New species of Aspergillus in sections Cavernicolarum and Nigri from terrestrial ecosystems of China (Eurotiales, Aspergillaceae)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1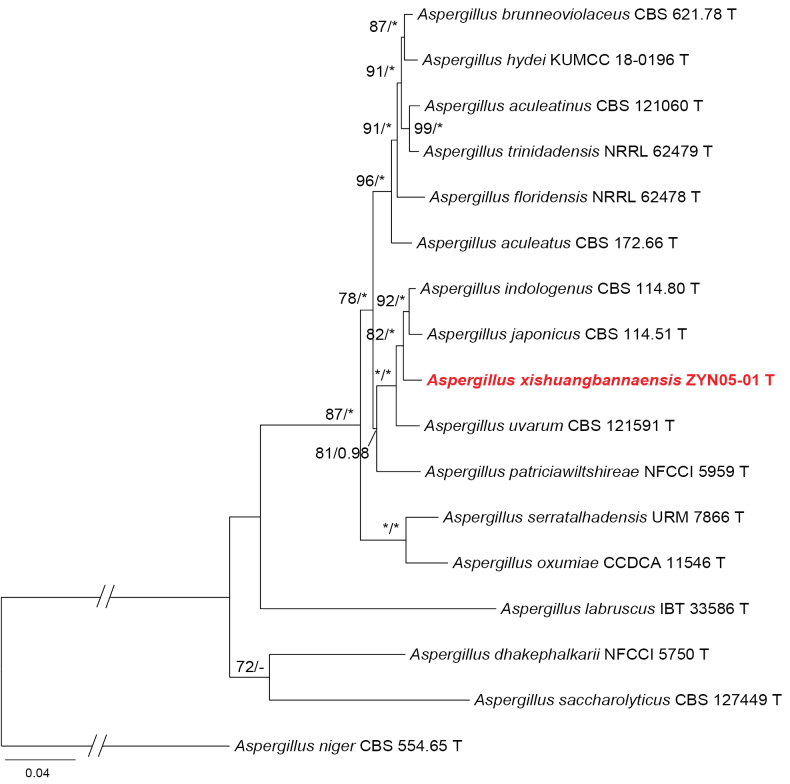 Figure 2
Figure 2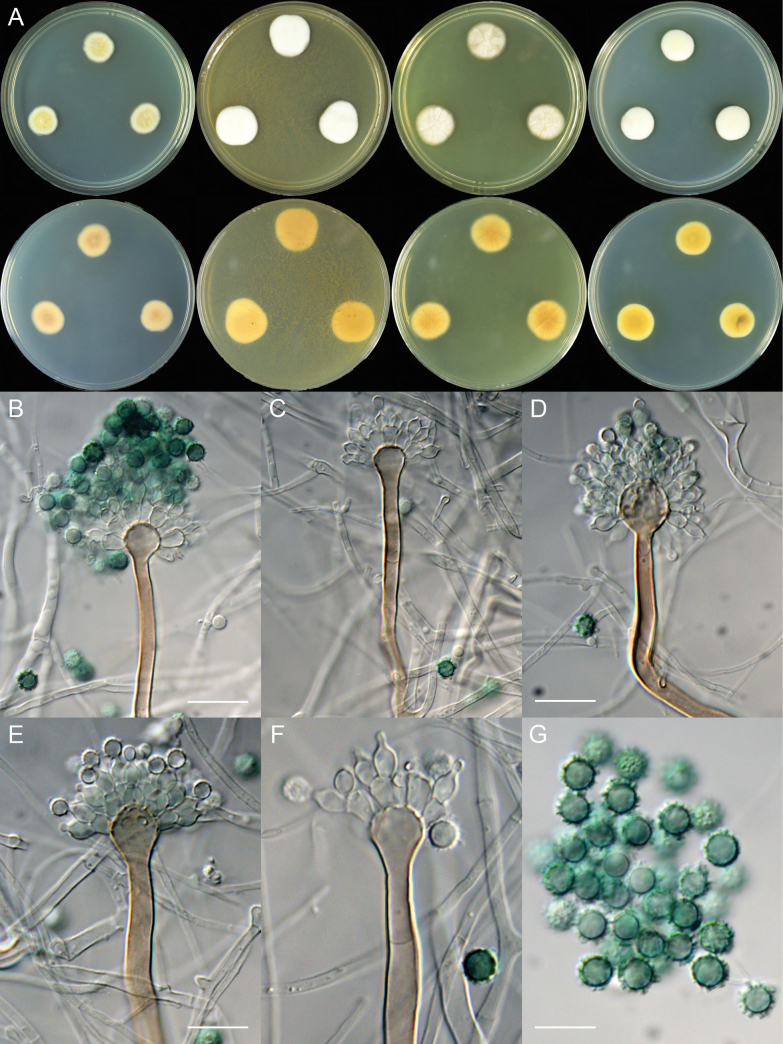 Figure 3
Figure 3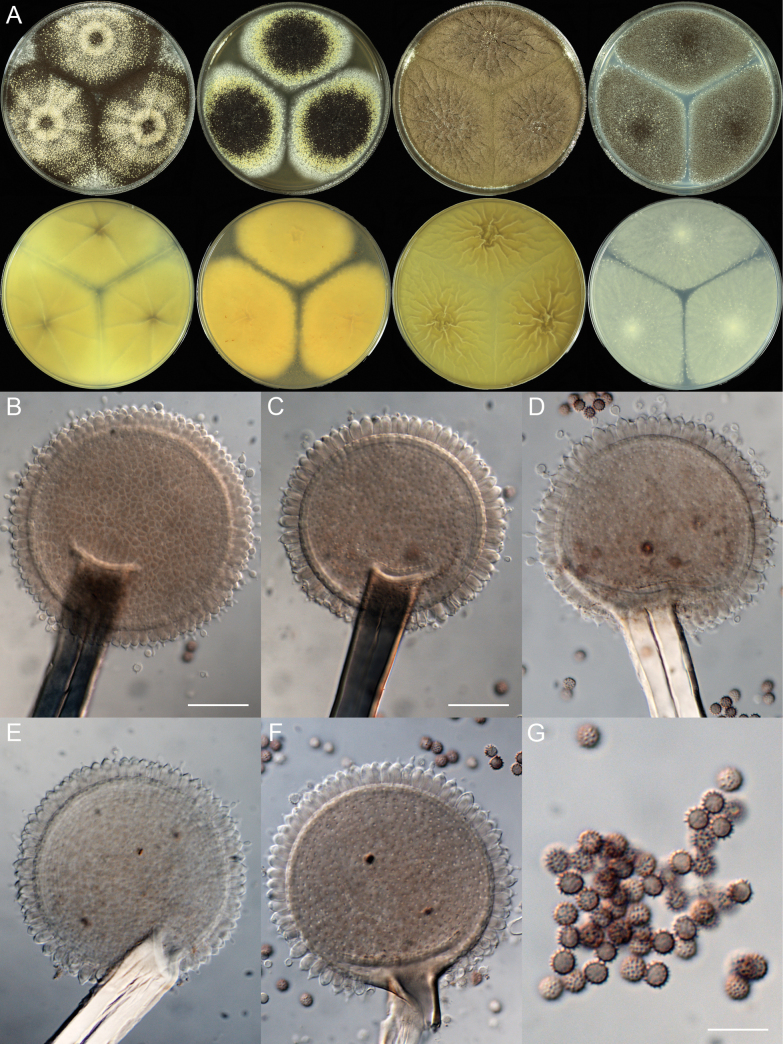 Figure 4
Figure 4| Series | Species | Strain | Locality | Substrate |
|
|
|
|
|---|---|---|---|---|---|---|---|---|
|
| CBS 123895 T | USA | chaparral of |
|
|
|
| |
| CBS 117.76 T | Romania | on walls of cave |
|
|
|
| ||
| CBS 419.69 T | Syria | soil |
|
|
|
| ||
| CBS 502.65 T | USA | desert soil |
|
|
|
| ||
|
| CBS 656.73 T | Egypt | sandy soil |
|
|
|
| |
|
| China: Hainan | sandy soil |
|
|
|
| ||
|
| JJJ40-31 T | China: Hebei | soil under |
|
|
|
| |
| JJJ40-12 | China: Hebei | soil under |
| n.a. |
|
|
| Series | Species | Strain | Locality | Substrate |
|
|
|
|
|---|---|---|---|---|---|---|---|---|
|
| CBS 121060 T | Thailand | dried parchment and green beans of |
|
|
|
| |
| CBS 172.66 T | Japan | unknown |
|
|
|
| ||
| CBS 621.78 T | Brazil | gills of |
|
|
|
| ||
| NFCCI 5750 T | India | rhizosphere soil associated with |
|
|
|
| ||
| NRRL 62478 T | USA | air |
|
|
|
| ||
| KUMCC 18-0196 T | China: Yunnan | air under |
|
|
|
| ||
| CBS 114.80 T | India | soil |
|
|
|
| ||
| CBS 114.51 T | unknown | unknown |
|
|
|
| ||
| IBT 33586 T | Brazil | fruit of |
|
|
|
| ||
| CCDCA 11546 T | Brazil | soil cultivated with |
| n.a. |
|
| ||
| NFCCI 5959 T | India | soil |
|
|
|
| ||
| CBS 127449 T | Denmark | toilet seat of treated |
|
|
|
| ||
| URM 7866 T | Brazil | soil |
