“Two scoops, please!”: Twin cryo-EM structures of an Arabidopsis thaliana DNA methyltransferase
Jan Wilhelm Hübbers

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
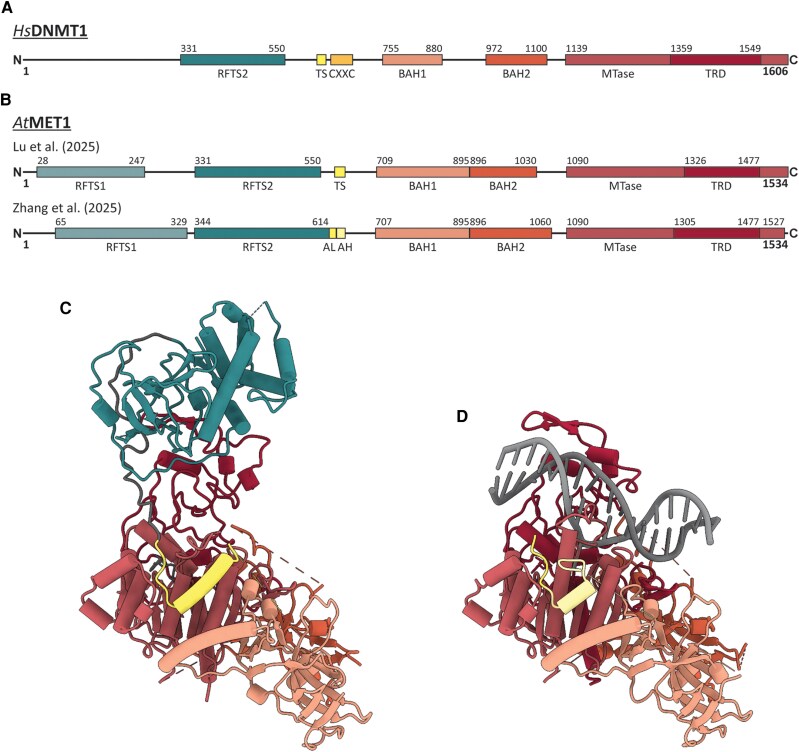 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant nutrient uptake and metabolism · Photosynthetic Processes and Mechanisms · Plant Genetic and Mutation Studies
If transcriptional enzymes such as RNA polymerases are vehicles on DNA roads, cytosine methylation is like placing cones on certain lanes: it slows or reroutes traffic rather than shutting the highway entirely. In this simplification, methyltransferase enzymes are the construction crews that place the methylation cones. Where and how this happens matters. DNA methylation marks promoters, long terminal repeats, transposable elements, and gene bodies, shaping expression and keeping transposable elements in check (Cokus et al. 2008).
The MET1 enzyme (UniProt P34881) maintains CG methylation in the model plant species Arabidopsis (A. thaliana). It belongs to the DNA methyltransferase (DNMT) family of enzymes that is largely conserved across eukaryotes (Schmitz et al. 2019). DNMTs have a characteristic domain architecture comprising a C-terminal methyltransferase (MTase) domain and regulatory N-terminal domains. For example, for human DNMT1 (HsDNMT1; UniProt P26358), these include an replication-foci-targeting sequence (RFTS), a CXXC zinc-finger, and 2 Bromo Adjacent Homology (BAH) domains (Zhang et al. 2015; Fig. A). The MTase domain of HsDNMT1 has an intrinsic substrate preference for hemi-methylated CG, while the RFTS domain autoinhibits DNMT1 in a DNA-competitive manner. This inhibition can be relieved via interactions involving histone H3. Unlike human DNMT1, A. thaliana MET1 has 2 N-terminal RFTS-like regions but lacks a canonical CXXC domain (Fig. B).
In new work, Jiuwei Lu and colleagues (Lu et al. 2025) and Zhihui Zhang and colleagues (Zhang et al. 2025) report 2 cryogenic electron microscopy (cryo-EM) structures of A. thaliana MET1. The first structure, SAH-bound AtMET1 (often termed “apoMET1” in the sense of lacking substrate), was solved at overall resolutions of 3.13 Å (Lu et al. 2025; Fig. C) and 2.92 Å (Zhang et al. 2025). In both apoMET1 structures, the N-terminal RFTS1 domain lacked density, probably due to its flexibility, and was not incorporated in the final model. The second structure, AtMET_ΔRFTS_-hmCG, was deprived of its autoinhibitory RFTS2 domain to enable complexing with hemi-methylated CG and was solved at resolutions of 2.90 Å (Lu et al. 2025) and 2.34 Å (Zhang et al. 2025; Fig. D).
Both studies show that RFTS2 directly associates with the target-recognition subdomain (TRD) of the AtMET1 MTase domain (Fig. B and C). The TRD helps grip DNA and position a flipped CpG cytosine so that the catalytic site can transfer a methyl group from SAM to the C5 position of the target cytosine in hmCG. By binding the TRD, RFTS2 blocks substrate recognition and autoinhibits MET1. The interface residues mediating this contact are conserved in plants, and targeted mutagenesis of several of them increases MET1 activity. Notably, the 2 groups altered different interface residues yet observed similarly increased AtMET1 methylation activities (Lu: P361G or S401A/E1421A; Zhang: Y363A or N1430A/T1431A).
