Bacteriophage Mu contamination impacts interbacterial competition
Kat Pick, Valeria Tsviklist, Lauren Stadel, Tracy Raivio

TL;DR
A commonly used bacterial strain was found to be contaminated with bacteriophage Mu, which affected competition outcomes in experiments.
Contribution
The study identifies Mu contamination in a bacterial strain and shows its impact on interbacterial competition.
Findings
Mu contamination was found in a Citrobacter rodentium ∆cpxRA mutant strain.
Strains carrying Mu outcompeted Mu-free competitors in interbacterial assays.
The contamination likely occurred during conjugation with a Mu-containing donor strain.
Abstract
Here, we report the identification of bacteriophage Mu contamination in a commonly used Citrobacter rodentium DBS100 ∆cpxRA mutant strain. After re-constructing a new Mu-free ∆cpxRA strain, we independently replicated the results of a recent study by A. Gilliland, C. Gavino, S. Gruenheid, and T. Raivio (Infect Immun 90:e00314-22, 2022, https://doi.org/10.1128/iai.00314-22). The only result from Gilliland et al. that was impacted by the presence of Mu was the outcome of interbacterial competition assays with the ∆cpxRA strain, as strains carrying Mu consistently outcompeted susceptible Mu-free competitors. These results are important for the field, as the contaminated DBS100 ∆cpxRA mutant strain has been used in six different studies. We believe that the Mu contamination occurred during the construction of the ∆cpxRA allele, during the conjugation of DBS100 with a popular Mu-containing…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
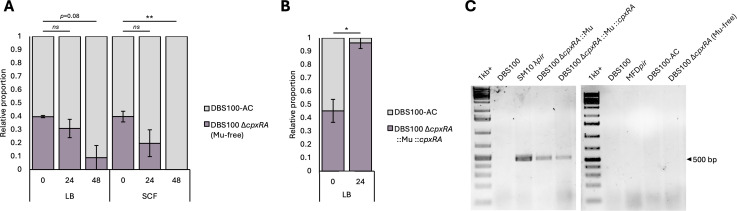 Fig 1
Fig 1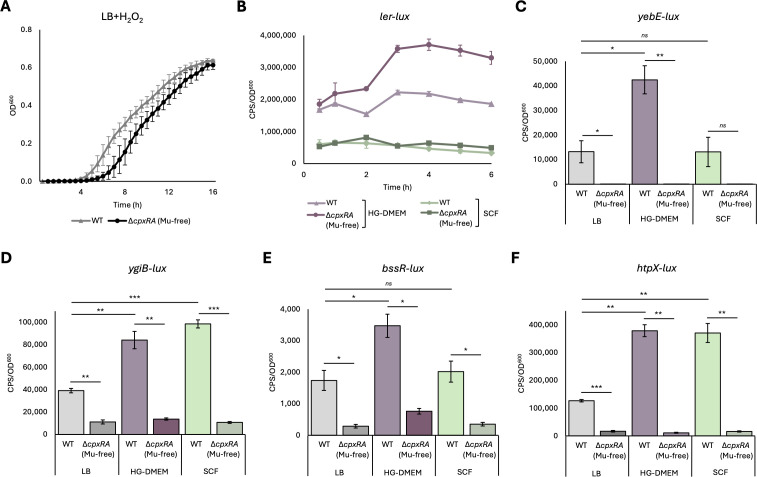 Fig 2
Fig 2| Name | Genotype | Source | Notes |
|---|---|---|---|
| DBS100 | ( | ||
| SM10 λ | ( | ||
| ∆ | DBS100 ∆ | ( | ::Mu |
| ∆ | DBS100 ∆ | ( | ::Mu |
| MFD | MG1655 RP4-2-Tc::[∆Mu1:: | ( | |
| VT298 | DBS100 ∆ | This study | Mu-free |
| DBS100-AC | DBS100 | ( | |
| AG90 | DBS100 (pNLP10 | ( | |
| VT359 | VT298 (pNLP10 | This study | |
| AG31 | DBS100 (pNLP10 | ( | |
| VT360 | VT298 (pNLP10 | This study | |
| AG32 | DBS100 (pNLP10 | ( | |
| VT361 | VT298 (pNLP10 | This study | |
| AG38 | DBS100 (pNLP10 | ( | |
| VT362 | VT298 (pNLP10 | This study | |
| AG61 | DBS100 (pNLP10 | ( | |
| VT363 | VT298 (pNLP10 | This study |
- —Canadian Institutes of Health Researchhttp://dx.doi.org/10.13039/501100000024
- —Natural Sciences and Engineering Research Council of Canadahttp://dx.doi.org/10.13039/501100000038
- —Natural Sciences and Engineering Research Council of Canadahttp://dx.doi.org/10.13039/501100000038
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Bacterial Genetics and Biotechnology · Vibrio bacteria research studies
MATTERS ARISING
The Cpx envelope stress response is a conserved two-component system comprised of the sensor histidine kinase CpxA and response regulator CpxR which together monitor and maintain the integrity of the bacterial envelope (1). A recent article by Gilliland et al. reported that Citrobacter rodentium DBS100 ∆cpxRA mutants have a competitive advantage over WT DBS100 in LB (Fig. 7B) (2). We were puzzled by this result, as C. rodentium ∆cpxRA mutants have been previously demonstrated to have colonization, virulence, and fitness defects (3–7), and Cpx mutants of related species typically have competitive defects (8, 9). Hypothesizing that some type of off-target mutation may have occurred in the ∆cpxRA strain, we performed whole-genome sequencing of the WT and ∆cpxRA strains. Illumina sequencing was performed by SeqCenter (Pittsburgh, PA, USA) using the Illumina DNA Prep Kit and IDT 10 bp UDI indices and sequenced on an Illumina Nextseq 2000 producing 2 × 151 bp reads. bcl-convert (v3.9.3) (Illumina) was used to demultiplex, trim adapters, and check quality. Trimmed (10) WT reads were first compared against the NCBI reference genome (accession no. CP038008.1) using breseq (11) to identify any laboratory-specific mutations that may have arisen in our stock. These mutations were then applied to the NCBI reference genome using gdtools APPLY (11) to obtain a complete reference genome for comparison against the ∆cpxRA strain. Trimmed (10) ∆cpxRA reads were then compared against this new reference genome using breseq (11), resulting in six predicted mutations.
