-limit imaging techniques to fully characterize, can influence environmental interactions at macroscopic scales by modulating interactions with light, sound, fluids and other biological matter [1]. For example, reflective microstructures on birds and butterflies can enhance coloration for mating displays [2], reduce visibility for predator avoidance [3,4] and aid in thermoregulation [5]. Sound-absorbing micro-scales on moth wings provide acoustic camouflage from predatory bats [6–8]. Hydrophobic textures help shed water from insect wings [9,10] and, when combined with hydrophilic structures, can enable water harvesting in both desert beetles [11] and rattlesnakes [12] as well as adhesion to air–water interfaces [13]. Micrometre-sized denticles on shark skins help to streamline flows around their bodies [14,15], enhance thrust [16] and protect delicate tissues like gill flaps [17]. Microstructures can also interlock (as in bird feathers [18], enhancing lift during flight) or prevent adhesion (as in carnivorous pitcher plants [19], aiding in prey capture by destabilizing insect footholds).
Microstructures can also directly enhance locomotion through specialized interactions with other surfaces. For example, setae on gecko feet utilize van der Waals forces to allow these animals to adhere to both smooth and rough surfaces [20–22]. Similar structures are found on the feet of flies and beetles, and, when combined with a sticky fluid secretion, these insects can cling to a variety of surfaces as well [23–26]. Other insects, such as ants, stick insects and katydids, have smooth pads with internal fibres that distort around surface features to increase contact-area-dependent forces that are further enhanced by the presence of a fluid secretion [23,24,26–28].
For limbless animals moving across terrestrial substrates, the ventral surface serves as the primary contact with the environment during locomotion. Consequently, it has been hypothesized that specialized ventral microstructures probably evolved to enhance frictional interactions. Many snake species have rearward-pointing micro-spikes (figure 1) that are thought to aid movement through the creation of anisotropic friction—providing low friction for forward sliding, high friction to prevent backward sliding [29], and high lateral friction to reduce sideways slip and enhance forward locomotion [30,31]. Intriguingly, these structures also appear to be linked to habitat; qualitative observations suggest that arboreal snakes have longer spikes than terrestrial species, potentially enhancing friction forces that are crucial for traversing trees, while aquatic and marine species tend to have shorter spikes, perhaps due to the decreased importance of interactions with solid substrates [32]. Fossorial squamates are unique in that the dorsal surfaces also interact with the substrate and may be smoother than the dorsal scales of non-fossorial species [33], possibly to reduce friction during burrowing and subterranean locomotion [34]. Notably, for some desert-dwelling sidewinding vipers, micro-spikes are either greatly reduced or completely absent. Instead, they possess an isotropic array of pits, found to have evolved independently at least twice in distantly related species of vipers [35]. Computational modelling revealed that sidewinding, a periodic pattern of movement in which a planar wave of body curvature is coupled to a vertical wave of body lifting (where lifted segments are moved forward and non-lifted segments are pressed against the substrate) that produces an overall sideways motion [36,37], is enhanced by isotropic friction, which these isotropic structures are hypothesized to produce [35].
The ventral scales of snakes possess microscopic textures. (a) A cornsnake (Pantherophis guttatus). (b) Zoomed-in image of several ventral scales, which serve as the primary animal–substrate interface during locomotion. (c) Microstructures within a 20 µm × 20 µm region of a single ventral scale from the shed skin of a cornsnake, imaged using atomic force microscopy. Many snake species have similar textures, composed of repeating rows of head-to-tail oriented spike-like features, that are thought to enhance frictional interactions during movement.
While these results strongly suggest an important interplay between microstructures in squamate reptiles (as well as in fishes, e.g. [38], though not explored here), movement strategies, habitats and phylogeny, establishing definitive links has been constrained by limited sample availability; previous studies have largely relied on shed skins [35,39–42], which can be difficult to obtain for rare or elusive squamate species.
In contrast, natural history museums house extensive collections of formalin-fixed and ethanol-preserved specimens of all forms on non-avian reptiles, samples of which are made available to researchers upon request for myriad avenues of biological research [43,44], including biometric (e.g. limb sizes, bone shapes and allometry) [45] and histological (e.g. cell arrangements and tissue strength) [46] measurements, information on basic natural history and life habits [47], and DNA extraction [48] for genome sequencing to understand the origin of traits [49] or to recover phylogenetic relationships [50]. While the preservation process is known to have some effects on reptile specimens, e.g. shortened snout–vent lengths, lower body masses [51] and reduced pigmentation [44], the effects of preservation on fine-scale surface microstructures are not known. Some previous studies have included measurements of microstructures on preserved skin samples [32,33,52–54]; however, whether the geometry of preserved structures differs from those in sheds was not explored or discussed. Given the stiffness of snake skin, which is composed of keratin [55–57], we hypothesize that these microstructures may remain intact and comparable to other sample types through the processes of specimen fixation and preservation and thus be useful for analyses using atomic force microscopy (AFM). If true, this would open extensive museum collections to the quantitative analysis of keratin microstructures across diverse species [58], thus alleviating the limitation of sampling based on the availability of shed skins and allowing comparisons with studies that have used other sample origins.