|
|
|
| ||
| NRRL 62479 T | Trinidad-Tobago | air |
|
|
|
| ||
| CBS 121591 T | Italy | grape berries |
|
|
|
| ||
|
| ZYN05-01 T | China: Yunnan | soil in limestone seasonal rainforest |
|
|
|
| |
|
| CBS 554.65 T | France | unknown |
|
|
|
|
| Dataset | Gene fragment | No. of seq. | Length of alignment (bp) | No. of variable sites | No. of parsimony-informative sites | Model for |
|---|---|---|---|---|---|---|
|
|
| 8 | 548 | 64 | 44 | |
|
| 7 | 471 | 150 | 83 | ||
|
| 8 | 553 | 204 | 129 | ||
|
| 8 | 1071 | 241 | 151 | ||
| 8 | 2095 | 595 | 363 | TIMef+I | ||
|
|
| 16 | 530 | 235 | 108 | |
|
| 17 | 580 | 245 | 128 | ||
|
| 17 | 1052 | 273 | 155 | ||
| 17 | 2162 | 753 | 391 | TrN+I+G |
| Series | Species | Vesicles (µm) | Conidia shape | Conidia color | Conidia wall | Conidia size (µm) | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
| 15–17 | 12–15 | 19–20 | 18–19 | 8.5–13 × 8.5–13 | subglobose | vivid green | strongly echinulate | 6.0–7.5 | This study |
|
| 18–20 | no growth | 16–17 | 21–22 | 7.5–13 × 9.0–13 | subglobose | hyaline | strongly echinulate | 6.0–9.5 |
| |
|
|
| 64–70 | 35–57 | 55–60 | 68–70 | 62–80 × 60.5–77.5 | subglobose | brown | echinulate | 4.5–5.0 × 4.0–5.0 | This study |
|
| 60–70 | 20–50 | 60–70 | n.a. | 20–35 | globose to subglobose | brown | echinulate | 3.5–5.0 | ||
|
| 90 | 16–22 | 90 | n.a. | 20–30 | globose to subglobose | brown to black | conspicuously spinose | 3.0–4.0 |
| |
|
| 63–70 | n.a. | 57–70 | 76–80 | 20–45 | globose | brown | coarsely roughened to echinulate | 3.0–4.0 |
|
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal Biology and Applications · Mycorrhizal Fungi and Plant Interactions · Mycotoxins in Agriculture and Food
