The genus Batophila Foudras, 1860 (Coleoptera, Chrysomeliae, Galerucinae, Alticitae) in Taiwan, with descriptions of 11 new species

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2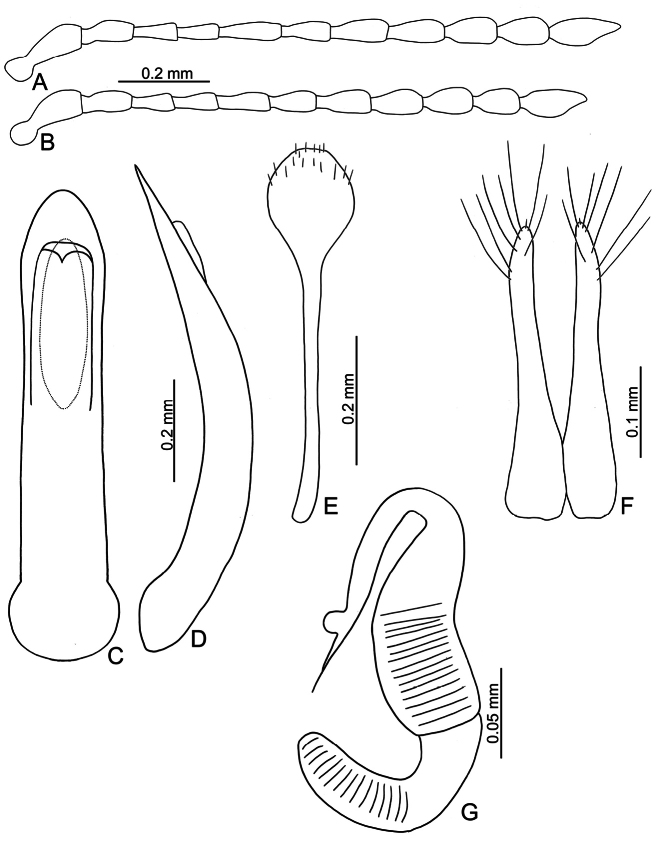 Figure 3
Figure 3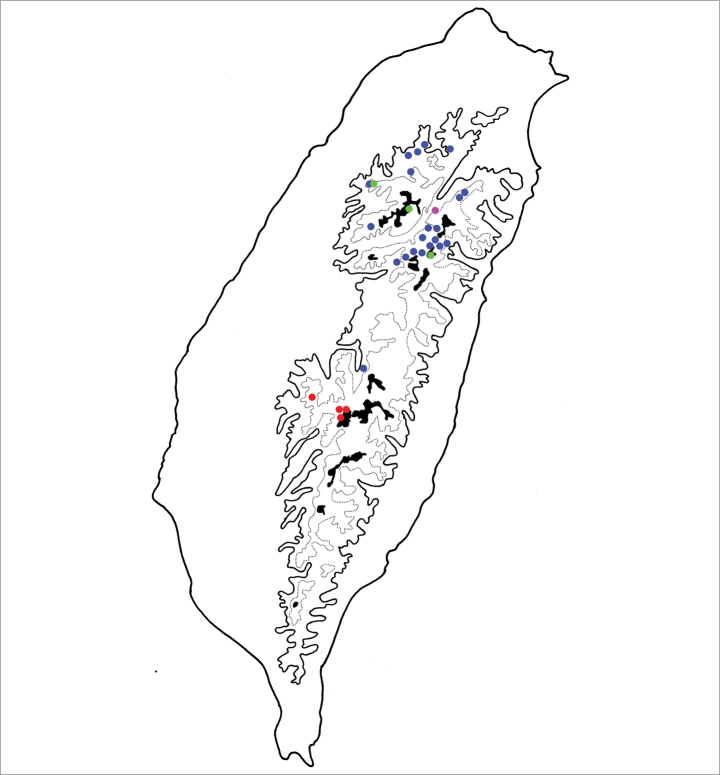 Figure 4
Figure 4 Figure 5
Figure 5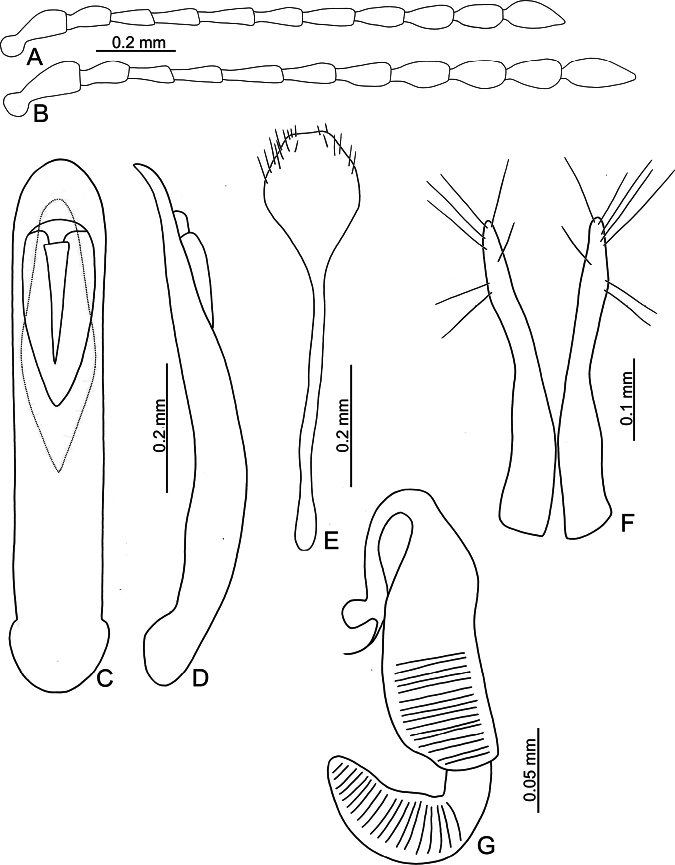 Figure 6
Figure 6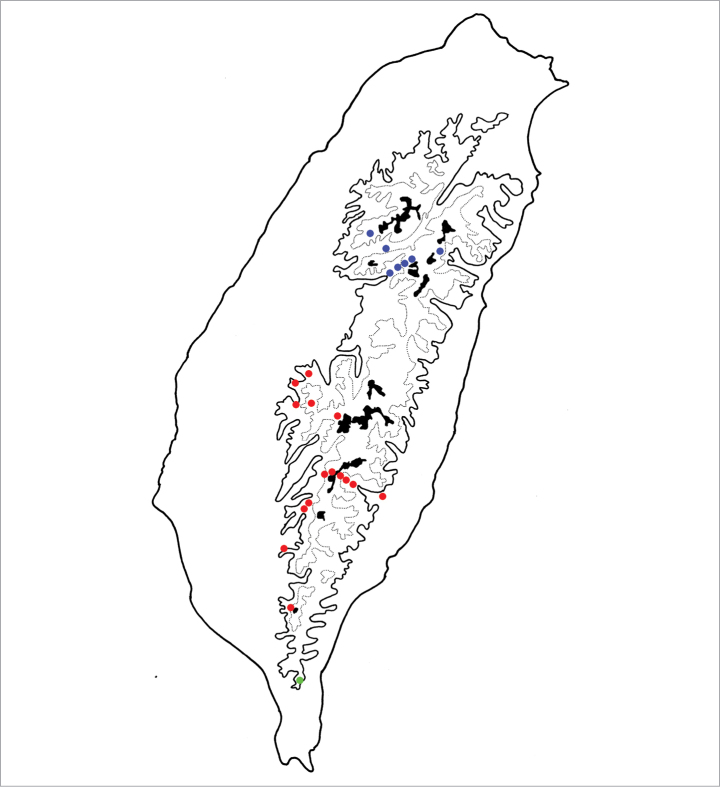 Figure 7
Figure 7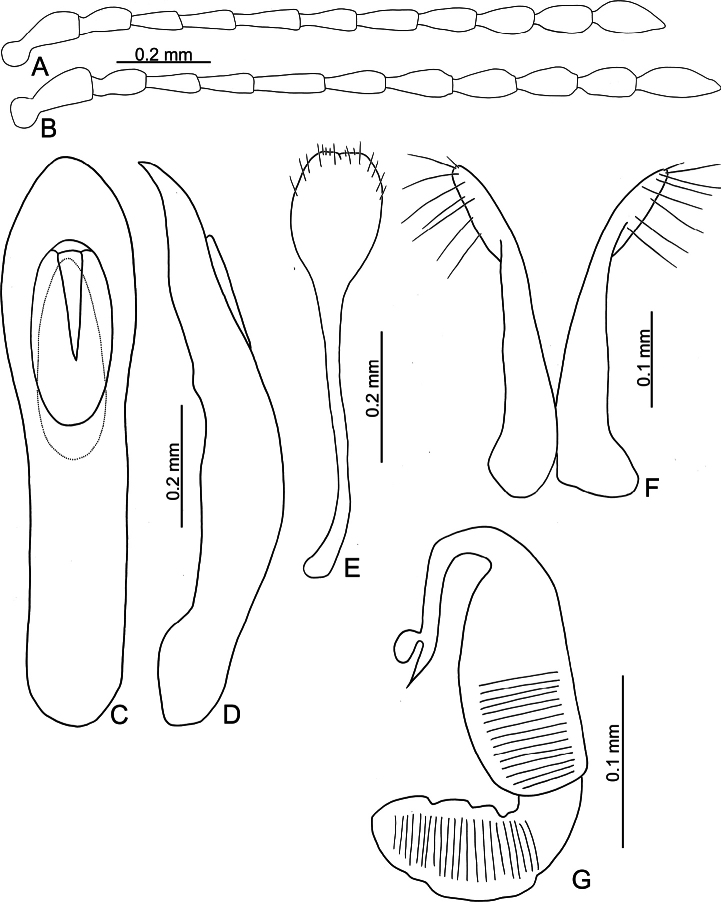 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10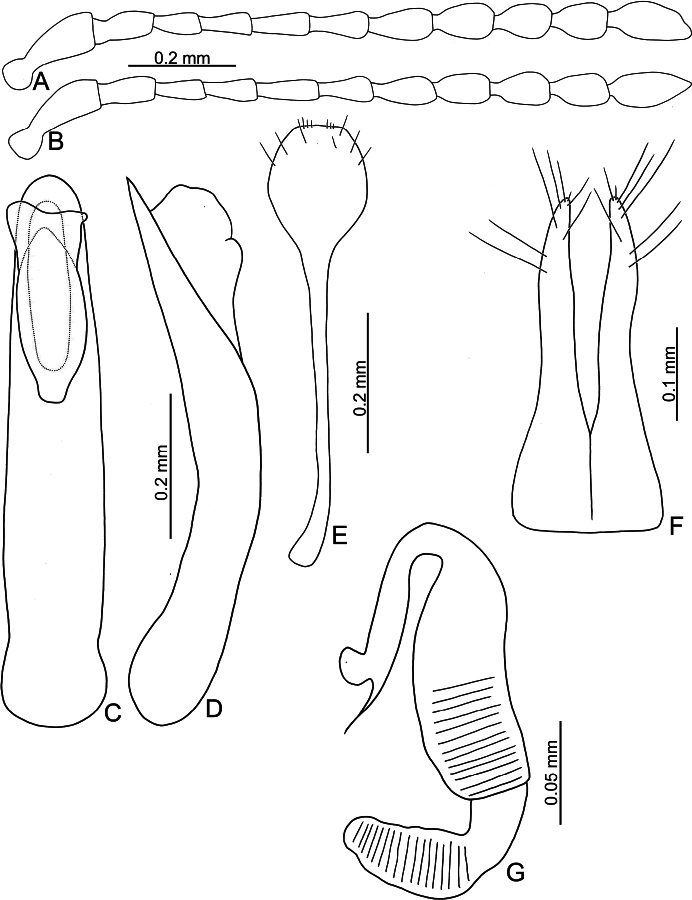 Figure 11
Figure 11 Figure 12
Figure 12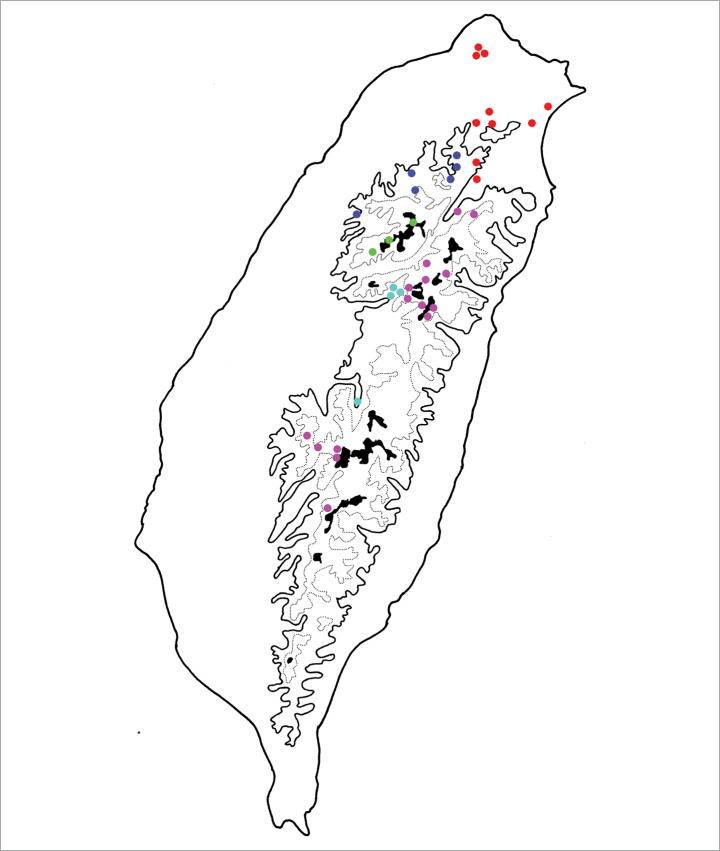 Figure 13
Figure 13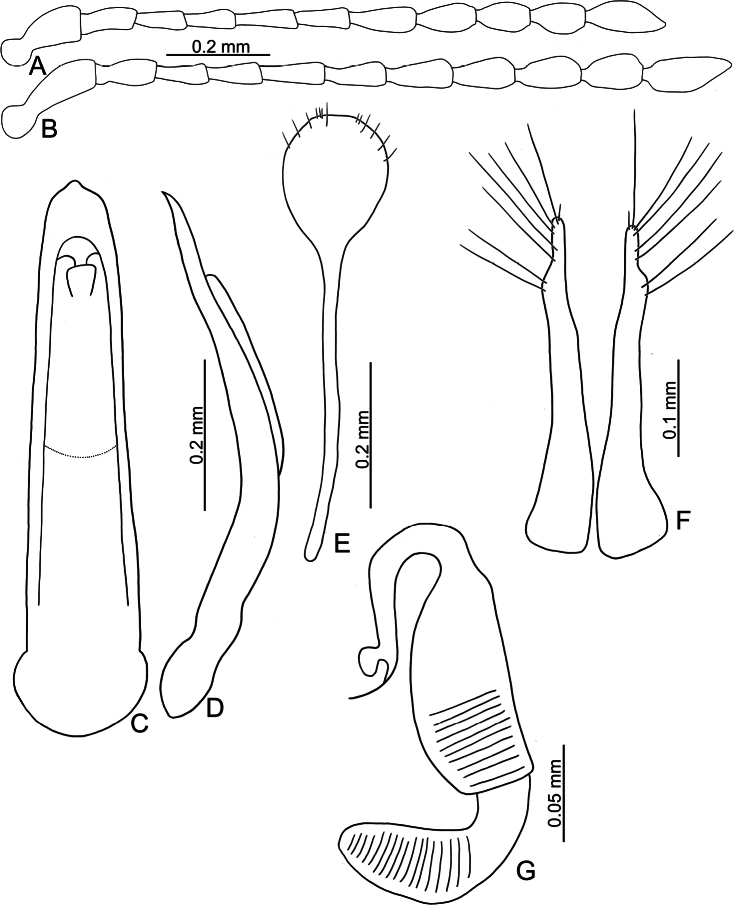 Figure 14
Figure 14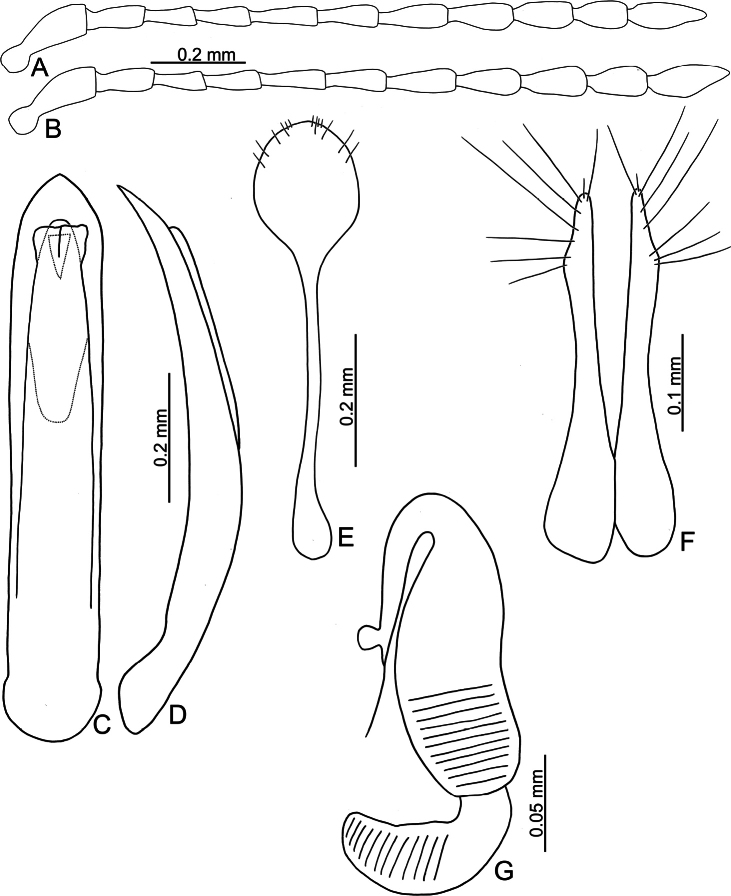 Figure 15
Figure 15 Figure 16
Figure 16 Figure 17
Figure 17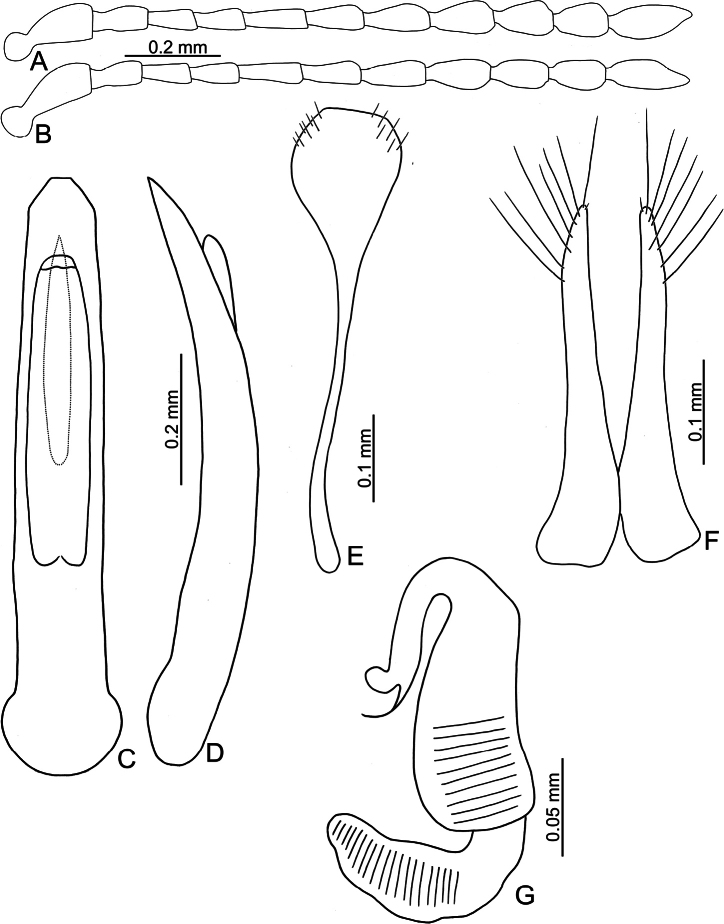 Figure 18
Figure 18 Figure 19
Figure 19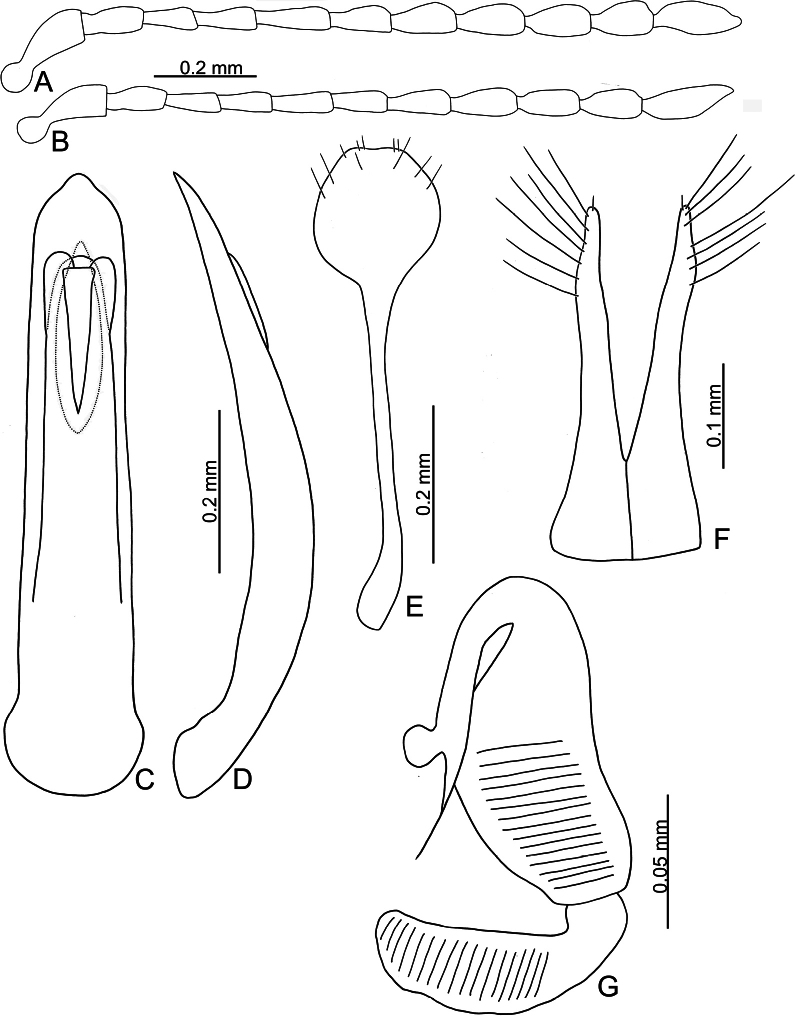 Figure 20
Figure 20 Figure 21
Figure 21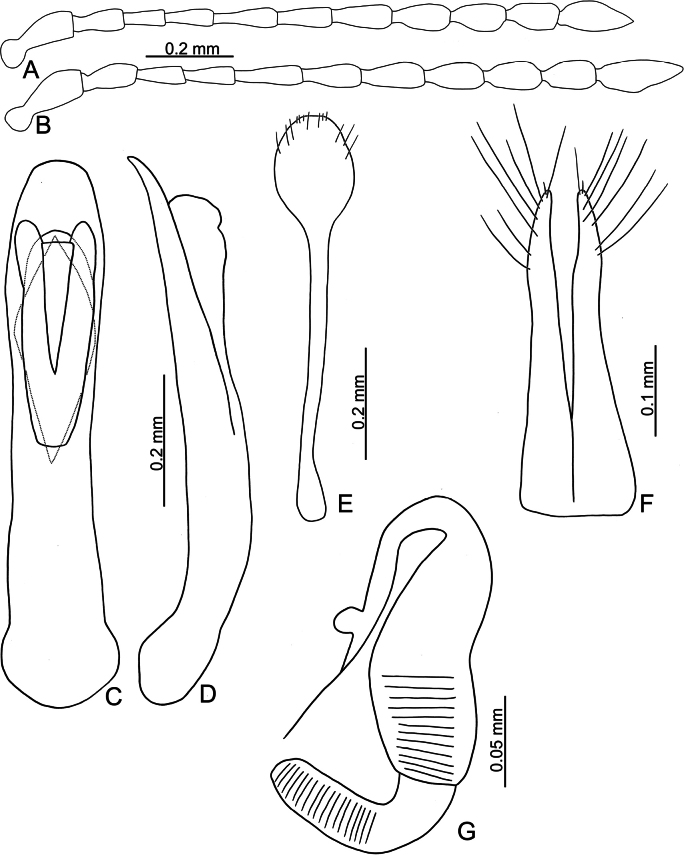 Figure 22
Figure 22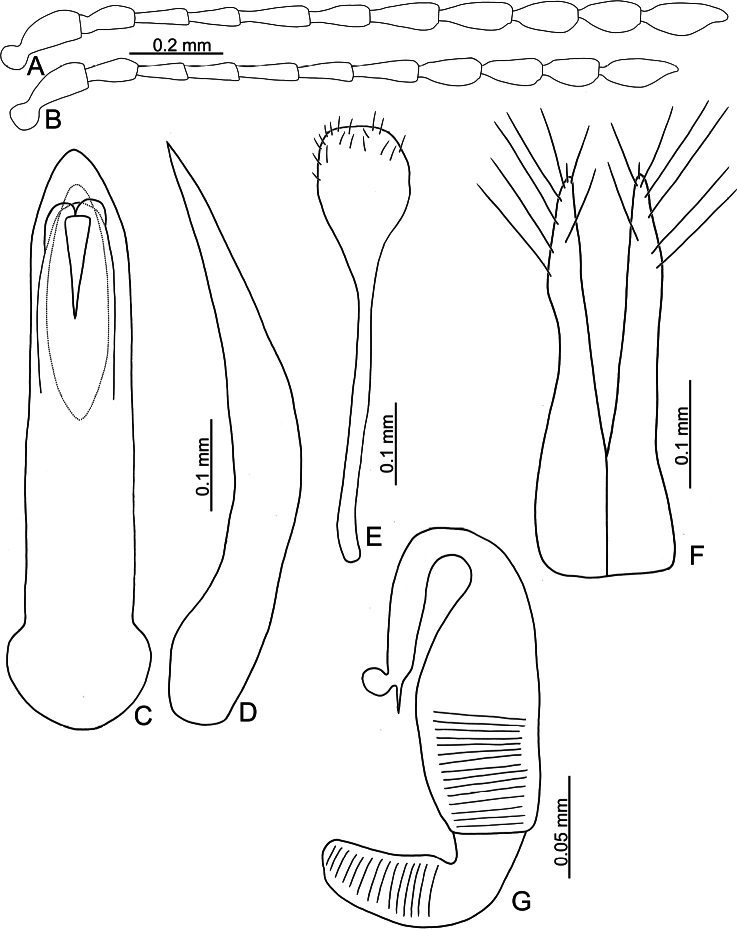 Figure 23
Figure 23 Figure 24
Figure 24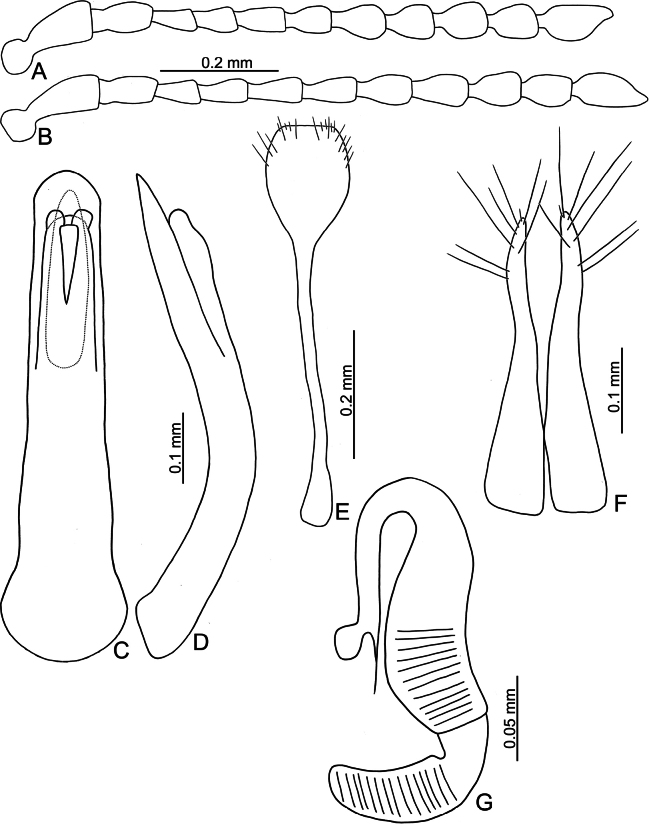 Figure 25
Figure 25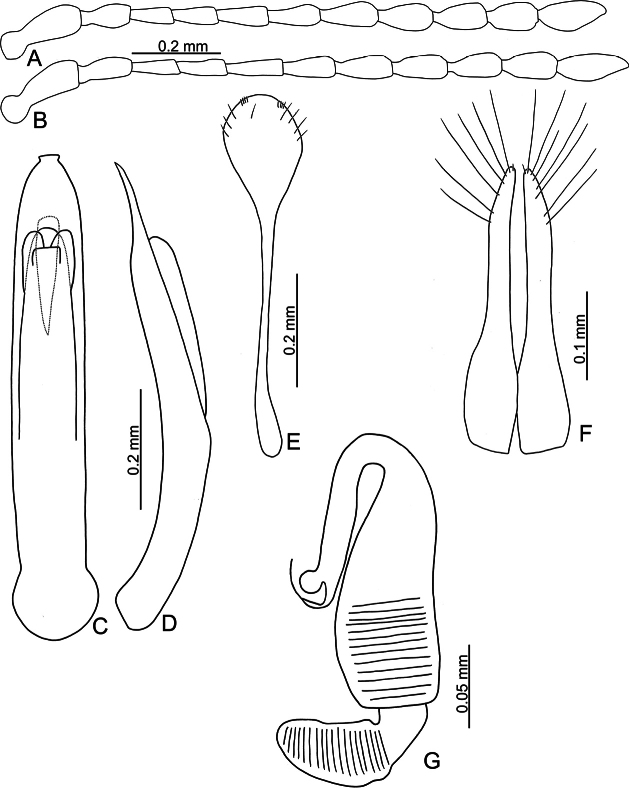 Figure 26
Figure 26| 1 | Elytra abruptly, apically narrowed (Figs |
|
| – | Elytra gradually, apically narrowed (Figs |
|
| 2 | Elytra dorsoventrally convex, apex not visible in dorsal view in both sexes; aedeagus parallel-sided (Fig. |
|
| – | Elytra dorsoventrally convex only in female but flattened in males; aedeagus subapically widened (Fig. |
|
| 3 | Aedeagus slightly wide in apical 1/5 (Fig. |
|
| – | Aedeagus extremely wide in apical 1/5 (Fig. |
|
| 4 | Elytral apices convergent, or divergent and rounded (Figs |
|
| – | Elytral apices truncate (Figs |
|
| 5 | Antennae stout, antennomeres VI–X 0.5 as long as antennomere I, apex of aedeagus widely rounded (Figs |
|
| – | Antennae slender, antennomeres VI–X > 0.5 as long as antennomere I (Figs |
|
| 6 | Longitudinal ridges on elytra distinct and sexually dimorphic; apex of aedeagus truncate (Fig. |
|
| – | Longitudinal ridges on elytra instinct or reduced; apex of aedeagus rounded but with one rounded process at middle (Fig. |
|
| 7 | Punctures on elytra tiny (Fig. |
|
| – | Punctures on elytra coarse (Figs |
|
| 8 | Apex of aedeagus tapering (Figs |
|
| – | Apex of aedeagus rounded, but with one process at middle of apical margin (Figs |
|
| 9 | Apex of aedeagus tapering from apical 1/10; in high mountains of Taichung and Miaoli counties (Fig. |
|
| – | Apex of aedeagus tapering from apical 1/5; in lowlands of Nantou County (Fig. |
|
| 10 | Apex of aedeagus widely rounded (Fig. |
|
| – | Apex of aedeagus rounded, with one process at middle of apical margin (Figs |
|
| 11 | Rounded apex of aedeagus with one rounded process at middle of apical margin (Fig. |
|
| – | Rounded apex of aedeagus with one truncate process at middle of apical margin (Fig. |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsColeoptera Taxonomy and Distribution · Coleoptera: Cerambycidae studies · Forest Ecology and Biodiversity Studies
Introduction
The flea beetle genus Batophila Foudras, 1860 can be recognized easily by the combination of the following characters: general color metallic; anterior coxal cavities open posteriorly; dorsum not pubescent; pronotum evenly convex, without a distinct antebasal, transverse impression; elytron with reduced humeral calli, punctures arranged into 10 rows; hind wings absent; prosternum exceeding posterior margin of coxae; hind tibia with apical, simple spine; tarsomere III bilobed (Yang et al. 2015).