In this study, we used AFM to demonstrate that formalin-fixed, ethanol-preserved museum specimens retain microstructures comparable to those present on shed skins. Unlike the more widely used scanning electron microscopy (SEM) [32,42,59,60], AFM provides nanometre-resolution topographical measurements that enable precise quantification of in-plane geometry, vertical height profiles and spatial anisotropy. SEM also requires sample surfaces to be sputtered with a metal coating such as silver or gold [40–42], and it is unknown how this affects sample topology. Gel-based stereo profilometry and white-light three-dimensional scans provide accurate reconstructions of raw surface topology in a variety of sample types, including live animals [38] (although their use is challenging for some species of squamates (e.g. vipers) that are difficult or dangerous to handle) and samples from taxa that do not shed skin layers (e.g. fishes) [33,38,54]. Despite the versatility of these approaches, a significant limitation is their resolution—they are unable to capture the sub-diffraction limit microstructures of interest in our study. Therefore, we use AFM, as it provides us with the highest resolution three-dimensional surface quantification possible without altering the surface in any way.
Using the AFM-based approach described below, we quantitatively assess both the height distributions and dominant spacings of features within skin microstructures. We first show that textures on shed skins of cornsnakes, Pantherophis guttatus, are similar within an individual (i.e. from distinct regions within a single scale), across individuals (i.e. from sheds collected from two different snakes), as well as for two preserved museum specimens. We then demonstrate that this structural similarity holds for multiple snake species as well as for a legless lizard (Ophisaurus attenuatus) and conclude that microstructures are retained over decades throughout the preservation and storage processes. Finally, we leverage these results by using preserved samples to test the previously framed hypothesis [35], that a new, fourth instance of the pitted microstructure associated with sidewinding may occur in Peringuey’s adder, Bitis peringueyi, a species known to use sidewinding locomotion [61]. If confirmed, this finding would reveal a third independent evolutionary origin of sidewinding behaviour in snakes accompanied by a convergent morphology involving the loss of micro-spikes in favour of an isotropic pitted microstructure. Further, our test provides a concrete example of how museum collections can expand sample availability for studies using AFM to enable hypothesis testing and broader comparative studies of the qualities and evolution of specialized ventral microstructures, ultimately generating unique insights that could establish new links between behaviour, habitat and phylogeny.
Results
We used AFM to image μm sections on snake ventral scales (see Methods). These images revealed that the surface features and detailed morphologies of microstructures on preserved specimens closely resembled those on shed skin samples (figure 2). We examined museum samples and shed samples from four snake species—the colubrids (P. guttatus (cornsnake) and Thamnophis saurita (ribbonsnake)) and the viperids (Crotalus polystictus (Mexican lance-headed rattlesnake) and Crotalus atrox (western diamond-backed rattlesnake)). In each case, the ventral microstructures consisted of periodic rows of raised, rearward-pointing, overlapping spikes, typically 1–5 μm in length. AFM enabled precise height measurements at the nanometre scale, revealing that spikes in P. guttatus, for example, were raised approximately 100 nm above the surface. Secondary features such as ridges and small pits are observed between spikes or between rows of spikes. These microstructures are remarkably well preserved, even in decades-old specimens; our oldest sample, from C. polystictus, was collected and preserved in 1939.
Microstructure topographies for shed skins and preserved samples. AFM scans of 20 x 20 μm regions on shed skins (top row) and museum-preserved samples (bottom row). Structures appear similar for four snake species (P. guttatus, C. polystictus, C. atrox and T. saurita, four left columns) and a glass lizard (O. attenuatus, the right column). Microstructures are retained even for older samples—the C. polystictus sample was preserved in 1939. Scale bars: 5 μm, and the top of each image is towards the head of the animal.
We also observed similar keratinous microstructures on another squamate reptile. In ventral scales from both shed skin and museum samples of the legless slender glass lizard O. attenuatus, the shapes of cell borders and sub-micrometre-scale pits appear similar in both samples. These observations suggest that a wide range of keratin surface structures can be studied using preserved specimens across a broad taxonomic diversity of reptiles, greatly expanding sample availability for comparative and evolutionary studies.
Quantitative similarity within and across shed skins
2.1.
To determine the extent to which ventral microstructures are similar within and across samples, as well as across sample types, we developed a robust quantification method inspired by previous work [35]. All analyses began with AFM-acquired height maps of 60 × 60 μm regions. We started by quantifying features on a shed skin sample (s1) from P. guttatus, chosen because of its common use in locomotion studies (e.g. [62–66]) and characterizing its microstructure may elucidate links to movement. To quantify the geometric arrangement of microstructural features, we first Gaussian-filtered each image to remove depressions and bulges occurring over tens of micrometres caused by irregularities in the sample (see electronic supplementary material, figures S2, S3, S5 and S6). These variations occur over much longer length scales (at least tens of micrometres) than the spiked structures we are interested in characterizing; removing these larger scale variations allows for direct comparisons of the heights and spacings of the spiked features of interest. The resulting mean-subtracted image displays height variations associated with in-plane microstructures that can be compared across sample types. To assess within-sample variability, we imaged six distinct regions (denoted a–f, spaced approximately mm apart) along a single ventral scale from a shed skin sample of P. guttatus (figure 3a). To compare height distributions across images, we used the Jensen–Shannon (JS) divergence, which is bounded between zero and one and measures the distinguishability of two distributions. A JS divergence value of zero indicates that two distributions are identical, while a value of one indicates that the distributions are perfectly distinguishable. We found that the height distributions across these sites were similar (figure 3b), with low JS divergence values (less than or equal to 0.04) between each pair of distributions (see electronic supplementary material, table S2).