Introduction
Species of Aspergillus P. Micheli ex Haller are ubiquitous in various environments and have a long history of exploitation by humans. Some species have been used for food fermentations for centuries, especially in East Asia. Aspergillus oryzae (Ahlb.) Cohn and A. sojae Sakag. & K. Yamada ex Murak. play crucial roles in the production of rice wine, soybean pastes, and soy sauce (Bennett 2009), and A. niger Tiegh. is used for fermentations of baijiu (Xu et al. 2022) as well as pu-erh tea (Frisvad et al. 2007). More than 100 years ago, the ability of A. niger to produce citric acid was industrially exploited in 1919 (Schuster et al. 2002). Aspergillus terreus Thom was the first of the major statins to produce lovastatin, a medicine used to lower cholesterol (Bennett 2009). Aspergillus niger and A. terreus are also efficient agents for bioleaching of rare earth elements with low environmental impact (Mowafy 2020). Aspergillus cvjetkovicii Jurjević et al. can protect against phytopathogens through interspecies chemical signaling in the phyllosphere of rice as a biocontrol agent (Fan et al. 2024). On the other hand, some species pose severe threats to human health. Mycotoxins are produced by certain Aspergillus species, causing food contamination, e.g., aflatoxins by A. flavus Link and A. parasiticus Speare, and ochratoxin A by A. ochraceus G. Wilh. and A. niger (Xue et al. 2025). Aspergillus fumigatus Fresen. is known as the primary causative agent of aspergillosis, followed by A. flavus, A. niger, and A. terreus (Khan et al. 2024).