Besides the expected ∆cpxRA mutation, we identified two missense mutations (N857D and T828P) in E2R62_02460 (autotransporter outer membrane beta-barrel domain-containing protein) and three SNPs in the intergenic region between two pseudogenes (E2R62_20730 and E2R62_20735). While certain autotransporters have been implicated in C. rodentium virulence (12), we did not find any clear links to interbacterial competition in the literature and doubted whether the predicted mutations would confer as strong a competitive advantage as seen by Gilliland et al., leading us to dig deeper into our sequencing results. When reads do not align to the reference genome, breseq outputs them into an unmatched.fastq file. De novo assembly of these unmapped reads using SPAdes (13) revealed a 33 kbp contig not present in WT DBS100. BLASTn (14) searches of this contig identified it as bacteriophage Mu (accession no. AF083977.1). Several prophages have been shown to provide a competitive advantage to their hosts (15, 16), leading us to hypothesize that the presence of Mu in the DBS100 ∆cpxRA strain is responsible for the previously reported competitive advantage (2). We constructed a new Mu-free DBS100 ∆cpxRA strain as described in Gilliland et al., using the Mu-free conjugal donor strain MFDpir (17), and repeated the competition assay from Gilliland et al. The Mu-free DBS100 ∆cpxRA strain now shows a fitness defect in LB (Fig. 1A), indicating that Mu likely provided a competitive advantage to the original ∆cpxRA strain. Interestingly, however, Gilliland et al. found that the ∆cpxRA strain still had a competitive defect in simulated colonic fluid (SCF), suggesting that the fitness benefit afforded by Mu is not significant enough to overcome the stressors associated with growth in SCF and was only relevant during competition in LB.
*Mu contamination of DBS100 ∆cpxRA drives competition outcomes. (A and B) Bacterial competition assays were performed as described in reference 1, using the bacterial strains listed in Table 1. Arcsine-transformed values were used to perform paired t-tests (ns P > 0.05, *P < 0.05, *P < 0.01). (C) Colony PCR of strains listed in Table 1, using primers targeting Mu portal protein (CAACCTGCGTGAGGAAGC, GGAATGCCGCCTCATCAATC).
Tracing the construction of the original ∆cpxRA strain (3), it is likely that it was contaminated with Mu during conjugation with χ7213, which has been reported to occur with other Mu-containing conjugal donors such as SM10 λpir (17). If this is the case, then the ∆cpxRA ::cpxRA strain that was constructed in the same manner (3) should also contain Mu and be unable to complement the competitive phenotype of the original ∆cpxRA strain. The ∆cpxRA ::cpxRA strain strongly outcompetes WT DBS100 (Fig. 1B) and was shown by PCR to be contaminated with Mu (Fig. 1C), confirming that the competitive advantage previously observed by Gilliland et al. is independent of the Cpx response and is Mu-driven. Strains constructed via conjugation with the Mu-free donor strain MFDpir are Mu-free (Fig. 1C).