Quantification of microscopic features reveals structural similarity across a single scale. (a) A zoomed-in view of microstructures on a P. guttatus shed skin (s1). Two quantities, r and k (the average row and spike spacing, respectively), are used to characterize and compare samples. Scale bar: 2 μm. (b) A third quantity, the relative height variations within a sample, characterized through distributions of mean-subtracted feature heights, provides an additional comparative measurement. Distributions of relative height variations from evenly spaced sites within a single scale (as illustrated by the inset) indicate similarities across sites. (c) Power spectrum of zoomed-out version of (a) (see s1c image in electronic supplementary material, figure S3), with dominant and secondary peaks associated with in-plane structural periodicities along and across spikes. Scale bar: 1 μm−1. (d) Radon transform of (c). The angular direction, θr, where the two primary peaks collapse is identified as the angle with maximal intensity. (e) Slices of the Radon transform used to calculate r and k. Secondary peaks in the dominant direction slice (θr, blue) correspond to the spatial frequency of rows of spikes. Peaks in the slice where the primary peaks collapse furthest apart (θk, red) correspond to the lateral spatial frequency of the spikes. The (f) row spacing r, (g) spike spacing k and (h) anisotropy indices a, are all similar across the six sites in the sample. In (f) and (g), error bars represent the sample variability estimated using the standard deviation (s.d.) of the Gaussian fit to the associated slices at each site (see electronic supplementary material, figure S1). In (h), error bars represent the s.d. of the anisotropy values in each of the four image quadrants.
For in-plane structure, we expanded previous analyses [35] to quantify two key spatial metrics: the spacing between the rows of spikes, , and the lateral spacing between adjacent spikes, (see labels in figure 3a). Starting with our Gaussian-filtered, mean-subtracted height map (electronic supplementary material, figure S3), we first computed the two-dimensional power spectrum (figure 3c). The frequencies associated with the two brightest peaks in the power spectrum correspond to high lateral periodicity (the spike spacing, ), while secondary peaks above and below the primary peaks reflect vertical periodicity (the row spacing, ). The broadness of the primary and secondary peaks as well as the slight rotational offset—caused by imperfections in sample mounting—made extracting dominant peak locations and orientations from these images challenging; therefore, we used a Radon transform, which projects the intensity of a two-dimensional image on to lines of varying orientation that pass through the centre of the image, to identify the directions associated with dominant peaks (see §4.4 for more details).
The Radon transform of the power spectrum (figure 3d) is the brightest for the orientation, , that projects the primary intensity peaks in the power spectrum on to the origin. In the slice at , the secondary peaks above and below the primary power spectrum peaks collapse, creating smaller off-centre peaks that identify the spatial frequency associated with the vertical spacing, , between rows of spikes. The orientation, , with the largest spacing between primary peaks in the Radon transform, corresponds to the spatial frequency associated with the lateral spacing, , between adjacent spikes.
Vertical slices through the Radon transform, taken at and , are shown in figure 3e. Row and spike spatial frequencies and variability were estimated by locally fitting Gaussians to relevant peaks (smaller off-centre peaks for rows and off-centre peaks for spikes; see §4.4 and electronic supplementary material, figure S1, for full details). The peak of each Gaussian fit (i.e. the mean value) represents the average spacing across the sample, and the width of the Gaussian (i.e. the standard deviation (s.d.)) represents the variability in these quantities across the sample. We also report uncertainties associated with the fit quality for each Gaussian, which provides an estimate for how accurately we can measure the sample mean and variability. We note that the fit uncertainties remain small compared with sample variability for all samples studied (electronic supplementary material, table S3). At site s1c, the row spacing is μm, and the spike spacing is μm. This approach allows for systematic comparisons across sites, and across six sites within one sample, variation across sites fell within the sample variability (s.d.s) at each site. This is reflected by the bar graphs in figure 3f,g—for each site, the height of each bar indicates the mean spacing value (given by peak of the Gaussian fit), and the error bar represents the sample variability (given by the s.d. of the Gaussian fit). We conclude from these plots that there is no systematic variation across sites, and that the variation of the mean values across sites is comparable to the variability within a site.