The genus Aspergillus was originally introduced in 1729 and validated in 1768. It was divided into six subgenera, 27 sections, and 75 series, with 446 species recognized (Houbraken et al. 2020). A new series, Aspergillus ser. Hainanici, was recently proposed (Wang and Zhuang 2022), and the accepted species of the genus increased to 453 (Visagie et al. 2024). Subsequently, 16 newly described species were added: one each from Europe and North America, two from South America, and the remaining 12 from Asia. Aspergillus albicolor D.S. Paiva was reported from Portugal, A. pseudoalabamensis Cañete-Gibas et al. from the USA, and A. alvaroi J.M.S. Lima et al. and A. guanovespertilionum J.M.S. Lima et al. from Brazil. Among the 12 Asian taxa, five were from China (A. cylindricus Zhi Y. Zhang et al., A. doliiformis Zhi Y. Zhang et al., A. liaoningensis C. Liu et al., A. plumeriae C. Liu et al., A. subinflatus C. Liu et al.), A. dhakephalkarii Rajeshk. et al. and A. patriciawiltshireae Rajeshk. et al. from India, A. hubkae Y.B. Zhou et al. and A. mahabadiensis Abdollahz. & O. Ghaderi from Iran, A. verrucosus R. Hagiuda & D. Hirose from Japan, A. ullungdoensis Hyang B. Lee from South Korea, and A. halopiscium V.N. Thanh et al. from Vietnam.
During investigations on the biodiversity of Aspergillaceae in China, two species of Aspergillus isolated from soil were identified as new to science based on sequence analyses and morphological comparisons. Detailed descriptions and illustrations are provided.
Materials and methods
Fungal materials
Cultures were isolated from soil samples collected from Hebei and Yunnan provinces, China. Dried cultures were preserved in the Herbarium Mycologicum Academiae Sinicae (HMAS, Beijing, China), and the living ex-type strains were deposited in the China General Microbiological Culture Collection Center (CGMCC, Beijing, China).
Morphological observations
Morphological characteristics were observed and recorded according to standardized methods (Samson et al. 2014). Four standard growth media were adopted: Czapek yeast autolysate agar (CYA; yeast extract, Oxoid, Hampshire, UK), malt extract agar (MEA; Amresco, Solon, OH, USA), yeast extract agar (YES), and potato dextrose agar (PDA). The methods for colonial inoculation, incubation, macroscopic and microscopic examinations, and digital capture followed our previous studies (Wang and Zhuang 2022; Wang et al. 2023; Peng et al. 2025).
DNA extraction, PCR amplification, and sequencing
DNA was extracted from living cultures grown on PDA for 7 days using the Plant Genomic DNA Kit (DP305; TIANGEN Biotech, Beijing, China). Polymerase chain reaction (PCR) amplifications of internal transcribed spacer (ITS), beta-tubulin (BenA), calmodulin (CaM), and RNA polymerase II second-largest subunit (RPB2) were conducted using routine methods (Samson et al. 2014). The forward and reverse primers used for each locus were as follows: ITS5 (5’-GGA AGT AAA AGT CGT AAC AAG G-3’) and ITS4 (5’-TCC TCC GCT TAT TGA TAT GC-3’) for ITS (White et al. 1990); Bt2a (5’-GGT AAC CAA ATC GGT GCT GCT TTC-3’) and Bt2b (5’-ACC CTC AGT GTA GTG ACC CTT GGC-3’) for BenA (Glass and Donaldson 1995); CMD5 (5’-CCG AGT ACA AGG ARG CCT TC-3’) and CMD6 (5’-CCG ATR GAG GTC ATR ACG TGG-3’) for CaM (Hong et al. 2005); and 5F (5’-GAY GAY MGW GAT CAY TTY GG-3’) and 7CR (5’-CCC ATR GCT TGY TTR CCC AT-3’) for RPB2 (Liu et al. 1999). The products were sequenced on an ABI 3730 DNA Sequencer (Applied Biosystems, Foster City, CA, USA).
