Two to tango: DTT1 regulates barley tapetum transition as part of a paired key
Julie Robinson

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
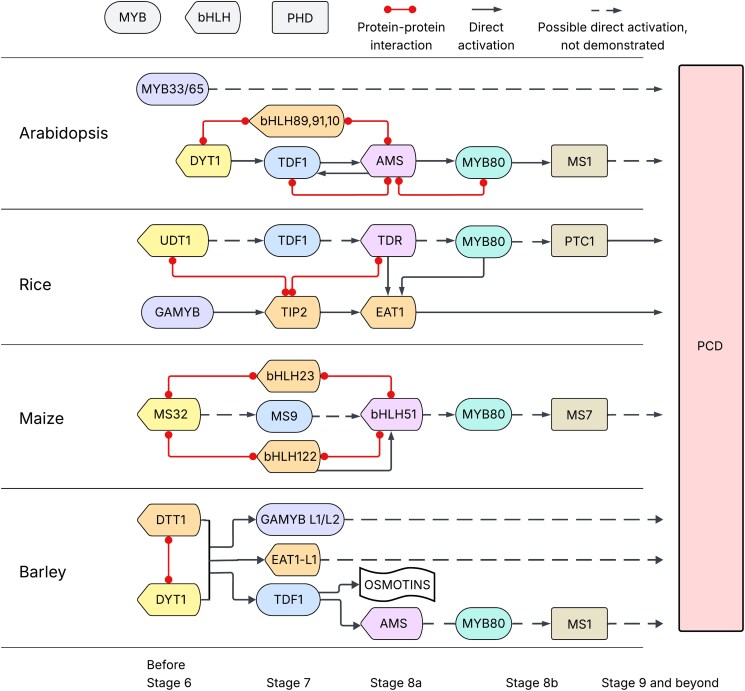 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLegume Nitrogen Fixing Symbiosis
As with the formation of all organs, anther development requires the precise coordination of multiple components to ensure faithful cell differentiation and the formation of a functional organ. In the case of reproductive organs, this is of the utmost importance for the survival of the organism on an evolutionary scale.
The sequential morphological changes that comprise anther development have been well characterized in model species; however, many questions still remain regarding the molecular pathways behind these changes and how well conserved they are among different species, particularly between monocots and dicots. In new work, Miaoyuan Hua and colleagues (Hua et al. 2025) identify DEFECTIVE TAPETUM TRANSITION1 (DTT1) as a protein with a key role in anther development in barley and characterize its function. DTT1 is an ortholog of TDR-INTERACTING PROTEIN2 (TIP2), a basic helix-loop-helix (bHLH) protein in rice that has been well characterized for its important role in anther development (Fu et al. 2014).
First, Hua and colleagues examined the expression pattern of DTT1 in barley and determined that it may be involved in anther development, just like its rice ortholog. Mutating dtt1 indeed resulted in male sterility, cementing its role as a key player in anther formation. Taking a closer look at what caused this sterility, the researchers found that mutant anthers failed to undergo the transition from stage 6 to stage 7 of the 14-stage process. During this transition, the tapetum, a cell layer that provides nutrients to developing pollen mother cells, usually undergoes differentiation that allows it to go on to regulate meiosis within the pollen mother cells. If differentiation fails at this stage, then the pollen does not develop properly and anther development becomes arrested, leading to male sterility.
To better understand the mechanism by which a lack of DTT1 results in failure to transition from stage 6 to stage 7 of anther development, the researchers performed differential gene expression analysis of RNA-sequencing data to identify potential targets of DTT1. Since the MYB transcription factor TDF1 is known to act downstream of DTT1, they eliminated genes that also show differential expression upon tdf1 mutation. The resulting 225 genes showed enrichment for tapetum development and metabolic processes. Among the identified genes were the bHLH transcription factors AMS and EAT1 as well as other MYB transcription factors.
bHLH proteins typically bind DNA as homodimers (two of the same protein) or heterodimers (two different proteins) via their basic regions (Chapman-Smith and Whitelaw 2006). Hua and colleagues investigated whether DTT1 forms homodimers, heterodimers, or both by performing protein interaction assays. They found that DTT1 did not homodimerize when expressed under the AtUbi10 promoter, but did heterodimerize with the bHLH transcription factor DYSFUNCTIONAL TAPETUM1 (DYT1), which is known to be a key player in anther development. A yeast two-hybrid assay showed that formation of this heterodimer was mediated by a highly conserved IKL motif within the ACT-like/bHLH protein interaction and function (BIF) domain of DTT1. However, knowing that most bHLH proteins form homodimers and that DTT1 is a putative paralog of other bHLH proteins that do so, Hua and colleagues revisited their assays with the CaMV35S promoter, which overexpresses genes at a higher level than AtUbi10. Under CaMV35S overexpression, the researchers detected DTT1 homodimers. It must consequently be noted that, when overexpressing proteins for interaction assays, one should consider the stoichiometry of these proteins under physiological conditions. Protein truncation assays showed that DTT1–DYT1 heterodimerization occurs through interaction between their bHLH domains and their ACT-like/BIF domains with themselves and with each other, suggesting a role for the ACT-like/BIF domain in binding partner selection.