To quantify the directional variability of the microstructure, we measured the anisotropy index, , a quantity introduced in a previous study [35]. The anisotropy index of a region is defined as the JS divergence between vertical slices of the Radon transform (of the power spectrum) separated by . An anisotropy index close to zero indicates a uniform structure with little direction dependence, while larger values indicate higher directionality. In our calculation, we divided each image into four quadrants and calculated the average and s.d. of the anisotropy index over all quadrants. The anisotropy index at site s1c was indicating a large degree of directionality. While anisotropy s.d.s do not always overlap, the anisotropy values are comparable across sites, providing a baseline for structural variability in like samples.
A second shed skin sample (s2) from P. guttatus showed that the height distribution and spacing of structures do not vary between individuals (figure 4c–e). This sample had a row spacing of μm and a spike spacing of μm, nearly identical to s1c. Structures from different individuals were also similarly anisotropic—s2 had a calculated anisotropy of —confirming that microstructural features are consistent between individuals of the same species (figure 4f).
Quantification of structural similarities across samples and sample types. AFM images of P. guttatus microstructures of (a) a shed skin and (b) a preserved museum sample are visually similar. Scale bars: 5 μm. (c) Distributions of height variations from two different individuals each for shed skins (s1 and s2) and museum samples (m1 and m2). The (d) row spacing and (e) spike spacing are quantitatively similar across sample types. The (f) anisotropy indices are slightly more variable but exhibit no systematic trend.
Microstructures are retained on preserved samples
2.2.
To assess similarities and differences across sample types, we compared microstructures from two shed skins and two preserved samples of P. guttatus (figure 4a,b). Distributions of feature heights (figure 4c) were consistent across all samples, with JS divergences less than or equal to 0.04, comparable to within-sample variation. Use of spatial frequency analyses described above showed that preserved specimens retained row- and spike-spacing values similar to those from shed skins (figure 4d,e). While one museum specimen (m1) showed slightly higher average spacings ( μm; μm), these values fell within the range of natural variation. The second museum specimen (m2) closely matched the shed skin measurements ( μm; μm). Given that all measurements of and are within our calculated variability of each other, we find that these values are retained through the preservation process and remain comparable to shed skin samples. While the values of shed skins and museum samples do not fall within a sample s.d. of each other, they do not vary more than our baseline variability established by comparing six sites on the same sample. Differences in , and values can be attributed to natural variation in the samples. It is worth noting that we also examined whether body size might influence structural spacing. Using ventral scale width as a proxy, we ranked specimens from largest to smallest: s1 ( mm), m1 ( mm), s2 ( mm) and m2 ( mm). No trend emerged linking individual size to microstructural spacing.
Extending our analysis beyond P. guttatus, we applied the same quantification technique to preserved samples from three additional snake species—C. polystictus, C. atrox and T. saurita—and a glass lizard, O. attenuatus. In all snake species, preserved samples had row and spike spacings and anisotropy indices consistent with those of shed skins (figure 5). In O. attenuatus, which only showed a dominant periodicity in the head-to-tail direction, both preserved and shed samples exhibited similar cell row dimensions and anisotropy indices, supporting the conclusion that keratin-based microtextures are preserved across diverse reptilian taxa. Electronic supplementary material, table S3 summarizes measurements for each sample studied, including the sample average row spacing with associated sample variability and fit uncertainty , the sample average spike spacing with associated sample variability and fit uncertainty , and the anisotropy index with associated sample variability .
Quantitative comparison of structural measurements across sample types for several species. (a) The preserved versus shed row spacings for four snake species and one lizard species all lie close to the dashed grey y=x line. (b) The preserved versus shed spike spacings also lie close to y=x, indicating similarity of different sample types and multiple species. (c) The anisotropy indices are also similar across sample types (including a frozen sample for C. atrox, blue triangle). Note that O. attenuatus only has a dominant structural frequency in the head-to-tail direction.
A third independent origin of pitted microstructure in sidewinding snakes
2.3.
Statistical ancestral state reconstruction of specialized sidewinding—as opposed to idiosyncratic instances of sometimes poorly manifested sidewinding in unusual or non-natural situations—has independently evolved five times in vipers [61]. These include once in pit vipers (subfamily Crotalinae), in the American sidewinder rattlesnake (Crotalus cerastes) and four times in the Old-World true vipers (subfamily Viperinae), including Echis carinatus, the genus Cerastes (three species), the common ancestor of the genera Pseudocerastes (three species) and Eristocophis (one species), and the common ancestor of a clade comprising the species B. caudalis, B. peringueyi and B. schneideri (the latter species is not considered a sidewinding specialist, thus it is a case of a potential evolutionary reversal per Tingle [61]). Using our result that microstructures are preserved on museum specimens, we analysed a museum-preserved sample of B. peringueyi, a sidewinding true viper native to the Namib Desert of southern Africa [67]. We found that its ventral microstructure closely resembles that of other sidewinding viper species. Previous work [35] with the pit viper (Crotalinae) Crotalus cerastes and the true vipers (Viperinae) Cerastes cerastes and Cerastes vipera identified the distinctive pitted microstructure and the reduced or missing rearward-facing micro-spikes that are associated with specialized sidewinding. However, that study was limited to shed skin samples, and no shed skins were available for B. peringueyi. By leveraging our results from this study and using a museum-preserved specimen, we were able to examine the relatively unrelated B. peringueyi, thus expanding the evolutionary context of sidewinding microstructures in four species of vipers.