Phylogenetic analyses
The newly generated forward and reverse sequences in this study were assembled using SeqMan v. 7.1.0 (DNASTAR Inc., Madison, WI, USA). The assembled sequences were deposited in GenBank, with accession numbers shown in bold (Tables 1, 2). The additional sequences used for phylogenetic analyses are also listed. Sequences were aligned using MAFFT v. 7.221 (Katoh and Standley 2013), either as individual single-gene datasets (ITS, BenA, CaM, and RPB2) or concatenated datasets. They were then manually edited and concatenated in BioEdit v. 7.1.10 (Hall 1999) and MEGA v. 11.0.13 (Tamura et al. 2021). Maximum likelihood (ML) analyses were performed using the IQ-TREE web server (Trifinopoulos et al. 2016) with the default automatic substitution model and bootstrap (BP) iteration (1,000 replicates) settings. Bayesian inference (BI) analyses were conducted with MrBayes v. 3.2.7 (Ronquist et al. 2012). Modeltest v. 3.7 (Posada and Crandall 1998) was adopted to determine appropriate nucleotide substitution models and parameters. Four MCMC chains (three heated and one cold) were run for at least 1 million generations, and posterior probability (PP) values were calculated based on the remaining 75% of trees after the burn-in phase. The consensus trees were viewed using FigTree v. 1.4.4 (http://tree.bio.ed.ac.uk/software/figtree, accessed on 28 December 2023).
Table 1.: Species and sequences used in the phylogenetic analyses for Aspergillus subgenus Nidulantes section Cavernicolarum.
Table 2.: Species and sequences used in the phylogenetic analyses for Aspergillus subgenus Circumdati section Nigri.
Results
To determine the species identities of the investigated strains, the single-gene datasets (ITS, BenA, CaM, and RPB2) and the concatenated three-locus (BenA + CaM + RPB2) dataset were compiled and analyzed. The detailed characteristics of the datasets are listed in Table 3.
In the phylogeny of section Cavernicolarum (Fig. 1), the strains JJJ40-12 and JJJ40-31, representing the same species, were located in series Hainanici as sister to Aspergillus hainanicus. The close relationship between these two species was strongly supported by statistical values, inferred either from the concatenated dataset (MLBP = 100, BIPP = 1.00; Fig. 1) or from the single-gene datasets (MLBP = 100; Suppl. material 1: figs S1–S4).
Maximum likelihood phylogeny of Aspergillus subgenus Nidulantes section Cavernicolarum inferred from the combined BenA, CaM, and RPB2 dataset. Bootstrap values ≥ 70% (left) or posterior probability values ≥ 0.95 (right) are indicated at nodes. Asterisks denote 100% bootstrap or 1.00 posterior probability.
In the phylogenetic tree of section Nigri series Japonici (Fig. 2), strain ZYN05-01 was grouped into a small clade associated with the following species: A. japonicus, A. indologenus, and A. uvarum. A similar tree topology was shown in the BenA analysis (Suppl. material 1: fig. S5), but it differed somewhat from the other two single-gene inferences. Strain ZYN05-01 clustered with A. uvarum in the CaM tree (MLBP = 82; Suppl. material 1: fig. S6), but it was closely related to A. japonicus in the RPB2 phylogeny (MLBP = 94; Suppl. material 1: fig. S7).
Maximum likelihood phylogeny of Aspergillus subgenus Circumdati section Nigri series Japonici inferred from the combined BenA, CaM, and RPB2 dataset. Bootstrap values ≥ 70% (left) or posterior probability values ≥ 0.95 (right) are indicated at nodes. Asterisks denote 100% bootstrap or 1.00 posterior probability.
Taxonomy
Aspergillus
hebeiensis
Taxon classificationFungiEurotialesAspergillaceae
X.C. Wang, L.Y. Peng & W.Y. Zhuang sp. nov.
55DC9DF9-D078-54D2-9A92-AAFA25F05FD1
Fungal Names: FN 572829
Etymology.
The specific epithet refers to the type locality.
In Aspergillus subgenus Nidulantes section Cavernicolarum series Hainanici.
Typification.
China • Hebei Province, Handan City, Fengfeng Mining District, Xiangtangshan Caves, one of the First Batch of Key Cultural Relics Units under National Protection of China, Northern Xiangtangshan, 36°32'2"N, 114°9'40"E, in soil under Platycladus orientalis (L.) Franco, 17 July 2023, Xin-Cun Wang, culture, Lu-Yao Peng, JJJ40-31 (holotype HMAS 354080, ex-type strain CGMCC 3.29151).
Aspergillus hebeiensis (JJJ40-31). A. Colonies: top row left to right, obverse CYA, MEA, YES, and PDA; bottom row left to right, reverse CYA, MEA, YES, and PDA; B–F. Conidiophores; G. Conidia. Scale bars: 17.5 µm (B, C); 15 µm (D); 12.5 µm (E); 10 µm (G, F).
DNA barcodes.
ITSPV883250, BenAPV877068, CaMPV877070, RPB2PV877073.
Colony diam.
7 days, 25 °C (unless stated otherwise): CYA 15–17 mm; CYA 37 °C 12–15 mm; MEA 19–20 mm; YES 18–19 mm; PDA 16–17 mm.