In mammals, E-box–like motifs serve as binding sites for transcription factors that regulate gene expression under hypoxic conditions (Kimura et al. 2001). In this work, Hua and colleagues show that the DTT1–DYT1 heterodimer binds both canonical E-box motifs and E-box–like motifs. The authors hypothesize that the DTT1–DYT1 complex may operate under hypoxic conditions, such as those found deep within the tapetum, noting that induction of hypoxia in maize has been shown to stimulate germ cell differentiation in germline-defective mutants (Kelliher and Walbot 2012). Altogether, Hua and colleagues show in this work that the DTT1–DYT1 heterodimer functions as a paired key to regulate the transition from stage 6 to stage 7 of anther development, during which this complex directly targets several key transcription factors. The identification and characterization of DTT1 in barley gives us more information about the evolution of anther development in monocots (Fig.).
Recent related articles in The Plant Cell
Tao et al. (2024) identified a regulatory cascade for starch biosynthesis during pollen development in rice. Chen et al. (2024) characterized an epigenetic mechanism involving polycomb repressive complexes by which stamen development is regulated in Arabidopsis. Chen et al. (2023) identified and characterized a protein that regulates homeostasis of uridine diphosphate sugars in rice in the context of pollen wall development.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chapman-Smith A, Whitelaw ML. Novel DNA binding by a basic helix-loop-helix protein. J Biol Chem. 2006:281(18):12535–12545. 10.1074/jbc.M 51214520016520375 · doi ↗ · pubmed ↗
- 2Chen H, Zhang S, Li R, Peng G, Chen W, Rautengarten C, Liu M, Zhu L, Xiao Y, Song F, et al BOTRYOID POLLEN 1 regulates ROS-triggered PCD and pollen wall development by controlling UDP-sugar homeostasis in rice. Plant Cell. 2023:35(9):3522–3543. 10.1093/plcell/koad 18137352123 PMC 10473207 · doi ↗ · pubmed ↗
- 3Chen Z, Lu J, Li X, Jiang D, Li Z. EMBRYONIC FLOWER 1 regulates male reproduction by repressing the jasmonate pathway downstream transcription factor MYB 26. Plant Cell. 2024:37(1):koae 287. 10.1093/plcell/koae 28739437257 PMC 11663553 · doi ↗ · pubmed ↗
- 4Fu Z, Yu J, Cheng X, Zong X, Xu J, Chen M, Li Z, Zhang D, Liang W. The rice basic helix-loop-helix transcription factor TDR INTERACTING PROTEIN 2 is a central switch in early anther development. Plant Cell. 2014:26(4):1512–1524. 10.1105/tpc.114.12374524755456 PMC 4036568 · doi ↗ · pubmed ↗
- 5Hua M, Yin W, Tidy AC, Fernandez Gomez J, Li H, Shi S, Xing G, Zong J, Wilson ZA. The b HLH transcription factor DTT 1 is part of a paired key to unlock the tapetum transition in barley anther development. Plant Cell. 2025:37(10):koaf 230. 10.1093/plcell/koaf 23041026905 PMC 12510322 · doi ↗ · pubmed ↗
- 6Kelliher T, Walbot V. Hypoxia triggers meiotic fate acquisition in maize. Science. 2012:337(6092):345–348. 10.1126/science.122008022822150 PMC 4101383 · doi ↗ · pubmed ↗
- 7Kimura H, Weisz A, Ogura T, Hitomi Y, Kurashima Y, Hashimoto K, D'Acquisto F, Makuuchi M, Esumi H. Identification of hypoxia-inducible factor 1 ancillary sequence and its function in vascular endothelial growth factor gene induction by hypoxia and nitric oxide. J Biol Chem. 2001:276(3):2292–2298. 10.1074/jbc.M 00839820011056166 · doi ↗ · pubmed ↗
- 8Tao J, Kong W, Luo W, Wang L, Dai X, Lin X, Dong H, Yang X, Mo B, Chen X, et al The rice micro RNA 159-SPOROCYTELESS EAR 2 module regulates starch biosynthesis during pollen development and maintains male fertility. Plant Cell. 2024:37(1):koae 324. 10.1093/plcell/koae 324PMC 1168408439665752 · doi ↗ · pubmed ↗