Using power spectra and Radon transforms (electronic supplementary material, figure S12), we measured the characteristic spatial frequency of the pitted textures, finding that Crotalus cerastes, two species of Cerastes and our new results for B. peringueyi all show an average pit spacing between 0.9 μm and 1.5 μm with overlapping variability up to μm (figure 6c). This overlap suggests convergence not only in structure type but possibly also in pit spacing. Additionally, all sidewinding species show low anisotropy indices (figure 6d), reflecting the isotropic nature of the pitted texture. These metrics are summarized in electronic supplementary material, table S4. Notably, B. peringueyi conforms to the isotropic pitted pattern, providing another example of convergent evolution in both behaviour and potentially beneficial ventral microstructures.
A preserved B. peringueyi sample reveals a third origin of convergently evolved microstructure in sidewinding vipers. (a) A phylogeny of viperids adapted from [50] shows the three branches represented in our sidewinder microstructure images. It is most parsimonious to conclude three independent origins of these microstructures and sidewinding locomotion than to propose a singular origin in the clade Viperidae with dozens of independent losses. (b) A similar pitted structure appears in each of the four sidewinding species imaged. Scale bars: 5 μm. (c) The spacing between pits in each species is similar, suggesting this structure may have evolved with a characteristic spacing. (d) The microstructure of each sidewinder species is similarly isotropic.
Previous work [35] established that the microstructures associated with specialized sidewinding evolved independently in crotaline Crotalus cerastes and the viperine genus Cerastes. Our results here indicate a third evolutionary origin of the distinctive sidewinding microstructure in B. peringueyi (figure 6a, which was adapted from [50] and [68]). This suggests the parsimonious hypothesis that similar microstructures have evolved once in the clade, including Eristocophis macmahoni and Pseudocerastes spp., and again independently in E. carinatus. In other words, the five independent origins of specialized sidewinding in vipers [61] should be accompanied by five independent origins of sidewinding microstructures. Three origins of such microstructures have been confirmed ([35]; this study), directing future work to investigate Echis, Eristocophis and Pseudocerastes. Collectively, these results suggest strong selective pressure for the performance-related microstructural trait among sidewinding vipers, probably in response to similar environmental challenges of sandy substrates [69]. Nevertheless, sidewinding has evolved independently in desert-dwelling members of the Viperinae and Crotalinae without notable changes in other aspects of body morphology [61,68]. Among the species studied here—three in Viperinae and one in Crotalinae—all exhibit similar pitted structures, while micro-spikes are absent (figure 6a,b). Anecdotal reports on non-specialized sidewinding exist for B. cornuta, and unclear reports of sidewinding-like behaviours are noted in B. arietans and B. gabonica. Bitis gabonica is an exceptionally heavy-bodied snake that is not associated with sandy substrates and is known to primarily use rectilinear locomotion (G.W.S., J.R.M., personal observation), so it is an unlikely candidate for either facultative sidewinding or microstructural adaptations for sidewinding. Yet, it does have notably reduced ventral micro-spikes ([35]; electronic supplementary material, figure S1). Rieser et al. ([35]; electronic supplementary material, figure S1) sampled three species of Echis (but not the known sidewinding species E. carinatus [61]) and found micro-spikes typical of non-sidewinding vipers in E. leucogaster and E. coloratus but an unusual, smooth texture in the non-sidewinding species [61] E. pyramidum. Considered together, the results of this study along with [35] and [61] indicate that more work on both sidewinding behaviours and ventral microstructures in viperines is warranted. Such work should consider morphology, behaviour and environment in an explicitly integrative evolutionary framework, as outlined by Tingle [70].
Discussion and conclusions
In this work, we used AFM to demonstrate that microstructures on the keratin-based skins of several preserved snakes and one lizard are each topographically similar to shed skin samples from the same species. Few studies have systematically quantified the in-plane geometry of snake microstructures; most have provided qualitative observations [42,60] or hand-measured the size of features on the surface [35]. By collecting height profile data and calculating feature spacings, we developed robust analyses that enabled comparisons of ventral microstructures of reptiles from both shed skins and those found on preserved skins.
Our analyses revealed that preserved specimens, including those preserved for over years, retained their fine-scale surface structures. Specifically, we found that surface heights, in-plane feature spacings and anisotropy indices were similar across sample types, with inter-sample variability falling within intra-sample variability. This validation greatly expands the range of specimens available for microstructural analysis beyond the limited supply and availability of shed skins and opens the door for broad studies linking structure to habitat, phylogeny and locomotion strategy.