Colony characteristics.
On CYA 25 °C, 7 days: Colonies nearly circular, slightly concave at centers; margins moderately wide, entire; mycelia white and then yellow; texture velutinous; sporulation sparse; conidia en masse greyish; soluble pigments absent; exudates absent; reverse buff to yellow brown.
On CYA 37 °C, 7 days: Colonies nearly circular or irregular, slightly protuberant at centers; margins narrow to moderately wide, fimbriate; mycelia white and then buff; texture velutinous; sporulation sparse; conidia en masse brownish; soluble pigments absent; exudates absent; reverse buff to yellow brown.
On MEA 25 °C, 7 days: Colonies irregular, protuberant; margins narrow, entire; mycelia white and then cream; texture velutinous; sporulation sparse; conidia en masse creamish; soluble pigments absent; exudates absent; reverse buff to yellow brown.
On YES 25 °C, 7 days: Colonies nearly circular, concave at centers, radially sulcate; margins narrow, entire; mycelia pale; texture velutinous; sporulation sparse; conidia en masse greyish; soluble pigments absent; exudates absent; reverse yellow brown to orange brown.
On PDA 25 °C, 7 days: Colonies nearly circular, protuberant; margins narrow, entire; mycelia white and then cream; texture velutinous; sporulation sparse; conidia en masse creamish to brownish; soluble pigments yellow; exudates absent; reverse buff to yellow brown, occasionally with dark brown sectors.
Micromorphology.
Conidial heads radiate; stipes short, 65–110 (–140) × 4.0–7.0 µm, not septate, walls thick, smooth, brown; vesicles 8.5–13 × 8.5–13 µm, subglobose to globose; biseriate; metulae 5.0–8.5 × 3.5–6.5 µm, cylindrical to obovate, covering almost a half to two-thirds surface of the vesicle; phialides 5.5–8.0 × 3.5–4.0 µm, flask-shaped; conidia 6.0–7.5 µm, subglobose, vivid green, strongly echinulate.
Additional strain examined.
China • Hebei Province, Handan City, Fengfeng Mining District, Xiangtangshan Caves, one of the First Batch of Key Cultural Relics Units under National Protection of China, Northern Xiangtangshan, 36°32'2"N, 114°9'40"E, in soil under Platycladus orientalis (L.) Franco, 17 July 2023, Xin-Cun Wang, culture, Yi-Fan Wang, JJJ40-12.
Notes.
This species is the second member of series Hainanici and sister to A. hainanicus (Fig. 1; Suppl. material 1: figs S1–S4). It differs from the latter by 29 bp for BenA (93.76% sequence identity), 16 bp for CaM (97.03%), and 21 bp for RPB2 (98.04%). Morphologically, although both species have short stipes, biseriate conidiophores, and strongly echinulate conidia, the new species is easily distinguished from A. hainanicus by growth on CYA at 37 °C, vivid green, and smaller conidia (6.0–7.5 vs. 6.0–9.5 µm, Table 4).
Aspergillus
xishuangbannaensis
Taxon classificationFungiEurotialesAspergillaceae
X.C. Wang, L.Y. Peng & W.Y. Zhuang sp. nov.
7BF0D4D4-54A3-526F-A181-CEFF4E455AB4
Fungal Names: FN 572923
Etymology.
The specific epithet refers to the type locality.
In Aspergillus subgenus Circumdati section Nigri series Japonici.
Typification.
China • Yunnan Province, Xishuangbanna Dai Autonomous Prefecture, Mengla County, Menglun Town, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Green Stone Forest, 21°54'39"N, 101°17'00"E, in soil of limestone seasonal rainforest, 28 May 2024, Zhao-Qing Zeng, culture, Xiao Mou, ZYN05-01 (holotype HMAS 354081, ex-type strain CGMCC 3.29152).
Aspergillus xishuangbannaensis (ZYN05-01). A. Colonies: top row left to right, obverse CYA, MEA, YES, and PDA; bottom row left to right, reverse CYA, MEA, YES, and PDA; B–F. Conidiophores; G. Conidia. Scale bars: 22.5 µm (B); 20 µm (C–F); 10 µm (G).
DNA barcodes.
ITSPV883252, BenAPV877069, CaMPV877072, RPB2PV877075.
Colony diam.
7 days, 25 °C (unless stated otherwise): CYA 64–70 mm; CYA 37 °C 35–57 mm; MEA 55–60 mm; YES 68–70 mm; PDA 66–70 mm.
Colony characters.