This genus contains 30 species recorded from Oriental and Palearctic regions (Nadein 2013), 24 of them from the Palearctic (Bezděk and Konstantinov 2024). Chûjô (1937) initially recorded the genus from Taiwan based on B. yangweii Chen, 1933, which was synonymized with B. acutangula Heikertinger, 1921 by Heikertinger (1948). Döberl (2010) added the second species, B. taiwanica Döberl, 2010, to the Taiwanese fauna. Adults of the genus are common in various forest types and easily collected by sweeping. More than 3,400 specimens were available for study, including historical collections at several museums, and extensive collections made by the Taiwan Chrysomelid Research Team (TCRT) (Lee 2025). More than 3,000 specimens were collected and deposited at the Taiwan Agricultural Research Institute during 1979–1984. The collecting sites include a number of places along Provincial Highway 14A, including Wushe (霧社, 1250 m a.s.l.), Yu-shih (幼獅, 1750 m a.s.l.), Sungkang (松崗, 2100 m), Meifeng (梅峰, 2150 m a.s.l.), Tsuifeng (翠峰, 2300 m a.s.l.), and Tayuling (大禹嶺, 2560 m a.s.l.). Other sites include Alishan (阿里山, 2400 m a.s.l.), Tungpu (東埔, 1200 m a.s.l.), and Wufeng (五峰, 400 m a.s.l.). The true species diversity and distributions can now be presented based on sufficient material.
Materials and methods
For taxonomic study, abdomens of adults were separated from the forebodies and boiled in 10% KOH solution, followed by washing in distilled water to prepare genitalia for illustrations. The genitalia were then dissected from the abdomens, mounted on slides in glycerin, and studied and drawn using a Leica M165 stereomicroscope. For detailed examinations, a Nikon ECLIPSE 50i microscope was used.
At least three males and females from each species were examined to delimit variability of diagnostic characters. For species collected from more than one locality or with color variations, at least one pair of each sex from each locality and color morph was examined. Length was measured from the anterior margin of the eye to the elytral apex, and width at the greatest width of the elytra. Nomenclature for morphological structures of adults follows Duckett and Daza (2004). Names of plant species follow the Taiwan Encyclopedia of Life (2024).
Specimens studied herein are deposited at the following institutes and collections:
KMNHKitakyushu Museum of Natural History and Human History, Kitakyushu, Japan [Yûsuke Minoshima]
MHNGMuséum d’Histoire Naturelle, Genève, Switzerland [Giulio Cuccodoro]
NHMBNaturhistorisches Museum, Basel, Switzerland [Matthias Borer]
NHMUKThe Natural History Museum, London, UK [Michael F. Geiser]
NMNSNational Museum of Natural Science, Taichung, Taiwan [Bao-Cheng Lai]
TARIApplied Zoology Division, Taiwan Agricultural Research Insitute, Taichung, Taiwan [Chi-Feng Lee]
Exact label data are cited for all type specimens of described species; a double slash (//) divides the data on different labels and a single slash (/) divides the data in different rows. Other comments and remarks are in square brackets: [p] – preceding data are printed, [h] – preceding data are handwritten, [r] – red label, [w] – white label.
Taxonomic account
Batophila
acutangula
Taxon classificationAnimaliaColeopteraChrysomelidae
Heikertinger, 1921
FFEDA544-141B-576D-85AE-D2606D5EB2D6
Batophila acutangula Heikertinger, 1921: 7. Batophila yangweii Chen, 1933: 250; Heikertinger, 1948: 53 (as synonym for B. acutangula).
Type specimens examined.
Lectotype ♂ (NHMB): “H. Frieb / Perewaja-Rjetsch / Ka-Tal, 7–8 km / nördlich von / Wladiwostok [h, w] // Batophila / acutangula m. [h] / det. Heiktgr. [p, w] // Batoph. / acutang. / Type! [h, r] // 1953 Coll. / Heikertinger [p, w] // lectotype [h] / J. Bechyne det., 1956 [p, w]”
Notes.