We leveraged our results to investigate an untested hypothesis [35], which discovered two evolutionarily independent losses of micro-spikes in favour of isotropic pitted microstructures found on three sidewinding viper species. A computational model demonstrated that such isotropic structures may enhance frictional interactions that benefit sidewinding movement, and therefore, the authors predicted similar microstructures on a fourth sidewinding species, B. peringueyi. While we were also unable to obtain a shed skin for this species, our results here indicate that preserved museum specimens can be used instead of shed skins. Indeed, AFM images of a museum specimen revealed that B. peringueyi has the predicted pitted microstructure observed on the other sidewinding vipers. Comparisons of quantified structures demonstrated that all sidewinding species had a similar spacing between pits, suggesting that this structure may have evolved with a characteristic geometry.
We note that although the morphology of snake microstructures was retained in museum samples, the effects of preservation on mechanical properties—e.g. elasticity, friction and wear—of the skin are not known. Though some prior work has measured elasticity [71,72], microscopic friction [29,30,39,73], microstructure wear and durability [72] for shed skins from a few species, how structures relate to mechanical properties is not well understood. Future work should explore these connections further, as well as investigate how chemical fixatives used in preservation may or may not affect mechanical properties. Either way, linking microstructures to mechanics, even if only shed skin samples can be used, would not only advance our understanding of biological surface function but could also inspire the design of engineered materials with tunable anisotropic friction [74,75].
While differences in mechanical properties across sample types remain to be explored, our results show that preserved specimens may be used in place of shed skins to study structural properties of reptile skins. This finding greatly expands opportunities for high-resolution comparative studies across a wide range of species, including rare or elusive taxa. As the environment affects both the body shape [76] and skin thickness [77] of snakes, it is reasonable to hypothesize that the form and function of microstructures can vary as well. For example, dorsal microstructures in snakes often are hydrophobic and show a strong phylogenetic signal [41,42]. In contrast, ventral structures appear more closely tied to locomotion and ecological niche, such as observations of longer spikes on arboreal snakes (which may enhance frictional interactions) and shorter or smoother structures that may reduce drag or debris accumulation for aquatic and fossorial species [32,33,78,79]. Museum collections can provide the phylogenetic diversity needed to gain new insights into drivers of the evolution of structures that are crucial for environmental interactions.
Methods
Sample preservation
4.1.
The preservation process involves injecting the animal with formalin (3.7% formaldehyde), positioning it and wrapping it in formalin-soaked cloth to cross-link proteins, position the specimen in a desired configuration and prevent decomposition [43,80]. Samples are then preserved indefinitely in 70% ethanol, which is relatively non-toxic and prevents microbial and fungal growth [81]. Some specimens are preserved directly in ethanol, which better preserves DNA, though ethanol concentrations must remain high to prevent decay [43]. Frozen samples were collected from freshly deceased individuals and kept on ice for two days, then submerged in water and placed in a freezer so ice would encase the specimen. Shed skins were collected from specimens housed at various locations, including the Rieser laboratory at Emory University and Zoo Atlanta in Atlanta, GA (see electronic supplementary material, table S1), and stored in sealed Ziplock bags at room temperature.
Sample preparation
4.2.
For this study, we examined three different types of snake skin samples: those from shed skins, preserved samples and frozen samples. For shed skins: sample s2 (P. guttatus) and the sample from O. attenuatus were collected from our laboratory population after each animal had naturally shed; sample s1 (P. guttatus) was collected at Zoo Atlanta, Georgia, along with samples from C. polystictus and Crotalus cerastes; samples from C. atrox were obtained from a captive population in the Chiricahua Desert Museum in New Mexico. Sections of two–three ventral scales were cut out of the sheds for imaging.
Preserved samples were obtained from either the Field Museum of Natural History in Chicago, Illinois, or the Amphibian and Reptile Diversity Research Center, University of Texas at Arlington. These samples came from specimens that had been formalin-fixed and preserved in greater than or equal to 70% ethanol. Samples from undamaged sections of freshly dead road-killed adult specimens of C. atrox from Cochise County, Arizona, were formalin-fixed and ethanol-preserved using standard museum protocols; additional samples of skin from those specimens also were frozen for subsequent attempts at AFM imaging. To obtain skin from these samples for imaging, the top layer of skin on individual ventral scales was peeled off the specimens.
Initially, we attempted to image frozen and preserved samples while the skin was still attached to the scale tissue. This approach proved to be difficult because the sample would relax during imaging and the AFM tip would lose contact. This problem could potentially be remedied by letting the samples dry overnight or by imaging them while partially submerged in a bath of water (but with the top surface exposed to air), but we found that these techniques worked inconsistently. The uppermost dermal layer, the stratum corneum, of skin would also occasionally become detached. As a result, we removed this layer and imaged it directly. Some museum samples that we obtained, however, did not have the top layer of skin present. We initially assumed that it was not removable on some samples, but after imaging the top layer of tissue on these samples, no microstructures were found. We concluded that the top layer of skin was absent in these cases; it is common to find the top layers of reptile scales detached from museum specimens and accumulated at the bottom of specimen jars. In order to accurately capture microstructures on museum or frozen samples, the stratum corneum layer of the epidermis must be present. The stratum corneum is the superficial skin layer, and because it naturally comes loose in museum specimens, curators do not consider removal of the layer from a single scale to represent destructive sampling of specimens. Scale layers found already loose in the specimen jar may be used, but care must be taken to allocate a sample to a specific specimen if more than one specimen is housed in the jar.