Structural alignment of AtMET1 with HsDNMT1 reveals high overall similarity between the 2 enzymes. However, AtMET1 lacks HsDNMT1's CXXC domain and its downstream inhibitory linker. In line with these architectural differences, the buried surface area at the RFTS–TRD interface is smaller in AtMET1 than in HsDNMT1. Lu and colleagues found that this difference affects the catalytic activity of both enzymes: whereas AtMET1 had a higher catalytic activity at 4 °C, HsDNMT1 was more active at 30 °C.
Both groups also report cryo-EM structures of AtMET_ΔRFTS_ bound to its substrate hmCG DNA. Upon relief of RFTS2-mediated autoinhibition, the same regulatory helix—termed the toggle-switch (TS) by Lu and colleagues and the activating helix (AH) by Zhang and colleagues (Fig. B)—undergoes a pronounced conformational change that pushes the catalytic loop in the MTase domain toward the target cytosine and sets up the active site for methyl transfer.
Altogether, the similarity of these reports is a testament to the robustness of cryo-EM and current structural biology approaches. Reproducibility remains one of the highest quality hallmarks in science. In this light, it is also worth noting that a third report on the structure of AtMET1 was recently published, further supporting the discoveries highlighted here (Kikuchi et al. 2025).
Recent related articles in The Plant Cell
Jiang et al. (2025) proposed a mechanism to ensure the development of precisely on megaspore mother cell in A. thaliana that relies on a balanced amount of methylated and demethylated DNA. Xin et al. (2024) reported cryo-EM structures of Alternative complex III that is essential for phototrophy in the bacterium Chloroflexus aurantiacus. Yao et al. (2024) showed that a 26S proteasome subunit degrades AtMET1 to boost the expression of TERMINAL FLOWER1, maintaining indeterminate growth in the inflorescence meristem. Cao et al. (2023) presented DNA methylomes from nearly 100 accessions of rice to unravel the epigenetic basis of crop domestication and de-domestication.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cao S, Chen K, Lu K, Chen S, Zhang X, Shen C, Zhu S, Niu Y, Fan L, Chen ZJ, et al Asymmetric variation in DNA methylation during domestication and de-domestication of rice. Plant Cell. 2023:35(9):3429–3443. 10.1093/plcell/koad 16037279583 PMC 10473196 · doi ↗ · pubmed ↗
- 2Cokus SJ, Feng S, Zhang X, Chen Z, Merriman B, Haudenschild CD, Pradhan S, Nelson SF, Pellegrini M, Jacobsen SE. Shotgun bisulphite sequencing of the Arabidopsis genome reveals DNA methylation patterning. Nature. 2008:452(7184):215–219. 10.1038/nature 0674518278030 PMC 2377394 · doi ↗ · pubmed ↗
- 3Jiang T, Li L, Hu Q, Kuang X, Zhang L, Zeng W, Miki D, Zheng B. The DNA methylation-demethylation balance prevents development of multiple megaspore mother cells in Arabidopsis. Plant Cell. 2025:37(2):koaf 023. 10.1093/plcell/koaf 02339899470 PMC 11827614 · doi ↗ · pubmed ↗
- 4Kikuchi A, Nishiyama A, Chiba Y, Nakanishi M, To TK, Arita K. Cryo-EM reveals evolutionarily conserved and distinct structural features of plant CG maintenance methyltransferase MET 1. Nat Commun. 2025:16(1):8524. 10.1038/s 41467-025-63765-941006297 PMC 12474915 · doi ↗ · pubmed ↗
- 5Lu J, Chen X, Fang J, Li D, Le H, Zhong X, Song J. Structure and autoinhibitory regulation of MET 1 in the maintenance of plant CG methylation. Plant Cell. 2025; 10.1093/plcell/koaf 24641082549 · doi ↗ · pubmed ↗
- 6Schmitz RJ, Lewis ZA, Goll MG. DNA methylation: shared and divergent features across eukaryotes. Trends Genet. 2019:35(11):818–827. 10.1016/j.tig.2019.07.00731399242 PMC 6825889 · doi ↗ · pubmed ↗
- 7Xin J, Min Z, Yu L, Yuan X, Liu A, Wu W, Zhang X, He H, Wu J, Xin Y, et al Cryo-EM structure of HQNO-bound alternative complex III from the anoxygenic phototrophic bacterium Chloroflexus aurantiacus. Plant Cell. 2024:36(10):4212–4233. 10.1093/plcell/koae 02938299372 PMC 11635291 · doi ↗ · pubmed ↗
- 8Yao WJ, Wang YP, Peng J, Yin PP, Gao H, Xu L, Laux T, Zhang XS, Su YH. The RPT 2a-MET 1 axis regulates TERMINAL FLOWER 1 to control inflorescence meristem indeterminacy in Arabidopsis. Plant Cell. 2024:36(5):1718–1735. 10.1093/plcell/koad 24937795677 PMC 11062425 · doi ↗ · pubmed ↗