Batophila acutangula Heikertinger, 1921 is not found in Taiwan although it was recorded from Taiwan previously (Chûjô 1937). Voucher specimens of B. yangweii (Chûjô 1937) are identified as B. houjayi sp. nov., B. huangi sp. nov., B. taiwanica Döberl, 2010, B. tsoui sp. nov., B. wusheensis sp. nov., and B. yuae sp. nov. Voucher specimens of B. acutangula (Kimoto 1971, 1989) are identified as B. alishanensis sp. nov., B. choui sp. nov., B. houjayi sp. nov., B. taiwanica Döberl, 2010. Several subspecies of B. acutangula in Japan were synonymized by Kimoto (1966); however, they may represent valid species since their aedeagi are different in shape, as illustrated by Kimoto (1966).
Type specimens and labels A. Batophila acutangula Heikertinger, 1921, lectotype, dorsal view; B. Ditto, lateral view; C. Labels pinned with lectotype; D. B. taiwanica Döberl, 2010, holotype, dorsal view; E. Ditto, lateral view; F. Labels pinned with holotype.
Batophila
alishanensis
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
5DB927FF-CBAD-5AEB-A8FB-5CDE62EEEBAF
https://zoobank.org/54992EF9-7381-4B29-91DD-F31FEE2343D3
Batophila acutangula : Kimoto 1989: 269 (part).
Type specimens examined (n = 47).
Holotype ♂ (TARI): Taiwan • Chiayi: Alishan (阿里山), 17–20.VIII.1982, leg. K. C. Chou & C. C. Pan. Paratypes • 9♂, 8♀ (TARI), same data as holotype; 1♀ (KMNH), same locality, 5.V.1971, leg. K. Kamiya; • 1♂, 1♀ (KMNH), same but with “25.V.1971"; 1♂, 2♀♀ (KMNH), same but with “26.V.1971"; • 2♂♂, 2♀♀ (KMNH), same locality, 22–25.VI.1974, leg. M. Owada; • 4♂♂, 9♀♀ (TARI), same locality, 5–9.VIII.1981, leg. L. Y. Chou & S. C. Lin; • 1♂, 1♀ (KMNH), same locality, 7.IX.1986, leg. K. Baba, identified as B. acutangula by Kimoto (1989); • 1♂ (KMNH), same locality, 6.VIII.1990, leg. S. Kimoto; • Chiayi: 1♂, 3♀♀ (NMNS), Lulinshan (鹿林山), 18.V.1991, leg. C. C. Chiang; • Nantou: 1♀ (TARI), Niitakayama (= Yushan, 玉山), 17.VII.1941, leg. S. Miyamoto; • 1♀ (TARI), Tatachia (塔塔加), 16–23.VI.2007, leg. C.-S. Tung.
Diagnosis.
Adults of B. alishanensis sp. nov. are similar to those of B. wusheensis sp. nov., B. houjayi sp. nov. (Figs 10, 12), B. yuae sp. nov., B. jungchani sp. nov., and B. huangi sp. nov. (Fig. 16) in possessing truncate elytral apices. It can be recognized easily by fine punctures on elytra (Fig. 2). The aedeagus of this new species is similar to that of the sympatric species B. houjayi sp. nov. but differs by the wider subapical area (Fig. 3C) [apically narrow aedeagus in B. houjayi sp. nov. (Fig. 11C)].
Habitus of Batophila alishanensis sp. nov. A. Male, paratype, dorsal view; B. Ditto, ventral view; C. Ditto, lateral view; D. Female, paratype, dorsal view; E. Ditto, ventral view; F. Ditto, lateral view.
Batophila alishanensis sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Description.
Male. Length 1.44–1.61 mm, width 0.74–0.80 mm. General color metallic dark bronze (Fig. 2A–C); legs yellowish but hind femora darkened. Antenna (Fig. 3A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.6: 0.5: 0.5: 0.8: 0.6: 0.7: 0.7: 0.6: 0.6: 0.9; ratio of length to width of antennomeres I–XI 2.9: 2.5: 2.3: 2.5: 3.4: 2.5: 2.4: 2.4: 1.9: 1.9: 2.6. Pronotum 1.19–1.25× wider than long; lateral margins slightly rounded, anterolateral angles separated from lateral margins by weak emarginations, slightly narrowed basally, distance between anterolateral angles 1.13–1.17× wider than basal margin. Elytra 1.27–1.30× longer than wide; lateral margins rounded, widest at basal 1/5, apex truncate; dorsoventrally flattened, apex visible in dorsal view; disc with longitudinal lines of fine punctures and with indistinct longitudinal grooves along lines present near base and sides. Tarsomeres I of front and middle legs slightly swollen. Aedeagus (Fig. 3C, D) elongate, 5.8× longer than wide; narrowest at apical 1/3, apically widened towards apical 1/7 and then narrowed, apex widely rounded, basally widened near base; dorsal opening starting from apical 1/10 and basally membranous, tectum composed of two lobes, mostly membranous; slightly curved in lateral view; ventral surface with membranous area narrower than dorsal opening, starting from apical 1/10 to 2/5.