Assembly of the DBS100 ∆cpxRA ::Mu strain using SPAdes (13) revealed that Mu is integrated into the ptrA pitrilysin gene (locus tag E2R62_10930) between nucleotides 67 and 68. ptrA expression was previously shown to be strongly reduced in DBS100 ∆cpxRA mutants, leading the authors to hypothesize that ptrA may be positively regulated by the Cpx response (6). However, our results suggest that the disruption by ptrA by Mu may instead be responsible. In DBS100, ptrA is the operon leader of two downstream genes involved in DNA repair, recB and recD, whose expression is likely also impacted by the disruption of ptrA by Mu. To confirm whether any other results from Gilliland et al. were impacted by the presence of Mu, we repeated the experiments presented in Fig. 2, 3A through D, and 5A with our Mu-free ∆cpxRA strain. ∆cpxRA phenotypes presented in Fig. 1A, 4, 5B of Gilliland et al. were already verified by complementation in the original article (Fig. S5), and ∆cpxRA phenotypes presented in Fig. 8 were previously verified by complementation elsewhere (3). We successfully replicated the results from Gilliland et al. Fig. 2, 3A through D, and 5A with the new Mu-free ∆cpxRA strains (Fig. 2), indicating that the only phenotype impacted by Mu was the competition assay in LB reported in Fig. 7B of Gilliland et al. We did, however, note that yebE-lux and bssR-lux expression in WT DBS100 were not significantly increased in SCF compared to LB (Fig. 2C and E), as was observed by Gilliland et al. (Fig. 3A, C). Since we used the strains originally constructed by Gilliland et al. for this experiment, this discrepancy is likely explained by minor differences in media composition. Since SCF is prepared by weighing eight different powders, and a fresh batch is prepared for each experiment, there is likely significant batch-to-batch variability.
*Replication of results from Gilliland et al. with Mu-free DBS100 ∆cpxRA. Growth curves (A), and luminescent reporter assays (B through F) were performed as described in reference 2, using the bacterial strains listed in Table 1. Paired t-tests were used to determine statistical significance (ns P > 0.05, *P < 0.05, *P < 0.01).
In summary, we have identified bacteriophage Mu as a contaminant found in C. rodentium DBS100 ∆cpxRA and demonstrated that the presence of Mu can impact bacterial competition outcomes. Mu was likely transferred to DBS100 from the conjugal donor strain χ7213 (3), and we strongly urge readers to instead use the Mu-free conjugal donor strain MFDpir (17). Strains constructed by conjugation with S17-1 λpir, SM10 λpir, or any of their Mu-containing derivatives should be carefully re-examined, as transfer of Mu from these strains into susceptible recipient strains is known to occur (17). Only one ∆cpxRA phenotype reported by Gilliland et al. was impacted by the presence of Mu and does not significantly change the overall conclusions of their study.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Raivio TL. 2014. Everything old is new again: an update on current research on the Cpx envelope stress response. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 1843:1529–1541. doi:10.1016/j.bbamcr.2013.10.01824184210 · doi ↗ · pubmed ↗
- 2Gilliland A, Gavino C, Gruenheid S, Raivio T. 2022. Simulated colonic fluid replicates the in vivo growth capabilities of Citrobacter rodentium cpx RA mutants and uncovers additive effects of Cpx-regulated genes on fitness. Infect Immun 90:e 0031422. doi:10.1128/iai.00314-2236000875 PMC 9476912 · doi ↗ · pubmed ↗
- 3Thomassin JL, Giannakopoulou N, Zhu L, Gross J, Salmon K, Leclerc JM, Daigle F, Le Moual H, Gruenheid S. 2015. The Cpx RA two-component system is essential for Citrobacter rodentium virulence. Infect Immun 83:1919–1928. doi:10.1128/IAI.00194-1525712925 PMC 4399039 · doi ↗ · pubmed ↗
- 4Thomassin JL, Leclerc JM, Giannakopoulou N, Zhu L, Salmon K, Portt A, Daigle F, Le Moual H, Gruenheid S. 2017. Systematic analysis of two-component systems in Citrobacter rodentium reveals positive and negative roles in virulence. Infect Immun 85:e 00654-16. doi:10.1128/IAI.00654-1627872242 PMC 5278178 · doi ↗ · pubmed ↗
- 5Giannakopoulou N, Mendis N, Zhu L, Gruenheid S, Faucher SP, Le Moual H. 2018. The virulence effect of Cpx RA in Citrobacter rodentium is independent of the auxiliary proteins Nlp E and Cpx P. Front Cell Infect Microbiol 8:320. doi:10.3389/fcimb.2018.0032030280092 PMC 6153362 · doi ↗ · pubmed ↗
- 6Vogt SL, Scholz R, Peng Y, Guest RL, Scott NE, Woodward SE, Foster LJ, Raivio TL, Finlay BB. 2019. Characterization of the Citrobacter rodentium Cpx regulon and its role in host infection. Mol Microbiol 111:700–716. doi:10.1111/mmi.1418230536519 · doi ↗ · pubmed ↗
- 7Woodward SE, Neufeld LMP, Peña-Díaz J, Feng W, Serapio-Palacios A, Tarrant I, Deng W, Finlay BB. 2024. Both pathogen and host dynamically adapt p H responses along the intestinal tract during enteric bacterial infection. P Lo S Biol 22:e 3002761. doi:10.1371/journal.pbio.300276139146372 PMC 11349234 · doi ↗ · pubmed ↗
- 8Lasaro M, Liu Z, Bishar R, Kelly K, Chattopadhyay S, Paul S, Sokurenko E, Zhu J, Goulian M. 2014. Escherichia coli isolate for studying colonization of the mouse intestine and its application to two-component signaling knockouts. J Bacteriol 196:1723–1732. doi:10.1128/JB.01296-1324563035 PMC 3993324 · doi ↗ · pubmed ↗