Before imaging, all skin samples were cleaned using a previously established protocol [35] to remove dirt and debris that may cover microstructures. The cut-out or peeled-off ventral scales were placed in a solution of 5% Micro-90 mixed with deionized water. We then used a Cole Parmer ultrasonic cleaner to sonicate the samples in the solution for 30 min. After sonicating, the samples were rinsed with tap water to remove the Micro-90 solution. The samples were then laid flat on a wire screen and left to air dry in the laboratory for at least 12 h to ensure they were completely dry before imaging. We used two-sided Scotch tape to adhere the scales to glass microscope slides. After this, the samples were ready for imaging. We ensured that the cleaning and drying process did not affect the microstructure itself by imaging samples that were clean without washing. Nevertheless, small amounts of residual debris are expected and do appear in some of the analysed images.
Our study does not capture variations in the structure based on the location on the body (reviewed in [32,52,60]). One study found no variation in surface topography at different sites on the body in snakes [33], comparing samples from different dorsal or ventral sites in individual specimens, but there are clear differences within individuals when comparing, for example in lizards, dorsal versus ventral surfaces as well as differences related to body size within species [54]. Our goals were to compare microstructures in ventral scales among individuals of the same species using samples that were prepared or preserved differently and to compare microstructures on samples taken from adult snakes of comparable size within species; size-related differences in comparing different species are confounded by the fact that species naturally differ in size. Our study did not investigate any potential ontogenetic changes in species, nor any size-related variation in ventral microstructures that may occur within species. Further work that examines size and ontogenetic variation within species may prove interesting in the context of functionality.
A detailed history of chemical methods of fixation and preservation is often not available for all museum specimens. Because fixation in approximately 10% formalin solution and preservation in 70% ethanol are traditional, we assume that our specimens, borrowed from museums, were prepared in this way. Our sampling and information available do not allow us to determine if microstructures persist under all possible methods of specimen preparation (e.g. fixation and preservation in isopropanol). All samples were taken from ventral scales in the middle third of each snake’s body.
Data collection: atomic force microscopy imaging
4.3.
All samples were imaged using a Jupiter XR large-sample AFM from Asylum Research at Oxford Instruments. We used tapping mode imaging with either a 160AC or 240AC integrated silicon nitride probe. Each sample section of 20 20 μm and 60 60 μm was imaged with 512 pixel resolution at 1 and 0.5 Hz speeds, respectively. Snake skin is made of keratin [55] and thus the imaging surfaces were very hard (elastic modulus of 1–10 GPa) [57] and were undamaged by the imaging process, evidenced by taking five–six successive scans over the same region of the sample and observing no visual changes in the surface morphology.
To identify structural variation across the same sample, we imaged the same sample of shed skin from P. guttatus at six locations across the same scale, roughly spaced 5 mm apart laterally. All other images in this study were taken in a central region of a scale. To show the variation between different samples from the same species, we imaged a second shed skin sample from P. guttatus. We then compared structures across sample types by comparing these images with those taken from two different preserved samples of P. guttatus. All images except those comparing variation across the same sample were taken in the central region of a scale. A similar series of images was taken for C. atrox samples including frozen specimens and analysed in electronic supplementary material, figure S4 to show the viability of frozen samples.
Analysis: quantifying the structures
4.4.
Images from the AFM were flattened, meaning an offset was removed from each scan line so that they all have the same average value, and the overall slope of each scan line was removed. This algorithm is built into the Igor-based AFM imaging software provided by Asylum Research. It is not a plane fit, instead acting on each scan line. The order of flattening can be selected, where order one removes a linear fit, order two removes a quadratic and so on. For each image, order one was chosen. Flattening accounts for thermal drift and sudden changes in tip behaviour, both of which can introduce non-physical offsets in height between consecutive scan lines and slopes in the sample. Flattening also corrects for small imperfections in sample mounting, where it may be impossible to get the sample perfectly flat to the nanoscale. Flattened images were taken to be raw images for our purposes. The raw images still had larger scale depressions and bulges occurring over tens of micrometres, a larger scale than the spikes and pits we wanted to characterize. To bring the features of interest into the same plane, we applied a Gaussian filter with a s.d. of 10 pixels to each image and subtracted the filter from the original image. The final step in processing was mean-subtracting each image. (See electronic supplementary material, figures S2, S3, S5 and S6 for the results of filtering images. Distributions of raw heights are shown in electronic supplementary material, figure S7.)
We first compared the distribution of heights in each of the filtered and mean-subtracted images. By calculating the JS divergence between pairs of distributions, we quantified how similar the heights of features and roughness are across images and sample types.