Female (Fig. 2D–F). Length 1.75–1.93 mm, width 0.85–0.96 mm. Antennae similar to males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 3B) 1.0: 0.6: 0.5: 0.5: 0.7: 0.6: 0.7: 0.7: 0.6: 0.6: 0.9; ratio of length to width of antennomere I–XI 2.6: 2.3: 2.4: 2.3: 2.8: 2.2: 2.2: 2.1: 1.8: 1.9: 2.5. Elytra 1.37–1.38× longer than wide; lateral margins rounded, widest at basal 1/3, apex truncate; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of fine punctures and with indistinct longitudinal grooves along lines, reduced in some individuals. Gonocoxae (Fig. 3F) slender, connected with each other at basal 1/5; each gonocoxa with seven long setae and one tiny seta from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 3E) weakly sclerotized apically, with several short setae at apical area, and some tiny setae at apical margin, spiculum extremely elongate. Spermathecal receptaculum (Fig. 3G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Food plants.
Rosaceae: Rubus sp.
Etymology.
This new species is named after its type locality, Alishan (阿里山).
Distribution.
Only known from the abovementioned localities, which are alpine habitats in southern Taiwan (Fig. 4).
Distribution map of Batophila species, solid line: 1000 m a.s.l., broken line: 2000 m a.s.l., black areas: 3000 m a.s.l. Red dots B. alishanensis sp. nov. Purple dots B. taiwanica Döberl Green dots B. yehi sp. nov. Brown dots B. meihuai sp. nov.
Batophila
choui
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
B2C97F06-5801-5200-BB98-28B3C699CEA7
https://zoobank.org/96FCCE48-AD8C-4E36-BFA8-14DCAD085CE2
Batophila acutangula : Kimoto 1971: 269 (part).
Type specimens examined (n = 558).
Holotype ♂ (TARI). Taiwan • Nantou: Meifeng (梅峰), 7–9.V.1981, leg. K. S. Lin & S. C. Lin. Paratypes • Hualien: 1♂, 1♀ (TARI), Tayuling (大禹嶺), 9–16.VI.1980, leg. K. S. Lin & B. H. Chen; • Nantou: 11♂♂, 7♀♀ (TARI), Meifeng (梅峰), 10.V.1979, leg. K. C. Chou; • 1♂, 2♀♀ (TARI), same locality, 18.VII.1979, leg. K. C. Chou; • 6♂♂, 8♀♀ (TARI), same locality, 2–4.VI.1980, leg. L. Y. Chou & C. C. Chen; • 6♂♂, 9♀♀ (TARI), same locality, 8.VI.1980, leg. K. S. Lin & B. H. Chen; • 3♂♂, 9♀♀ (TARI), same locality, 26.VIII.1980, leg. K. S. Lin & C. H. Wang; • 1♂, 1♀ (TARI), same locality, 5–9.X.1980, leg. C. C. Chen & C. C. Chien; • 25♂♂, 18♀♀ (TARI), same locality, 7–9.V.1981, leg. K. S. Lin & S. C. Lin; • 49♂♂, 25♀♀ (TARI), same locality, 24–26.VI.1981, leg. K. S. Lin & W. S. Tang; • 6♂♂, 6♀♀ (TARI), same locality, 28–29.VIII.1981, leg. L. Y. Chou & S. C. Lin; • 6♂♂, 1♀ (TARI), same locality, 22.V.1982, leg. L. Y. Chou; • 33♂♂, 22♀♀ (TARI), same locality, 15.VII.1982, S. C. Lin & C. N. Lin; • 9♂♂, 13♀♀ (TARI), same locality, 31.VIII.–2.IX.1982, leg. L. Y. Chou & K. C. Chou; • 45♂♂, 25♀♀ (TARI), same locality, 4–7.X.1982, leg. K. C. Chou; 1♀ (TARI), same locality, 19–21.IV.1983, leg. K. C. Chou & S. P. Huang; • 5♂♂, 1♀ (TARI), same locality, 30.VII.1983, leg. L. Y. Chou; • 1♂, 2♀♀ (TARI), same locality, 8–11.V.1984, leg. K. C. Chou & C. C. Pan; • 1♂ (TARI), same locality, 23.VII.1984, leg. K. S. Lin; • 1♀ (NMNS), same locality, 9.I.–6.II.2007, leg. C. S. Lin & W. T. Yang, Malaise trap; • 2♂♂, 4♀♀ (TARI), same locality, 20.IV.2025, leg. C.-F. Lee; • 17♂♂, 5♀♀ (TARI), Sungkang (松崗), 15–17.VIII.1984, leg. K. C. Chou; • 10♂♂, 5♀♀ (TARI), same locality, 13–15.IX.1984, leg. K. S. Lin & S. C. Lin; • 2♂♂, 2♀♀ (KMNH), same locality, 2.VIII.1990, leg. S. Kimoto, of which one male and one female identified as B. acutangula by Kimoto in 1990; • 1♂, 1♀ (TARI), same locality, 20.IV.2011, leg. C.-F. Lee; • 4♂, 2♀ (KMNH), Sungkang (松崗) – Tsifen (sic!) (翠峰), 29.VI.1965, leg. S. Kimoto, identified as B. acutangula by Kimoto (1971); • 1♀ (NHMUK), (near Sungkang, 松崗) sheep farm, 24°03.121'N, 121°09.643'E, 1916 m, 7.VIII.2008, leg. M. V. L. Barclay & Mendel; • 4♂♂, 1♀ (TARI), Tsuifeng (翠峰), 21.VI.1979, leg. K. S. Lin & B. H. Chen; • 4♀♀ (TARI), same locality, 3.VI.1980, leg. L. Y. Chou & C. C. Chen; • 45♂♂, 28♀♀ (TARI), same locality, 25–27.VI.1981, leg. K. S. Lin & W. S. Tang; • 10♂♂, 3♀♀ (TARI), same locality, 1–3.VIII.1981, leg. T. Lin & W. S. Tang; • 1♂, 1♀ (TARI), same locality, 27.VIII.1981, leg. L. Y. Chou & S. C. Lin; • 1♂, 1♀ (TARI), same locality, 23.V.1982, leg. L. Y. Chou; • 4♂♂, 4♀♀ (TARI), same locality, 1–3.IX.1982, leg. L. Y. Chou & K. C. Chou; • 3♀♀ (TARI), same locality, 20.IV.1983, leg. K. C. Chou & S. P. Huang; • 1♂ (TARI), same locality, IV.1984, Malaise trap, leg. K. S. Lin & K. C. Chou; • 1♂ (TARI), same but with “V.1984"; • 5♂♂ (TARI), same locality, 5.VIII.1984, leg. K. S. Lin; • 3♂♂ (TARI), same locality, 15–16.VIII.1984, leg. K. C. Chou; • 1♂, 5♀ (TARI), same locality, 12–14.IX.1984, leg. K. S. Lin & S. C. Lin; • 1♀ (TARI), Yu-shih (幼獅), 4.VIII.1981, leg. T. Lin & W. S. Tang; • Taichung: 1♂ (TARI), Anmashan (鞍馬山), 6–7.VII.1979, leg. L. Y. Chou; • 1♀ (TARI), same but with “6–9.VII.1979"; • 1♀ (NMNS), same locality, 1.V.1990, leg. C. C. Chiang; • 2♀♀ (TARI), same locality, 21.IV.2010, leg. C.-F. Lee; • 8♀♀ (TARI), Chiapaotai (佳保台)—Liming (黎明), 4.VI.1942, leg. S. Issiki.
Diagnosis.
Adults of B. choui sp. nov., B. chungi sp. nov., and B. tsoui sp. nov. are recognized by their strongly apically narrowed elytra, and divergent elytral apex. They differ by the presence of convex elytra and elytral apices not visible in dorsal view in both sexes (Fig. 5C, E) [flattened elytra in males but convex elytra in females of B. chungi sp. nov. and B. tsoui sp. nov. (Fig. 21C, E)], and parallel-sided aedeagus (Fig. 6C) [widened apex of aedeagus in B. chungi sp. nov. (Fig. 8C) and B. tsoui sp. nov. (Fig. 22C)].
Habitus of Batophila choui sp. nov. A. Male, paratype, dorsal view; B. Ditto, ventral view; C. Ditto, lateral view; D. Female, paratype, dorsal view; E. Ditto, ventral view; F. Ditto, lateral view.
Batophila choui sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Description.
Male. Length 1.99–2.18 mm, width 0.83–0.90 mm. General color metallic dark bronze (Fig. 5A–C); legs yellowish but femora of hind legs darkened. Antenna (Fig. 6A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.6: 0.6: 0.6: 0.8: 0.7: 0.7: 0.7: 0.7: 0.7: 0.9; ratio of length to width of antennomeres I–XI 2.8: 2.3: 2.6: 2.9: 3.4: 2.6: 2.8: 2.6: 2.1: 2.1: 2.3. Pronotum 1.13–1.14× wider than long; lateral margins slightly rounded, anterolateral angles separated from lateral margins by weak emargination, slightly and basally narrowed, distance between anterolateral angles 1.11–1.17× wider than basal margin. Elytra 1.52–1.54× longer than wide; lateral margins rounded, widest at basal 1/5, apically and strongly narrowed, apex truncate but diverge; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of extremely coarse punctures and with distinct longitudinal grooves along punctures, punctures and grooves apically abbreviated from apical 1/3. Tarsomeres I of front and middle legs slightly swollen. Aedeagus (Fig. 6C, D) elongate, 6.0× longer than wide; parallel-sided, apex widely rounded; dorsal opening starting from apical 1/10–1/3, tectum composed of three lobes, median lobe more basal relative to lateral lobes, apical margin truncate, mostly membranous; slightly curved in lateral view, apex moderately curved; ventral surface with membranous area wider than dorsal opening, starting from apical 1/12–1/2.
Female (Fig. 5D–F). Length 2.20–2.38 mm, width 0.96–1.02 mm. Antennae similar to males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 6B) 1.0: 0.6: 0.6: 0.6: 0.8: 0.7: 0.7: 0.7: 0.7: 0.7: 0.9; ratio of length to width of antennomeres I–XI 2.5: 2.1: 2.7: 3.0: 3.5: 2.7: 2.4: 2.1: 2.0: 1.9: 2.5. Elytra 1.53–1.62× longer than wide; lateral margins rounded, widest at basal 1/5, apically and strongly narrowed, apex truncate but diverge; dorsoventrally convex, apex curved strongly downward in lateral view; disc with longitudinal lines of extremely coarse punctures and with distinct longitudinal grooves along punctures, punctures and grooves apically abbreviated from apical 1/3. Gonocoxae (Fig. 6F) slender, connected at basal 1/5; each gonocoxa with seven long setae and one tiny seta from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 6E) weakly sclerotized apically, with several short setae at apical area, and some tiny setae at apical margin, spiculum extremely elongate. Spermathecal receptaculum (Fig. 6G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Food plants.
Rosaceae: Rubus corchorifolius L. f.
Etymology.
This new species is named after late Dr. Liang-Yih Chou (周樑鎰), who worked as Researcher at the TARI, conducted insect diversity projects during 1979–1984 and collected most of type series of this new species.
Distribution.
This species is widespread in mountainous areas of central Taiwan (Fig. 7).
Distribution map of Batophila species, solid line: 1000 m a.s.l., broken line: 2000 m a.s.l., black areas: 3000 m a.s.l. Red dots B. tsoui sp. nov. Purple dots B. choui sp. nov. Green dot B. chungi sp. nov.
Batophila
chungi
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
0DBAFE99-18D4-5588-96A6-6B45D5A48BD6
https://zoobank.org/C28A03E4-F0BE-4E2E-B7D6-6E244EDEB403
Type specimens examined (n = 20).
Holotype ♂ (TARI): Taiwan • Pingtung: Tahanshan (大漢山), 26.II.2025, leg. Y.-T. Chung. Paratypes • 4♂♂, 6♀♀ (TARI), same data as holotype; • 1♂ (TARI), same but with “12.III.2025"; • 1♀ (TARI), same locality, 6.II.2008, leg. S.-F. Yu; • 1♂ (TARI), same locality, 14.VIII.2011, leg. Y.-T. Wang; • 1♀ (TARI), same locality, Malaise trap, 3.IV.–2.V.2020, leg. Y.-C. Chiu; • 3♂♂, 2♀♀ (TARI), same locality, 8.III.2025, leg. J.-C. Chen.
Diagnosis.
Adults of B. chungi sp. nov., B. tsoui sp. nov., and B. choui sp. nov. are recognized by their strongly apically narrowed elytra, and divergent elytral apices, but B. chungi sp. nov. and B. tsoui sp. nov. differ in possessing flattened elytra in males and convex elytra in females (Fig. 21C, E) [convex elytra and elytral apex not visible in dorsal views in both sexes of B. choui sp. nov. (Fig. 5C, E)], and widened apex of aedeagus (Fig. 8C) [parallel-sided aedeagus in B. choui sp. nov. (Fig. 6C)]. Adults of B. chungi sp. nov. are not separable from those B. tsoui sp. nov. by external morphology but the aedeagus of B. chungi sp. nov. (Fig. 8C) is much wider than that of B. tsoui sp. nov. (Fig. 22C).
Batophila chungi sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Description.