CYA 25 °C, 7 d: Colonies deep, plane, radially sulcate; margins entire; mycelium white; texture velutinous; sporulation moderately dense; conidia en masse dark brown; soluble pigments absent; exudates absent; sclerotia yellow, abundant; reverse yellow brown.
CYA 37 °C, 7 d: Colonies deep, protuberant, radially sulcate; margins narrow, entire; mycelium white; texture velutinous; sporulation dense; conidia en masse dark brown; soluble pigments absent; exudates tiny; reverse yellow brown to greyish black.
MEA 25 °C, 7 d: Colonies plane; margins moderately wide, entire; mycelium white; texture velutinous; sporulation dense; conidia en masse blackish brown; soluble pigments absent; exudates absent; sclerotia yellow, abundant; reverse yellow brown.
YES 25 °C, 7 d: Colonies deep, radially sulcate; margins entire; mycelium white; texture velutinous; sporulation very dense; conidia en masse purplish brown; soluble pigments absent; exudates absent; sclerotia yellow or white; reverse yellow brown.
PDA 25 °C, 7 d: Colonies plain; margins narrow, entire; mycelium white; texture velutinous; sporulation dense; conidia en masse dark brown; soluble pigments absent; exudates absent; sclerotia yellow; reverse white.
Micromorphology.
Conidial heads radiate; stipes 400–975 × 12–21.5 µm, not septate, walls thick, smooth, hyaline, brownish or black; vesicles 62–80 × 60.5–77.5 µm, globose or subglobose; uniseriate; phialides 6.5–8.5 × 3.5–5.5 µm, flask-shaped and cover the entire surface of the vesicle; conidia 4.5–5.0 × 4.0–5.0 µm, subglobose, echinulate, light brown to dark brown when mature.
Notes.
Aspergillus xishuangbannaensis is molecularly and morphologically differentiated from its closely related sisters: A. japonicus, A. uvarum, and A. indologenus. For the BenA gene, it differs from A. japonicus by 13 bp (97.27% sequence identity), from A. uvarum by 16 bp (96.26%), and from A. indologenus by 14 bp (97.06%); for the CaM region, it differs from A. japonicus by 20 bp (95.97%), from A. uvarum by 15 bp (96.98%), and from A. indologenus by 17 bp (96.72%); and for the RPB2 fragment, it differs from A. japonicus by five bp (99.50%), from A. uvarum by 19 bp (98.19%), and from A. indologenus by seven bp (99.33%). Although species of this series are similar in gross morphology, the new species can be easily separated from its sisters by much larger vesicles (Table 4). Additionally, sclerotia were not observed in A. indologenus, while white to cream sclerotia were often produced by A. japonicus, yellow ones in the new species, and dark brown to black ones in A. uvarum (Klich 2002; Samson et al. 2007; Perrone et al. 2008; Varga et al. 2011).
Discussion
In our previous study, four new species of Aspergillus were described from the Xisha Islands in the South China Sea (Wang and Zhuang 2022). That was the first report of a new species of the genus from Chinese tropical islands, suggesting that the biodiversity level of Aspergillus might be underestimated in marine environments. Aspergillus liaoningensis, A. plumeriae, and A. subinflatus were later found in tidal flat sediments (Liu et al. 2023), and A. halopiscium occurred in the dried marine anchovy Stolephorus commersonnii Lacepède (Crous et al. 2025). These accumulated findings reveal that marine environments are biodiversity hotspots for the genus.
In contrast, the two new species discovered in this study are both from terrestrial ecosystems, including a cultural heritage site and a nature reserve. This suggests that terrestrial ecosystems also should not be overlooked in biodiversity investigations.
The Xiangtangshan Caves, located in the southern part of Hebei Province in North China, are a famous cultural treasure containing 16 caves adorned with over 4,000 Buddhist sculptures dating back more than 1,400 years. The new species A. hebeiensis was isolated from a soil sample collected in this area. Chinese Buddhist temples have been shown to play an important role in preserving regional biodiversity (Wang et al. 2020; Huang et al. 2025). Further explorations in these traditional cultural and religious areas should be emphasized.
Caves are another biodiversity hotspot, and several Aspergillus species were originally reported from them. Aspergillus alvaroi was isolated from sediment in a Brazilian cave, A. guanovespertilionum from hematophagous bat guano (Lima et al. 2024), A. lebretii V.C.S. Alves et al. from cave air (Alves et al. 2022), A. okavangoensis Visagie & Nkwe from bat guano-contaminated soil in a Botswana cave (Visagie et al. 2021), and A. limoniformis Z.F. Zhang & L. Cai, A. phialiformis Z.F. Zhang & L. Cai, and A. phialosimplex Z.F. Zhang & L. Cai from dung of Chiroptera, rock, and plant debris in Chinese caves, respectively (Zhang et al. 2021).