To calculate the periodicity of in-plane surface features, we built on a previous analysis technique [35]. We calculated a power spectrum of the height values in each image to identify dominant spatial frequencies. Using a fast Fourier transform rather than a power spectrum yielded similar results (electronic supplementary material, figure S13). Images may be slightly tilted from their desired orientation, so we calculated a Radon transform with an angular increment of from these power spectra to more easily measure the location of dominant frequencies independent of the axes. A Radon transform projects the intensity of a two-dimensional image on to a single axis of varying orientation passing through the centre of the image. For example, the Radon transform of a vertical line through the origin would have the line collapse to a single point summing all values along the line at , then gradually increase in width until it reproduces the line at and then decreases in width back to a single point at .
In order to accurately capture the spatial frequency of the structure, we imaged large regions that contained at least 15–20 rows of micro-spikes. For sidewinders, 20 20 μm images were deemed to have enough features to be adequate for this analysis. However, 20 20 μm images from non-sidewinding species only captured a few rows of spikes, so 60 μm images with several rows of spikes were used for this analysis. (Power spectra and Radon transforms for each of our samples are shown in electronic supplementary material, figures S8–S12.)
For spiked structures (all those on non-sidewinding snake species), the Radon transform slice with the highest peak has two secondary peaks whose frequency values correspond to the spacing between rows of spikes, , in the image. The dominant peaks in the slice with two dominant peaks furthest apart correspond to the lateral spacing between spikes, (see electronic supplementary material, figure S1a,b). In each case, the data were smoothed to identify the desired peaks and a linear model was fit to the minima on either side of the peak (electronic supplementary material, figure S1c,e). Subtracting the linear model from the slice allows us to fit a Gaussian curve to the peak and measure its width (electronic supplementary material, figure S1d,f). The mean of this distribution corresponds to the spatial frequency, and the s.d. gives a measure of the variation in the spatial frequency across the image. From these values, we are able to calculate the row spacing, , spike spacing, , and the variability associated with each.
Images from sidewinding species produced a much more uniform Radon transform. By taking an average of all slices in the Radon transform, we were able to identify mirrored peaks on either side of 0 Hz corresponding to a singular dominant spatial frequency. We subtracted the average Radon value at 0 Hz from the data and fit a Gaussian distribution to each side. We calculated the spacing between pits and the associated sample variability using the mean and s.d. of these distributions.
We also adapted a previous technique [35] to calculate an anisotropy index, , from each sample. We first divided each image into quadrants. We then calculated a power spectrum and Radon transform for each. From each Radon transform, we took a slice in the dominant direction containing the maximum intensity, and a second slice at . Note that these slices are different from those used to calculate the row and spike spacing. Those slices tended to be approximately 90° apart but were found independently. The JS divergence between the two slices serves as the anisotropy index for that quadrant. Finding the mean and s.d. of the anisotropy indices gives a measure of the anisotropy index and its variability for the whole image.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Riiska CA, Nguyen C, Peleg O, Rieser JM. 2024 The physics of animal behavior: form, function, and interactions. Annu. Rev. Condens. Matter Phys. 15, 325–350. (10.1146/annurev-conmatphys-040821-120442) · doi ↗
- 2Mc Coy DE, Shultz AJ, Vidoudez C, van der Heide E, Dall JE, Trauger SA, Haig D. 2021 Microstructures amplify carotenoid plumage signals in tanagers. Sci. Rep. 11, 8582. (10.1038/s 41598-021-88106-w)33883641 PMC 8060279 · doi ↗ · pubmed ↗
- 3Vukusic P, Sambles JR. 2003 Photonic structures in biology. Nature 424, 852–855. (10.1038/nature 01941)12917700 · doi ↗ · pubmed ↗
- 4Vieira‐Silva A, Evora GB, Freitas AVL, Oliveira PS. 2024 The relevance of flash coloration against avian predation in a Morpho butterfly: a field experiment in a tropical rainforest. Ethology 130, e 13517. (10.1111/eth.13517) · doi ↗
- 5Krishna A, Nie X, Warren AD, Llorente-Bousquets JE, Briscoe AD, Lee J. 2020 Infrared optical and thermal properties of microstructures in butterfly wings. Proc. Natl Acad. Sci. USA 117, 1566–1572. (10.1073/pnas.1906356117)31919285 PMC 6983360 · doi ↗ · pubmed ↗
- 6Shen Z, Neil TR, Robert D, Drinkwater BW, Holderied MW. 2018 Biomechanics of a moth scale at ultrasonic frequencies. Proc. Natl Acad. Sci. USA 115, 12200–12205. (10.1073/pnas.1810025115)30420499 PMC 6275474 · doi ↗ · pubmed ↗
- 7Neil TR, Shen Z, Robert D, Drinkwater BW, Holderied MW. 2020 Moth wings are acoustic metamaterials. Proc. Natl Acad. Sci. USA 117, 31134–31141. (10.1073/pnas.2014531117)33229524 PMC 7733855 · doi ↗ · pubmed ↗
- 8Neil TR, Shen Z, Robert D, Drinkwater BW, Holderied MW. 2022 Moth wings as sound absorber metasurface. Proc. R. Soc. A 478, 20220046. (10.1098/rspa.2022.0046)35756872 PMC 9199070 · doi ↗ · pubmed ↗