Male. Length 1.95–2.32 mm, width 0.79–0.91 mm. General color metallic dark bronze; legs yellowish but femora of hind legs darkened. Antenna (Fig. 8A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.6: 0.6: 0.6: 0.8: 0.7: 0.7: 0.8: 0.7: 0.6: 0.9; ratio of length to width of antennomeres I–XI 2.3: 2.1: 2.7: 2.9: 3.3: 2.6: 2.2: 2.4: 2.0: 1.9: 2.3. Pronotum 1.10–1.12× wider than long; lateral margins slightly rounded, anterolateral angles separated from lateral margins by weak emarginations, slightly narrowed basally, distance between anterolateral angles 1.16–1.20× wider than basal margin. Elytra 1.57–1.60× longer than wide; lateral margins rounded, widest at basal 1/5, apically and strongly narrowed, apex truncate but divergent; dorsoventrally flattened, apex visible in dorsal view; disc with longitudinal lines of extremely coarse punctures and distinct longitudinal grooves along punctures, punctures and grooves apically abbreviated from apical 1/3. Tarsomeres I of front and middle legs slightly swollen. Aedeagus (Fig. 8C, D) elongate, 4.4× longer than wide; widest at apical 1/5, apically narrowed towards apex, apex widely rounded, basally widened near apical 2/5, then parallel-sided near base; dorsal opening starting from apical 1/7–9/20, tectum composed of three lobes, median lobe more basal relative to lateral lobes, apical margin truncate, mostly membranous; slightly curved in lateral view, apex moderately curved; ventral surface with membranous area as wide as dorsal opening, starting from apical 1/6–1/2.
Female. Length 2.11–2.45 mm, width 0.91–1.03 mm. Antennae similar to males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 8B) 1.0: 0.6: 0.6: 0.6: 0.8: 0.7: 0.8: 0.7: 0.7: 0.7: 1.0; ratio of length to width of antennomeres I–XI 2.6: 2.0: 2.7: 2.7: 3.6: 2.7: 2.3: 2.3: 2.1: 2.0: 2.9. Elytra 1.49–1.57× longer than wide; lateral margins rounded, widest at basal 1/5, apex truncate but divergent; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of extremely coarse punctures and with distinct longitudinal grooves along punctures, punctures and grooves apically abbreviated from apical 1/3. Gonocoxae (Fig. 8F) slender, connected at basal 1/5; each gonocoxa with seven long setae and one tiny seta from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 8E) weakly sclerotized apically, with several short setae at apical area, and some tiny setae at apical margin, spiculum extremely elongate. Spermathecal receptaculum (Fig. 8G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Food plants.
Rosaceae: Rubus formosensis Kuntze; R. taitoensis var. aculeatiflorus (Hayata) H. Ohashi & C. F. Hsieh (Fig. 9A, B).
Field photographs of Batophila species A. Rubus taitoensis var. aculeatiflorus (Rosaceae); B. Adult of B. chungi sp. nov. on underside of leaf of R. taitoensis var. aculeatiflorus; C. Adult of B. houjayi sp. nov. resting on underside of leaf of Persicaria chinense (Polygonaceae); D. Adults of B. tsoui sp. nov. resting on underside of leaf of R. wallichianus (Rosaceae); E. Otanthera scaberrima (Melastomataceae); F. Adults of B. tsoui sp. nov. feeding on underside of leaf of O. scaberrima
Etymology.
This new species is named for Yi-Ting Chung (鍾奕霆), the first member of TCRT to collect specimens.
Distribution.
Only known from the type locality in southern Taiwan (Fig. 7).
Batophila
houjayi
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
D2119E9C-F283-50E2-8205-3CAEC537D261
https://zoobank.org/3A0988F9-87A4-46FA-A4A5-FF7719FA8A03
Batophila acutangula : Kimoto 1989: 269 (part). Batophila yangweii : Chûjô 1937: 54 (part).
Type specimens examined (n = 868).
Holotype ♂ (TARI). Taiwan • Nantou: Tatachia (塔塔加), 21.IX.2009, leg. C.-F. Lee. Paratypes • 4♀♀ (TARI), same data as holotype; Taiwan • Chiayi: 1♂, 3♀♀ (KMNH), Alishan (阿里山), 21.V.1971, leg. K. Kamiya; • 4♂♂, 2♀♀ (KMNH), same but with “25.V.1971"; • 1♀ (KMNH), same but with “26.V.1971"; • 1♀ (KMNH), same locality, 22–25.VI.1974, leg. M. Owada; • 45♂♂, 30♀♀ (TARI), same locality, 5–9.VIII.1981, leg. L. Y. Chou & S. C. Lin; • 129♂♂, 56♀♀ (TARI), same locality, 17–20.VIII.1982, leg. K. C. Chou & C. C. Pan; • 1♀ (TARI), same locality, 25.IV.2009, leg. H.-J. Chen; • 3♂♂ (NMNS), same locality, 4.X.1988, leg. K. S. Huang; • 28♂♂, 10♀♀ (KMNH), same locality, 6.VIII.1990, leg. S. Kimoto, of which one male identified as B. acutangula by Kimoto in 1990; • 3♂♂, 1♀ (TARI), same locality, 12.V.2011, leg. C.-F. Lee; • 8♂♂, 4♀♀ (NHMUK), SE of Alishan, 23°28'20"N, 120°51'E, 2300 m, 6–8.XI.2008, leg. L. Dembický; • 2♂♂, 1♀ (TARI), Tzuchung (自忠), 21.IX.2009, leg. M.-H. Tsou; • 2♂♂ (TARI), same locality, 22.II.2016, leg. Y.-T. Wang; • Hualien: 16♂♂, 17♀♀ (TARI), Tayuling (大禹嶺), 9–16.VI.1980, leg. K. S. Lin & B. H. Chen; • 2♂♂, 2♀♀ (TARI), same locality and collectors, 10–16.VI.1980, Malaise trap; • 4♂♂, 8♀♀ (TARI), same locality, 12–15.IX.1980, leg. K. S. Lin & C. H. Wang; • 1♂, 1♀ (NHMUK), SW above Tayuling, 24°10'N, 121°17'30E, 2950 m, 31.X.2008, leg. L. Dembický; • Ilan: 1♀ (TARI), Sikikun (= Sunchitsun, 四季村), 11.VII.1933, leg. M. Chujo, identified as B. yangweii by Chûjô (1937); • 1♂, 2♀♀ (TARI), same locality (= Ssuchi), 7.VII.2009, leg. H.-J. Chen; • 1♂ (TARI), same but with “19.V.2010"; • 1♂ (TARI), Taiheizan (= Taipingshan, 太平山), 9.VII.1933, leg. M. Chujo, identified as B. yangweii by Chûjô (1937); • 5♂♂, 5♀♀ (TARI), same locality, 7.VII.1940, leg. R. Matuda; • Kaohsiung: 1♂, 1♀ (KMNH), Tienchi (天池), 2.V.1986, leg. K. Baba, identified as B. acutangula by Kimoto (1989); • Nantou: 1♀ (TARI), Huakang (華崗), 14.IX.2010, leg. C.-F. Lee; • 1♂, 1♀ (NMNS), Kahoershan (卡賀爾山), 7.V.1992. leg. W. T. Yang; • 1♂ (NMNS), Lienhuachih (蓮華池), 9.IV.–19.V.1998, leg. C. S. Lin & W. T. Yang; • 1♂ (KMNH), Lushan Wenchuan (廬山溫泉), 6.VI.1976, leg. H. Makihara; • 7♂♂, 16♀♀ (NMNS), Nanhuashan (南華山), 6.V.1992, leg. Yang & Huang; • 1♂ (NHMUK), (near Sungkang, 松崗) sheep farm, 24°03.121'N, 121°09.643'E, 1916 m, 7.VIII.2008, leg. M. V. L. Barclay & Mendel; • 1♂, 2♀♀ (TARI), Tatachia (塔塔加), 29.X.2009, leg. H. Lee; • 1♂ (TARI), same locality, 17.XI.2009, leg. C.-F. Lee; • 1♀ (TARI), same locality, 18.XI.2009, leg. H. Lee; • 3♂♂, 14♀♀ (TARI), Tsuifeng (翠峰), 21.VI.1979, leg. K. S. Lin & B. H. Chen; • 8♀♀ (TARI), same locality, 3.VI.1980, leg. L. Y. Chou & C. C. Chen; • 4♂♂, 35♀♀ (TARI), same locality, 25–27.VI.1981, leg. K. S. Lin & W. S. Tang; • 32♂♂, 90♀♀ (TARI), same locality, 1–3.VIII.1981, leg. T. Lin & W. S. Tang; • 1♂, 1♀ (TARI), same locality, 8.XI.1981, leg. S. C. Lin & W. S. Tang; • 2♂♂, 2♀♀ (TARI), same locality, 23.V.1982, leg. L. Y. Chou; • 33♂♂, 66♀♀ (TARI), same locality, 1–3.IX.1982, leg. L. Y. Chou & K. C. Chou; • 3♀♀ (TARI), same locality, 20.IV.1983, leg. K. C. Chou & S. P. Huang; • 1♀ (TARI), same locality, IV.1984, Malaise trap, leg. K. S. Lin & K. C. Chou; • 6♂♂, 17♀♀ (TARI), same locality, 9.V.1984, leg. K. C. Chou & C. C. Pan; • 6♂♂, 29♀♀ (TARI), same locality, 5.VIII.1984, leg. K. S. Lin; • 13♂♂, 26♀♀ (TARI), same locality, 12–14.IX.1984, leg. K. S. Lin & S. C. Lin; • 4♂♂, 1♀ (NMNS), Yuanfeng (鳶峰), 9.III.–9.IV.1998, leg. C. S. Lin & W. T. Yang, Malaise trap; • 5♂♂, 5♀♀ (NMNS), same but with “12.III.–9.IV.2002"; • 2♂♂, 2♀♀ (NMNS), same but with “9.IV.–7.V.2002"; • 2♀♀ (NMNS), same but with “11.VI.–9.VII.2002"; • 1♂, 2♀♀ (NMNS), same but with “17.IV.–7.V.2003"; • 1♀ (NMNS), same but with “11.VI.–8.VII.2003"; • 3♀♀ (NMNS), same but with “11.V.–13.VII.2004"; • 1♀ (NMNS), same but with “13.III.–10.IV.2007"; • 6♀♀ (NMNS), Yunhaipaohsienso (雲海保線所), 4.V.1992, leg. W. T. Yang; • 1♀ (KMNH), Yushan (玉山), 19.V.1981, leg. N. Ito, identified as B. acutangula by Kimoto in 1987.
Diagnosis.
Adults of B. houjayi sp. nov. are not separable from those of B. wusheensis sp. nov., B. yuae sp. nov., B. jungchani sp. nov., and B. huangi sp. nov. that are characterized by truncate elytral apices based on external morphology except for the aedeagus (see below). However, these species can be recognized by their allopatric distributions [B. houjayi sp. nov. inhabits high mountains in Chiayi, Ilan, Hualien, and Nantou counties, B. wusheensis sp. nov. in lowlands of Nantou County, B. yuae sp. nov. in lowlands of Taipei and New Taipei Cities, and Ilan County, B. jungchani sp. nov. in high mountains of Taichung and Miaoli counties, B. huangi sp. nov. in lowlands of Miaoli County and high mountains in Hsinchu and Taoyuan counties (Fig. 13)]. Aedeagal shapes are diagnostic [widely rounded apex of aedeagus in B. houjayi sp. nov. (Fig. 11C), apically tapering aedeagus from apical 1/5 in B. wusheensis sp. nov. (Fig. 23C), rounded apex of aedeagus with truncate process at middle of apical margin in B. yuae sp. nov. (Fig. 26C), subapically tapering apex of aedeagus in B. jungchani sp. nov. (Fig. 15C), and rounded apex of aedeagus with small, rounded process at middle of apical margin in B. huangi sp. nov. (Fig. 14C)].
Description.
Male. Length 1.63–1.92 mm, width 0.82–0.94 mm. General color metallic dark bronze (Fig. 10A–C); antennae yellowish brown but six apical antennomeres darker; legs yellowish but femora of hind legs darkened. Antenna (Fig. 11A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.6: 0.5: 0.5: 0.6: 0.6: 0.6: 0.6: 0.6: 0.6: 0.8; ratio of length to width of antennomeres I–XI 2.6: 2.1: 2.2: 2.3: 2.6: 2.2: 2.0: 1.8: 1.7: 1.6: 2.3. Pronotum 1.22–1.30× wider than long; lateral margins slightly rounded, anterolateral angles separated from lateral margins by weak emarginations, slightly narrowed basally, distance between anterolateral angles 1.10–1.17× wider than basal margin. Elytra 1.28–1.34× longer than wide; lateral margins rounded, widest at basal 1/5, apex truncate; dorsoventrally flattened, apex visible in dorsal view; disc with longitudinal lines of coarse punctures, and distinct longitudinal grooves along punctures apically abbreviated, ridges present between longitudinal grooves and apically abbreviated from basal 1/3. Tarsomeres I of front and middle legs slightly swollen. Aedeagus (Fig. 11C, D) elongate, 5.7× longer than wide; lateral margins subapically parallel, then slightly widened basally, widest at basal 1/3, apex widely rounded; dorsal opening starting from apical 1/10–2/5, tectum membranous; moderately curved in lateral view; ventral surface with membranous area narrower than dorsal opening, starting from apical 1/20–1/3.
Females (Fig. 10D–F). Length 1.79–2.21 mm, width 0.96–1.10 mm. Antennae similar to males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 11B) 1.0: 0.6: 0.5: 0.5: 0.6: 0.5: 0.6: 0.6: 0.6: 0.6: 0.8; ratio of length to width of antennomeres I–XI 2.9: 2.2: 2.5: 2.2: 3.0: 2.3: 2.2: 1.8: 1.8: 1.7: 2.4. Elytra 1.26–1.38× longer than wide; lateral margins rounded, widest at basal 1/5, apex truncate; dorsoventrally convex, elytral apex not visible in dorsal view; disc with longitudinal lines of fine punctures, and distinct longitudinal grooves along punctures apically abbreviated from apical 1/3, ridges present between 2^nd^ and 3^rd^, 4^th^ and 5^th^ longitudinal grooves and apically abbreviated from basal 1/3. Gonocoxae (Fig. 11F) slender, connected from basal 1/5 to base; each gonocoxa with seven long setae and one tiny seta from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 11E) weakly sclerotized apically, with several short setae at sides of apex, and some tiny setae at sides of apical margin, spiculum extremely elongate. Spermathecal receptaculum (Fig. 11G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Habitus of Batophila houjayi sp. nov., from Tsuifeng (翠峰) A. Male, paratype, dorsal view; B. Ditto, ventral view; C. Ditto, lateral view; D. Female, paratype, dorsal view; E. Ditto, ventral view; F. Ditto, lateral view.