The tropical seasonal rain forest in Xishuangbanna is one of the most species-rich forest ecosystems in China. This area is also one of the biodiversity hotspots in the world. Xishuangbanna National Nature Reserve is one of China’s first nature reserves, established in 1958. It consists of five geographically disconnected sub-reserves: Mangao Reserve, Mengla Reserve, Menglun Reserve, Mengyang Reserve, and Shangyong Reserve. Xishuangbanna Tropical Botanical Garden, located in Menglun Town and covering 1,125 hectares, is home to more than 14,000 plant species, including rare orchids and ancient cycads. Many fungal species were first described from Xishuangbanna, e.g., Chloridium xishuangbannaense W.P. Wu & Y.Z. Diao (Wu and Diao 2022), Talaromyces bannicus L. Wang (Wei et al. 2021), and Trichoderma bannaense Kai Chen & W.Y. Zhuang (Chen and Zhuang 2017). However, no new species of Aspergillaceae had been recorded from this district. Aspergillus xishuangbannaensis, described here, was isolated from a soil sample collected in this area. More surveys are urgently needed to explore the species diversity of this group in diverse terrestrial ecosystems in China.
Supplementary Material
XML Treatment for Aspergillus hebeiensis
XML Treatment for Aspergillus xishuangbannaensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alves VCS Lira RA Lima JMS Barbosa RN Bento DM Barbier E Bernard E Souza-Motta CM Bezerra JDP (2022) Unravelling the fungal darkness in a tropical cave: Richness and the description of one new genus and six new species.Fungal Systematics and Evolution 10: 139–167. 10.3114/fuse.2022.10.0636741552 PMC 9875697 · doi ↗ · pubmed ↗
- 2Bennett JW (2009) Aspergillus: A primer for the novice. Medical Mycology 47(Suppl 1): S 5–S 12. 10.1080/1369378080271251519253144 · doi ↗ · pubmed ↗
- 3Chen K Zhuang WY (2017) Discovery from a large-scaled survey of Trichoderma in soil of China. Scientific Reports 7: 9090. 10.1038/s 41598-017-07807-3PMC 556733028831112 · doi ↗ · pubmed ↗
- 4Crous PW Catcheside DEA Catcheside PS Alfenas AC Alfenas RF Barreto RW Lebel T Balashov S Broadbridge J Jurjevic Z De la Pena-Lastra S Hoffmann R Mateos A Riebesehl J Shivas RG Soliz Santander FF Tan YP Altes A Bandini D Carriconde F Cazabonne J Czachura P Gryta H Eyssartier G Larsson E Pereira OL Rigueiro-Rodriguez A Wingfield MJ Ahmad W Bibi S Denman S Esteve-Raventos F Hussain S Illescas T Luangsa-Ard JJ Moller L Mombert A Noisripoom W Olariaga I Pancorbo F Paz A Piatek M Polman-Short C Suarez E Afshan NS Ali H Arzanlou M Ayer F Barratt J Bellanger JM Bidaud A Bishop-Hurley SL Bohm M Bose T Campo EC · doi ↗ · pubmed ↗
- 5Fan XY Matsumoto H Xu HR Fang HD Pan QQ Lv TX Zhan CF Feng XX Liu XY Su DR Fan MY Ma ZH Berg G Li SJ Cernava T Wang MC (2024) Aspergillus cvjetkovicii protects against phytopathogens through interspecies chemical signalling in the phyllosphere.Nature Microbiology 9: 2862–2876. 10.1038/s 41564-024-01781-z 39103572 · doi ↗ · pubmed ↗
- 6Frisvad JC Larsen T Ode Vries R Meijer M Houbraken J Cabanes FJ Ehrlich K Samson RA (2007) Secondary metabolite profiling, growth profiles and other tools for species recognition and important Aspergillus mycotoxins.Studies in Mycology 59: 31–37. 10.3114/sim.2007.59.0418490955 PMC 2275202 · doi ↗ · pubmed ↗
- 7Glass NL Donaldson GC (1995) Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes.Applied and Environmental Microbiology 61: 1323–1330. 10.1128/aem.61.4.1323-1330.19957747954 PMC 167388 · doi ↗ · pubmed ↗
- 8Hall TA (1999) Bio Edit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT.Nucleic Acids Symposium Series 41: 95–98.