Batophila houjayi sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Variation.
Longitudinal ridges on elytra present in most individuals from Alishan (Fig. 12) but absent in specimens from other areas.
Habitus of Batophila houjayi sp. nov., from Alishan (阿里山) A. Male, paratype, dorsal view; B. Ditto, ventral view; C. Ditto, lateral view; D. Female, paratype, dorsal view; E. Ditto, ventral view; F. Ditto, lateral view.
Distribution map of Batophila species, solid line: 1000 m a.s.l., broken line: 2000 m a.s.l., black areas: 3000 m a.s.l. Red dots B. yuae sp. nov. Blue dots B. wusheensis sp. nov. Green dot B. jungchani sp. nov. Pink dots B. houjayi sp. nov. Purple dots B. huangi sp. nov.
Batophila huangi sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Food plants.
Rosaceae: Rubus lambertianus Ser. and R. croceacanthus H. Lév.; Polygonaceae: Persicaria thunbergia Sieb. et Zucc. and P. chinense L. (Fig. 9C).
Etymology.
This new species is named for Hou-Jay Chen (陳厚潔), the first member of TCRT to collect specimens.
Distribution.
This species is widespread in mountainous areas of south and central Taiwan (Fig. 13).
Batophila
huangi
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
C4F8F5ED-3220-52F5-80B4-713F6D5456B7
https://zoobank.org/3D4EAF0C-A47A-43F0-94D2-9AE69C825FBD
Batophila yangweii : Chûjô 1937: 54 (part).
Type specimens examined (n = 22).
Holotype ♂ (NMNS). Taiwan • Miaoli: Taian (泰安), 20.XII.1989, leg. K. W. Huang. Paratypes • 8♂♂, 3♀♀ (NMNS), same data as holotype; Hsinchu: • 2♂♂ (TARI), Talulintao (大鹿林道), 17.II.2008, leg. M.-H. Tsou; 1♂ (TARI), Wufeng (五峰), 14–16.VII.1982, leg. K. C. Chou & C. C. Pan; • 1♂ (NMNS), same locality, 21.XII.1989, leg. K. W. Huang; • Taoyuan: 1♂ (TARI), Kayahara (= Hsuanyuan, 萱源), 23.VII.1929, leg. Y. Miwa, identified as B. yangweii by Chûjô (1937); • 1♂, 1♀ (TARI), Lalashan (拉拉山), 8.III.2009, leg. H. Lee; • 1♂ (TARI), same but with “leg. H.-J. Chen”; • 1♀ (TARI), same but with “leg. C.-F. Lee”; • 1♀ (NMNS), Upper Plain (sic!) (= Balung, 上巴陵), 24°41'12.1"N, 121°23'39.3"E, 600 m, 11.IV.1998, leg. Miller, Stange, Wang.
Diagnosis.
Adults of B. huangi sp. nov. are not separable from those of B. houjayi sp. nov., B. wusheensis sp. nov., B. yuae sp. nov., and B. jungchani sp. nov. that are characterized by truncate elytral apices based on external morphology (Figs 10, 12) except for the aedeagus (see below). However, these species can be recognized by their allopatric distributions [B. huangi sp. nov. inhabits lowlands in Miaoli County and high mountains in Hsinchu and Taoyuan counties, B. jungchani sp. nov. in high mountains in Taichung and Miaoli counties, B. yuae sp. nov. in lowlands of Taipei and New Taipei Cities, and Ilan County, B. wusheensis sp. nov. in lowlands of Nantou County, B. houjayi sp. nov. at high mountains in Chiayi, Ilan, Hualien, and Nantou counties (Fig. 13)]. Aedeagal shapes are diagnostic [rounded apex of aedeagus with small, rounded process at middle of apical margin in B. huangi sp. nov. (Fig. 14C), apically tapering apex of aedeagus from apical 1/10 in B. jungchani sp. nov. (Fig. 15C), rounded apex of aedeagus with truncate process at middle of apical margin in B. yuae sp. nov. (Fig. 26C), apically tapering aedeagus from apical 1/5 in B. wusheensis sp. nov. (Fig. 23C), and widely rounded apex of aedeagus in B. houjayi sp. nov. (Fig. 11C)].
Description.
Male. Length 1.52–1.75 mm, width 0.77–0.86 mm. General color metallic dark bronze; antennae yellowish brown but six apical antennomeres darker; legs yellowish but femora of hind legs darkened. Antenna (Fig. 14A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.6: 0.6: 0.6: 0.7: 0.6: 0.8: 0.7: 0.6: 0.6: 0.9; ratio of length to width of antennomeres I–XI 2.4: 2.5: 2.5: 2.5: 3.2: 2.5: 2.7: 2.1: 1.8: 1.8: 2.7. Pronotum 1.18–1.25× wider than long; lateral margins slightly rounded, anterolateral angles separated from lateral margins by weak emarginations, slightly narrowed basally, distance between anterolateral angles 1.17–1.18× wider than basal margin. Elytra 1.35–1.37× longer than wide; lateral margins rounded, widest at basal 1/5, apex truncate; dorsoventrally flattened, apex visible in dorsal view; disc with longitudinal lines of coarse punctures, and indistinct longitudinal grooves along punctures, lacking ridges present between longitudinal grooves. Tarsomeres I of front and middle legs slightly swollen. Aedeagus (Fig. 14C, D) elongate, 5.3× longer than wide; lateral margins basally and slightly widened, widest near base, apex widely rounded and with small rounded process at middle of apical margin; dorsal opening starting from apical 1/10, basally weakly sclerotized; tectum composed of three lobes, median lobe more ventral relative to lateral lobes and apical margin truncate, mostly membranous; moderately curved subapically and medially in lateral view; ventral surface with membranous area same width and height as dorsal opening, starting from apical 1/10–1/2.
Female. Length 1.97–2.19 mm, width 1.00–1.06 mm. Antennae similar to males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 14B) 1.0: 0.6: 0.5: 0.5: 0.6: 0.6: 0.6: 0.6: 0.6: 0.6: 0.8; ratio of length to width of antennomeres I–XI 3.2: 2.3: 2.6: 2.6: 3.0: 2.6: 2.3: 2.1: 2.1: 1.8: 2.6. Elytra 1.36–1.39× longer than wide; lateral margins rounded, widest at basal 1/5, apex truncate; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of coarse punctures, and indistinct longitudinal grooves along punctures, lacking ridges present between longitudinal grooves. Gonocoxae (Fig. 14F) slender, connected from basal 1/5 to base; each gonocoxa with seven long setae and one tiny seta from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 14E) weakly sclerotized apically, with several short setae at sides of apex, and some tiny setae at sides of apical margin, spiculum extremely elongate. Spermathecal receptaculum (Fig. 14G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Food plants.
Rosaceae: Rubus sp.
Etymology.
This new species is named for Dr. Kun-Wei Huang (黃坤煒), who was a former research scientist at the NMNS and collected most of the type specimens.
Distribution.
This species is widespread in mountainous areas of northwestern Taiwan (Fig. 13).
Batophila
jungchani
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
3BC830BB-A2E5-5658-AA33-1797188417E2
https://zoobank.org/0B81AB2D-59CC-4E7E-9152-21A4EA66F560
Type specimens examined (n = 36).
Holotype ♂ (NMNS). Taiwan • Taichung: Anmashan (鞍馬山), 3.V.1992, leg. C. Y. Li. Paratypes • 3♂♂, 12♀♀ (NMNS), same data as holotype; • 1♂, 1♀ (NMNS), same locality, 3.V.1990, leg. C. C. Chiang; • 5♀♀ (TARI), same locality, 21.IV.2010, leg. C.-F. Lee; • Miaoli: 1♂ (TARI), Hsiaopangchih (小胖池), 28.VIII.2021, leg. Y.-F. Hsu; • 4♂♂, 4♀♀ (TARI), same but with “16.XI.2021–21.IV.2022"; • Taichung: 2♂♂ (TARI), Hsuehshan (雪山), 24°23'15"N, 121°11'55"E, 29.IV.–28.VI.2012, leg. L.-P. Hsu; • 2♀♀ (TARI), Tahsuehshan (大雪山), 18.IV.2011, leg. J.-C. Chen.
Diagnosis.
Adults of B. jungchani sp. nov. are not separable from those of B. houjayi sp. nov., B. wusheensis sp. nov., B. yuae sp. nov., and B. huangi sp. nov. that are characterized by truncate elytral apices based on external morphology (Figs 10, 12) except for the aedeagus (see below). However, these species can be recognized by their allopatric distributions [B. jungchani sp. nov. inhabits high mountains in Taichung and Miaoli counties, B. yuae sp. nov. in lowlands of Taipei and New Taipei Cities, and Ilan County, B. wusheensis sp. nov. in lowlands of Nantou County, B. houjayi sp. nov. in high mountains in Chiayi, Ilan, Hualien, and Nantou counties, B. huangi sp. nov. in lowlands of Miaoli County and high mountains of Hsinchu and Taoyuan counties (Fig. 13)]. Aedeagal shapes are diagnostic [apically tapering aedeagus from apical 1/10 in B. jungchani sp. nov. (Fig. 15C), rounded apex of aedeagus with truncate process at middle of apical margin in B. yuae sp. nov. (Fig. 26C), apically tapering aedeagus from apical 1/5 in B. wusheensis sp. nov. (Fig. 23C), widely rounded apex of aedeagus in B. houjayi sp. nov. (Fig. 11C), and rounded apex of aedeagus with small, rounded process at middle of apical margin in B. huangi sp. nov. (Fig. 14C)].
Description.
Male. Length 1.93–2.01 mm, width 0.94–0.97 mm. General color metallic dark bronze; antennae and legs reddish brown. Antenna (Fig. 15A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.6: 0.6: 0.6: 0.7: 0.7: 0.8: 0.7: 0.7: 0.6: 0.9; ratio of length to width of antennomeres I–XI 2.7: 2.3: 2.6: 2.4: 3.0: 2.7: 2.4: 2.3: 2.2: 2.1: 3.0. Pronotum 1.22–1.25× wider than long; lateral margins slightly rounded, anterolateral angles separated from lateral margins by weak emarginations, slightly narrowed basally, distance between anterolateral angles 1.14–1.15× wider than basal margin. Elytra 1.31–1.35× longer than wide; lateral margins rounded, widest at basal 1/5, apex truncate; dorsoventrally flattened, apex visible in dorsal view; disc with longitudinal lines of coarse punctures, and distinct longitudinal grooves along punctures, punctures and grooves apically abbreviated, lacking ridges present between longitudinal grooves. Tarsomeres I of front and middle legs slightly swollen. Aedeagus (Fig. 15C, D) elongate, 6.4× longer than wide; parallel-sided, apically tapering from apical 1/10; moderately curved in lateral view; dorsal opening starting from apical 1/15, basally weakly sclerotized; tectum composed of three lobes, median lobe more ventral relative to lateral lobes and apical margin truncate, mostly membranous; moderately curved subapically and medially in lateral view; ventral surface with membranous area same width and height as dorsal opening, starting from apical 1/15 to 1/3.
Batophila jungchani sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Female. Length 2.20–2.34 mm, width 1.06–1.17 mm. Antennae similar to males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 15B) 1.0: 0.6: 0.5: 0.6: 0.7: 0.6: 0.7: 0.7: 0.7: 0.6: 0.9; ratio of length to width of antennomeres I–XI 2.9: 2.5: 2.6: 3.0: 3.6: 2.8: 2.9: 2.5: 2.2: 2.0: 2.6. Elytra 1.32–1.38× longer than wide; lateral margins rounded, widest at basal 1/5, apex truncate; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of coarse punctures, and indistinct longitudinal grooves along punctures, lacking ridges present between longitudinal grooves. Gonocoxae (Fig. 15F) slender, connected from basal 1/5 to base; each gonocoxa with seven long and one tiny setae from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 15E) weakly sclerotized apically, with several short setae at sides of apex, and some tiny setae at sides of apical margin, spiculum extremely elongate. Spermathecal receptaculum (Fig. 15G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Variation.
Some individuals have reddish-brown elytra, especially from Anmashan (鞍馬山) (Fig. 16).
Habitus of Batophila jungchani sp. nov., female, paratype, from Anmashan (鞍馬山) A. Dorsal view; B. Ventral view; C. Lateral view.
Food plants.
Unknown.
Etymology.
This new species is named for Jung-Chan Chen (陳榮章), the first person to collect specimens.
Distribution.
This species is widespread in alpine areas of central Taiwan (Fig. 13).
Batophila
meihuai
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
AF39B7D6-497A-5AA2-B631-3B1A91E16B74
https://zoobank.org/6CB4AAAB-B575-4854-ACBF-FCA0BD610C15
Type specimens examined (n = 14).
Holotype ♂ (TARI). Taiwan • Ilan: Ssuyuan (思源), 25.IV.2009, leg. C.-F. Lee. Paratypes • 2♂♂, 1♀ (TARI), same data as holotype; • 4♂♂, 4♀♀ (TARI), same locality, 28.IV.2009, leg. M.-H. Tsou; • 1♂♂ (TARI), same locality, 31.VII.2009, leg. H.-J. Chen; • Taichung: 1♂♂ (NMNS), same locality (= Ssuyuanyakou, 思源啞口), 25.VI.2007, leg. P. H. Chan & W. L. Lien. The type locality “Ssuyuan” is located at the border between Ilan County and Taichung County.
Diagnosis.
Adults of B. meihuai sp. nov. are similar to those of B. taiwanica Döberl and B. yehi sp. nov. in possessing convergent elytral apices. However, adults of B. yehi sp. nov. are recognized by their stout antennae, length of antennomeres VI–X 0.5× length of antennomere I (Fig. 15A, B) [> 0.5× in B. taiwanica (Fig. 22A, B) and B. meihuai sp. nov. (Fig. 18A, B)]. Adults of B. meihuai sp. nov. are also characterized by their distinct and sexually dimorphic longitudinal ridges on the elytra (Fig. 17) [indistinct or reduced longitudinal ridges on the elytra in B. taiwanica (Fig. 19) and B. yehi sp. nov. (Fig. 24)]. The aedeagus of these species are diagnostic: truncate apex in B. meihuai sp. nov. (Fig. 18C), rounded apex with narrowly rounded process at middle of apical margin in B. taiwanica (Fig. 22C), and widely rounded apex in B. yehi sp. nov. (Fig. 25C).
Description.
Male. Length 1.64–2.08 mm, width 0.76–0.88 mm. General color metallic dark bronze (Fig. 17A–C); antennae yellowish brown but six apical antennomeres darker; legs yellowish but femora of hind legs apically darkened. Antenna (Fig. 18A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.6: 0.5: 0.5: 0.7: 0.6: 0.7: 0.7: 0.6: 0.6: 0.9; ratio of length to width of antennomeres I–XI 2.5: 2.3: 2.3: 2.2: 3.2: 2.3: 2.4: 2.0: 1.7: 1.6: 2.5. Pronotum 1.19–1.32× wider than long; lateral margins straight, anterolateral angles not separated from lateral margins, slightly narrowed basally, distance between anterolateral angles widest, 1.12–1.14× wider than basal margin. Elytra 1.40–1.56× longer than wide; lateral margins parallel between basal 1/5 to apical 1/3, apex convergent; dorsoventrally flattened, apex visible in dorsal view; disc with longitudinal lines of coarse punctures, and longitudinal grooves along punctures, distinct ridges present between longitudinal grooves, apices of ridges rounded, apically abbreviated. Tarsomeres I of front and middle legs strongly swollen. Aedeagus (Fig. 18C, D) elongate, 6.9× longer than wide; parallel-sided, strongly tapering near apex, apical margin truncate; dorsal opening from apical 1/7 to basal 1/3, tectum membranous; slightly curved in lateral view; ventral surface with membranous area narrower than dorsal opening, starting from apical 1/8 to 1/2.
Habitus of Batophila meihuai sp. nov. A. Male, paratype, dorsal view; B. Ditto, ventral view; C. Ditto, lateral view; D. Female, paratype, dorsal view; E. Ditto, ventral view; F. Ditto, lateral view.
Batophila meihuai sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Female (Fig. 17D–F). Length 1.96–2.16 mm, width 0.90–1.03 mm. Antennae similar to males, but antennomeres VIII–X narrower than those of females, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 18B) 1.0: 0.5: 0.5: 0.5: 0.6: 0.6: 0.6: 0.6: 0.6: 0.6: 0.8; ratio of length to width of antennomeres I–XI 2.8: 2.2: 2.4: 2.4: 3.0: 2.6: 2.3: 2.2: 2.1: 1.9: 2.8. Elytra 1.42–1.43× longer than wide; lateral margins parallel between basal 1/5 to apical 1/3, apex convergent; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of coarse punctures, and longitudinal grooves along punctures, distinct ridges present between longitudinal grooves, apices of ridges acute, apically abbreviated. Gonocoxae (Fig. 18F) slender, connected from basal 1/5 to base; each gonocoxa with seven long setae and one tiny seta from apical 1/6 to apex, subapically slightly curved. Ventrite VIII (Fig. 18E) weakly sclerotized apically, with several short setae at sides of apex, spiculum extremely elongate. Spermathecal receptaculum (Fig. 18G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Food plants.
Rosaceae: Rubus sp.
Etymology.
This new species is named for Mei-Hua Tsou (曹美華), the first member of TCRT to collect specimens.
Distribution.
Only known from the type locality in northeast Taiwan (Fig. 4).
Batophila
taiwanica
Taxon classificationAnimaliaColeopteraChrysomelidae
Döberl, 2010
205D69B6-4FD0-593F-A469-746B9F2787E2
Batophila acutangula : Kimoto 1971: 73 (part); Kimoto 1989: 269 (part). Batophila yangweii : Chûjô 1937: 54 (part).
Type specimen examined.
Holotype (♂, MNHG) (Fig. 1D–F): “TAIWAN Taichung Hsien / Hseuhshan, nr. Hseuhshan- / Tun-Feng 3170 m 11.V.91 / A. Smetana [176] [p, w] // HOLOTYPE / Batophila / taiwanica / det. Döberl 1997 [p, r] // MHNG / ENTO / 0261580 [p, w]”.
Additional materials examined (n = 1346).
Taiwan • Hsinchu: 1♂ (NMNS), Ching Chuan (清泉), 21.XII.1989, leg. K. W. Huang; • 1♂, 1♀ (NMNS), Kuanwu (觀霧), 25–29.IV.1989, leg. C. S. Lin; • 4♀♀ (TARI), same locality, 30.IV.2010, leg. C.-F. Lee; • 2♀♀ (TARI), same but with “leg. M.-H. Tsou; • 1♂ (TARI), Peitelaman (北得拉曼), 26.VI.2008, leg. S.-F. Yu; • 2♀♀ (TARI), Tahunshan (大混山), 24.II.2009, leg. S.-F. Yu; • 1♀ (TARI), same but with “8.IX.2009”; • 4♀♀ (TARI), Talulintao (大鹿林道), 17.II.2008, leg. M.-H. Tsou; • 3♀♀ (TARI), Wufeng (五峰), 8.II.2009, leg. H.-J. Chen; • Hualien: 1♂ (TARI), Karenko (= Hualien, 花蓮), 20.VII.–4.VIII.1919, leg. T. Okuni, identified as B. yangweii by Chûjô (1937); • 2♂♂ (TARI), Kuanyuan (關原), 2.VI.2016, leg. B.-X. Guo; • 1♂ (TARI), same but with “leg. Y.-T. Chung”; • 2♂♂, 6♀♀ (KMNH), Tayuling (大禹嶺), 21.VI.1976, leg. H. Makihara; • 14♂♂, 11♀♀ (TARI), same locality, 9–16.VI.1980, leg. K. S. Lin & B. H. Chen; • 7♂♂, 11♀♀ (TARI), same locality and collectors, 10–16.VI.1980, Malaise trap; • 2♂♂, 5♀♀ (TARI), same locality, 12–15.IX.1980, leg. K. S. Lin & C. H. Wang; • 5♂♂, 1♀ (NMNS), same locality (= Tai Yu Lin), 25.VII.1990, leg. W. C. Chuang; • 1♀ (NHMUK), SW above Tayuling, 24°10'N, 121°17'30E, 2950 m, 31.X.2008, leg. L. Dembický; • Ilan: 3♂♂, 3♀♀ (TARI), Taipingshan (太平山), 8.VII.2008, leg. H.-J. Chen; • 1♂ (TARI), same locality, 19.II.2009, leg. H. Lee; • 1♂, 4♀♀ (TARI), same locality, 30.IV.2009, leg. C.-F. Lee; • 3♂♂, 3♀ (TARI), Tsuifenghu (翠峰湖), 3.VII.2010, leg. M.-H. Tsou; • Miaoli: 1♀ (TARI), Hsuehchien (雪見), 6.XII.2013, leg. W.-B. Yeh; • 3♂♂ (TARI), same but with “23.III.2014”; • Nantou: 1♂, 1♀ (KMNH), Hohuangshan (合歡山), 3.VI.1971, leg. Kanmiya; • 1♀ (NMNS), same locality (Ho Huang Shan), 26.VII.1990, leg. W. C. Chuang; • 5♀♀ (TARI), same locality (= Hehuanshan), 18.V.2009, leg. M.-H. Tsou; • 1♂ (TARI), Huakang (華岡), 26.VII.2010, leg. M.-H. Tsou; • 9♂♂, 27♀♀ (TARI), Meifeng (梅峰), 10.V.1979, leg. K. C. Chou; • 8♀ (TARI), same locality, 18.VII.1979, leg. K. C. Chou; • 12♂♂, 20♀♀ (TARI), same locality, 2–4.VI.1980, leg. L. Y. Chou & C. C. Chen; • 3♂♂, 19♀♀ (TARI), same locality, 8.VI.1980, leg. K. S. Lin & B. H. Chen; • 3♂♂, 12♀♀ (TARI), same locality, 26.VIII.1980, leg. K. S. Lin & C. H. Wang; • 1♀ (TARI), same locality, 5–9.X.1980, leg. C. C. Chen & C. C. Chien; • 13♂♂, 35♀♀ (TARI), same locality, 7–9.V.1981, leg. K. S. Lin & S. C. Lin; • 57♂♂, 59♀♀ (TARI), same locality, 24–26.VI.1981, leg. K. S. Lin & W. S. Tang; • 12♂♂, 16♀♀ (TARI), same locality, 28–29.VIII.1981, leg. L. Y. Chou & S. C. Lin; • 1♂, 1♀ (TARI), same locality, 7.XI.1981, leg. S. C. Lin & W. S. Tang; • 2♂♂, 2♀♀ (TARI), same locality, 22.V.1982, leg. L. Y. Chou; • 50♂♂, 62♀♀ (TARI), same locality, 15.VII.1982, S. C. Lin & C. N. Lin; • 17♂♂, 25♀♀ (TARI), same locality, 31.VIII.–2.IX.1982, leg. L. Y. Chou & K. C. Chou; • 23♂♂, 28♀♀ (TARI), same locality, 4–7.X.1982, leg. K. C. Chou; • 2♀♀ (TARI), same locality, 19–21.IV.1983, leg. K. C. Chou & S. P. Huang; • 4♂♂, 10♀♀ (TARI), same locality, 30.VII.1983, leg. L. Y. Chou; • 1♂, 3♀♀ (NMNS), same locality, 27.II.1992, leg. Y. C. Shiau; • 2♂♂ (NMNS), same locality, 14.XII.2004–11.I.2005, leg. C. S. Lin & W. T. Yang, Malaise trap; • 1♂, 1♀ (NMNS), same but with “11.I.–15.II.2005”; • 1♂ (NMNS), same but with “3.V.–7.VI.2005”; • 1♂, 2♀ (NMNS), same but with “9.I.–6.II.2007”; • 1♂, 3♀♀ (NMNS), same but with “6.II.–13.III.2007”; • 1♀ (NMNS), same but with “13.III.–10.IV.2007”; • 2♀♀ (KMNH), Piluchi (碧綠溪), 5.VIII.1986, leg. K. Baba, identified as B. acutangula by Kimoto (1989); • 5♂♂ (NMNS), same locality, 4.XII.1991, leg. Y. C. Shiau; • 1♂, 2♀♀ (NMNS), Piluhsi For. Res. Stn. (畢祿溪試驗站), 23–24.V.1999, leg. C. W. & L. B. O’Brien; • 1♂, 1♀ (TARI), Sungkang (松崗), 6.VIII.1984, leg. K. S. Lin; • 15♂♂, 43♀♀ (TARI), same locality, 15–17.VIII.1984, leg. K. C. Chou; • 19♂♂, 17♀♀ (TARI), same locality, 13–15.IX.1984, leg. K. S. Lin & S. C. Lin; • 2♂♂, 1♀ (NHMUK), (near Sungkang, 松崗) sheep farm, 24°03.121'N, 121°09.643'E, 1916 m, 7.VIII.2008, leg. M. V. L. Barclay & Mendel; • 3♂♂, 5♀♀ (KMNH), Sungkang (松崗) – Tsifen (sic!) (翠峰), 29.VI.1965, leg. S. Kimoto, identified as B. acutangula by Kimoto (1971); • 2♂♂, 2♀♀ (KMNH), Tsuifeng (翠峰), 26.VII.1971, leg. Y. Miyake; • 8♂♂, 10♀♀ (TARI), same locality, 21.VI.1979, leg. K. S. Lin & B. H. Chen; • 4♂♂, 16♀♀ (TARI), same locality, 3.VI.1980, leg. L. Y. Chou & C. C. Chen; • 77♂♂, 82♀♀ (TARI), same locality, 25–27.VI.1981, leg. K. S. Lin & W. S. Tang; • 11♂♂, 22♀♀ (TARI), same locality, 1–3.VIII.1981, leg. T. Lin & W. S. Tang; • 1♀ (TARI), same locality, 8.XI.1981, leg. S. C. Lin & W. S. Tang; • 5♂♂, 6♀♀ (TARI), same locality, 23.V.1982, leg. L. Y. Chou; • 11♂♂, 8♀♀ (TARI), same locality, 1–3.IX.1982, leg. L. Y. Chou & K. C. Chou; • 1♂, 1♀ (TARI), same locality, 6.X.1982, leg. K. C. Chou; • 1♂, 13♀♀ (TARI), same locality, 20.IV.1983, leg. K. C. Chou & S. P. Huang; • 2♂♂, 18♀♀ (TARI), same locality, IV.1984, Malaise trap, leg. K. S. Lin & K. C. Chou; • 59♂♂, 68♀♀ (TARI), same locality, 9.V.1984, leg. K. C. Chou & C. C. Pan; • 4♂♂, 21♀♀ (TARI), same locality, 5.VIII.1984, leg. K. S. Lin; • 2♂♂, 8♀♀ (TARI), same locality, 15–16.VIII.1984, leg. K. C. Chou; • 3♂♂, 7♀♀ (TARI), same locality, 12–14.IX.1984, leg. K. S. Lin & S. C. Lin; • 28♂♂, 30♀♀ (TARI), Tungpu (東埔), 20–22.VI.1980, leg. C. C. Chen; • 1♀ (TARI), same locality, 25–29.IX.1980, leg. L. Y. Chou & T. Lin; • 1♀ (TARI), same locality, 28.IV.–2.V.1981, leg. T. Lin & C. J. Lee; • 1♂, 1♀ (TARI), same locality, 18–23.XI.1981, leg. T. Lin & W. S. Tang; • 1♂ (TARI), same locality, 19–23.VII.1982, leg. L. Y. Chou & T. Lin; • 2♀♀ (TARI), same locality, 20–24.VI.1983, leg. K. C. Chou & C. Y. Wong; • 2♂♂, 2♀♀ (TARI), same locality, 16–20.IV.1984, leg. K. C. Chou & C. H. Yung; • 1♂ (NMNS), same locality, 31.I.1989, leg. K. W. Huang; • 1♂ (NMNS), Yuanfeng (鳶峰), 19.II.–12.III.2002, leg. C. S. Lin & W. T. Yang, Malaise trap; • Taichung: 2♂♂, 3♀♀ (NMNS), Anmashan (鞍馬山), 3.V.1992, leg. C. Y. Li; • 2♂♂, 2♀♀ (TARI), same locality 21.IV.2010, leg. C.-F. Lee; • 6♀♀ (TARI), Pilu (畢祿), 17.V.2009, leg. C.-F. Lee; • 1♂, 2♀♀ (TARI), Tahsuehshan (大雪山), 18.IV.2011, leg. J.-C. Chen; • Taoyuan: 1♂ (KMNH), Palon (巴陵?), 3.VIII.1986, leg. K. Baba, identified as B. acutangula by Kimoto (1989).
Diagnosis.
Adults of B. taiwanica Döberl are similar to those of B. yehi sp. nov. and B. meihuai sp. nov. in possessing convergent elytral apices. However, adults of B. taiwanica (Fig. 20A, B) and B. meihuai sp. nov. (Fig. 18A, B) have slender antenna, length of antennomeres VI–X > 0.5× antennomere I [stout antenna in B. yehi sp. nov., antennomeres VI–X 0.5× antennomere I (Fig. 25A, B)]. Adults of B. taiwanica (Fig. 19) and B. yehi sp. nov. (Fig. 24) possess indistinct or reduced longitudinal ridges on the elytra which are different from B. meihuai sp. nov. by their distinct and sexually dimorphic longitudinal ridges on the elytra (Fig. 17). The aedeagus of these species are diagnostic: rounded apex with narrowly rounded process at middle of apical margin in B. taiwanica (Fig. 20C), widely rounded apex in B. yehi sp. nov. (Fig. 25C), and truncated apex in B. meihuai sp. nov. (Fig. 18C).
Habitus of Batophila taiwanica Döberl A. Male, dorsal view; B. Ditto, ventral view; C. Ditto, lateral view; D. Female, dorsal view; E. Ditto, ventral view; F. Ditto, lateral view.
Batophila taiwanica Döberl A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Description.
Male. Length 1.52–1.67 mm, width 0.80–0.83 mm. General color metallic dark bronze (Fig. 19A–C); antennae yellowish brown but six apical antennomeres darker; legs yellowish but apical halves of femora of hind legs darkened. Antenna (Fig. 20A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.6: 0.5: 0.6: 0.7: 0.6: 0.7: 0.7: 0.7: 0.6: 0.9; ratio of length to width of antennomeres I–XI 2.7: 2.3: 2.4: 2.7: 3.2: 2.4: 2.2: 2.4: 2.0: 1.8: 2.7. Pronotum 1.21–1.23× wider than long; lateral margins slightly rounded, anterolateral angles separated from lateral margins by weak emarginations, slightly narrowed basally, distance between anterolateral angles 1.04–1.08× wider than basal margin. Elytra 1.35–1.36× longer than wide; lateral margins rounded, widest at basal 1/3, apex rounded but divergent; dorsoventrally flattened, apex visible in dorsal view; disc with longitudinal lines of coarse punctures and with distinct longitudinal grooves along lines. Tarsomeres I of front and middle legs slightly swollen. Aedeagus (Fig. 20C, D) elongate, 5.8× longer than wide; lateral margins basally and slightly widened towards base, apex narrowly rounded; dorsal opening starting from apical 1/7 and basally membranous, tectum composed of three lobes, median lobe more ventral relative to lateral lobes and apical margin truncate, mostly membranous; moderately curved in lateral view; ventral surface with membranous area narrower than dorsal opening, starting from apical 1/10–2/5.
Female (Fig. 19D–F). Length 1.96–2.04 mm, width 1.06–1.13 mm. Antennae similar to males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 20B) 1.0: 0.6: 0.6: 0.5: 0.7: 0.6: 0.7: 0.7: 0.7: 0.7: 1.0; ratio of length to width of antennomeres I–XI 2.8: 2.3: 2.6: 2.6: 3.0: 2.5: 2.5: 2.5: 2.4: 2.0: 3.1. Elytra 1.29–1.36× longer than wide; lateral margins rounded, widest at basal 1/3, apex rounded but divergent; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of coarse punctures and with distinct longitudinal grooves along punctures. Gonocoxae (Fig. 20F) slender, connected at basal 1/5; each gonocoxa with seven long setae and one tiny seta from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 20E) weakly sclerotized apically, with several short setae at sides of apex, and some tiny setae at sides of apical margin, spiculum extremely elongate. Spermathecal receptaculum (Fig. 20G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Food plants.
Rosaceae: Rubus croceacanthus H. Lév.
Distribution.
This species is widespread in mountainous areas of central Taiwan (Fig. 4).
Batophila
tsoui
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
3461572B-B7C4-5F7B-966F-9F903DF32E10
https://zoobank.org/20C50E88-25E5-4D8D-81CA-70F628403ABD
Batophila acutangula : Kimoto 1989: 269 (part). Batophila yangweii : Chûjô 1937: 54 (part).
Type specimens examined (n = 178).
Holotype ♂ (TARI): Taiwan • Kaohsiung: Chuyunshan logging trail (出雲山林道), 24.III.2009, leg. C.-F. Lee. Paratypes. 3♂♂, 7♀♀ (TARI), same data as holotype; Taiwan • Chiayi: 1♂, 1♀ (TARI), Arisan (= Alishan, 阿里山), 10.X.1912, leg. I. Nitobe, both identified as B. yangweii by Chûjô (1937); • 1♀ (TARI), same locality, 2–23.X.1918, leg. J. Sonan, identified as B. yangweii by Chûjô (1937); • 1♀ (TARI), same locality, 10.VI.1940, leg. M. Chujo; • 1♂, 1♀ (KMNH), same locality, 9.IV.1965, leg. Y. Hirashima, the female identified as B. acutangula in 1975; • 1♂, 1♀ (KMNH), same locality, 17.V.1968, leg. B.-S. Chang, both identified as B. acutangula by Kimoto in 1973; • 1♀ (KMNH), same locality, 5.V.1971, leg. K. Kamiya; • 1♀ (KMNH), same but with “21.V.1971”; • 3♀♀ (KMNH), same but with “26.V.1971”, of which one is identified as B. acutangula by Kimoto in 1971 ; • 1♂ (KMNH), same locality, 22–25.VI.1974, leg. M. Owada; • 7♂♂, 4♀♀ (TARI), same locality, 5–9.VIII.1981, leg. L. Y. Chou & S. C. Lin; • 7♂♂, 4♀♀ (TARI), same locality, 17–20.VIII.1982, leg. K. C. Chou & C. C. Pan; • 1♀ (NMNS), Fenchifu (奮起湖), 14.XII.1988, leg. K. W. Huang; • Kaohsiung: 1♀ (TARI), Erchituan (二集團), 8.III.2013, leg. B.-X. Guo; • 1♂ (TARI), Shihshan (石山), 30.XII.2008–6.I.2009, leg. C.-T. Yao; • 4♂♂, 4♀♀ (KMNH), same locality (= Shyk Shan), 25.IV.1986, leg. K. Baba, identified as B. acutangula by Kimoto (1989); • 3♀♀ (TARI), Shihshan logging trail (石山林道), 24.III.2009, leg. M.-H. Tsou; • 10♂♂, 4♀♀ (NMNS), Tengchih (藤枝), 7.IX.1989, leg. K. W. Huang; • 1♀ (NMNS), same locality, 21–24.XI.1995, leg. M. L. Chan; • 1♂ (TARI), same locality, 18.II.2007, leg. S.-F. Yu; • 1♂ (TARI), same locality, 2–5.VI.2008, leg. C.-F. Lee; • 4♀♀ (TARI), same locality, 9.XI.2013, leg W.-C. Liao; • 1♀ (KMNH), Tienchi (天池), 2.VI.1986, leg. K. Baba, identified as B. acutangula by Kimoto (1989); • 1♂, 5♀♀ (NMNS), Tona forest road (多納林道), 28.IV.1998, leg. M. L. Chan; • 1♀ (TARI), same locality (= Tonalintao), 16.II.2011, leg. J.-C. Chen; • 2♀♀ (KMNH), Yakou (啞口), 1.VIII.1986, leg. K. Baba, identified as B. acutangula by Kimoto (1989); • Nantou: 10♂♂, 15♀♀ (TARI), Hsitou (溪頭), 25.IV.2025, leg. C.-F. Lee; • 1♂ (TARI), Tatachia (塔塔加), 29.X.2009, leg. C.-F. Lee; • 1♂ (TARI), same but with “9.V.2011”; • 5♂♂, 8♀♀ (TARI), same but with “23.IV.2025”; • Pingtung: 2♂♂, 5♀♀ (TARI), Peitawushan (北大武山), 13.III.2025, leg. J.-C. Chen; • 2♂♂, 3♀♀ (TARI), same but with “20.III.2025”; • Taitung: 2♂♂, 3♀♀ (MHNUK), 14 km W of Chihshang (池上), 23°09'N, 121°04'E, 900 m, 16.XI.2008, leg. L. Dembický; • 1♂, 6♀♀ (TARI), Hsiangyang (向陽), 1.VII.2009, leg. M.-H. Tsou; • 3♂♂, 4♀♀ (TARI), Motien (摩天), 23.V.2011, leg. C.-F. Lee; • 1♂, 1♀ (TARI), Liyuan (栗園), 24.I.2014, leg. W.-C. Huang; • 2♂♂, 1♀ (TARI), same but with “14.III.2014”; • 1♀ (NHMUK), Yakou Country Inn (啞口山莊), 23°16.063'N, 120°58.419'E, 2582 m, 11.VIII.2008, leg. H. Mendel & M. V. L. Barclay; • Yunlin: 3♂♂, 1♀ (NMNS), Shihpi (石壁), 20.II.1991, leg. C. C. Chiang; • 1♂, 1♂ (NMNS), same but with “22.II.1991”; • 2♂♂, 1♀ (NMNS), same locality, 27.X.1992, leg. W. T. Yang.
Diagnosis.
Adults of B. tsoui sp. nov., B. chungi sp. nov., and B. choui sp. nov. are recognized by their strongly and apically narrowed elytra, and divergent elytral apices, but B. tsoui sp. nov. and B. chungi sp. nov. differ in possessing flattened elytra in males (Fig. 21C) but convex elytra in females (Fig. 21F) [convex elytra and elytral apex not visible in dorsal views in both sexes of B. choui sp. nov. (Fig. 5C, F)], and widened apex of aedeagus (Fig. 22C) [parallel-sided aedeagus in B. choui sp. nov. (Fig. 6C)]. Adults of B. tsoui sp. nov. are not separable from those B. chungi sp. nov. by external morphology but the aedeagus of B. tsoui sp. nov. (Fig. 22C) is narrower than that of B. chungi sp. nov. (Fig. 8C).
Habitus of Batophila tsoui sp. nov. A. Male, paratype, dorsal view; B. Ditto, ventral view; C. Ditto, lateral view; D. Female, paratype, dorsal view; E. Ditto, ventral view; F. Ditto, lateral view.
Batophila tsoui sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Description.
Male. Length 1.86–1.94 mm, width 0.77–0.80 mm. General color metallic dark bronze (Fig. 21A–C); legs yellowish but femora of hind legs darkened. Antenna (Fig. 22A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.7: 0.5: 0.6: 0.8: 0.7: 0.8: 0.7: 0.7: 0.7: 0.9; ratio of length to width of antennomeres I–XI 2.8: 2.5: 2.4: 3.0: 3.3: 2.6: 2.4: 2.3: 2.0: 2.1: 2.5. Pronotum 1.17–1.22× wider than long; lateral margins slightly rounded, anterolateral angles separated from lateral margins by weak emarginations, slightly narrowed basally, distance between anterolateral angles 1.14–1.22× wider than basal margin. Elytra 1.48–1.49× longer than wide; lateral margins rounded, widest at basal 1/5, apically and strongly narrowed, apex truncate but divergent; dorsoventrally flattened, apex visible in dorsal view; disc with longitudinal lines of extremely coarse punctures and with distinct longitudinal grooves along punctures, punctures and grooves apically abbreviated from apical 1/3. Tarsomeres I of front and middle legs slightly swollen. Aedeagus (Fig. 22C, D) elongate, 5.8× longer than wide; widest at apical 1/5, apically narrowed towards apex, apex widely rounded, basally widened near apical 2/5, then widened near base; dorsal opening starting from apical 1/9–1/3, tectum composed of three lobes, median lobe more ventral relative to lateral lobes, apical margin truncate, mostly membranous; slightly curved in lateral view, apex moderately curved; ventral surface with membranous area wider than dorsal opening, starting from apical 1/10–3/5.
Female (Fig. 21D–F). Length 1.99–2.56 mm, width 0.88–1.05 mm. Antennae similar to males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 22B) 1.0: 0.6: 0.6: 0.6: 0.8: 0.7: 0.8: 0.7: 0.7: 0.6: 1.0; ratio of length to width of antennomeres I–XI 2.6: 2.3: 2.7: 3.0: 3.5: 2.6: 2.5: 2.2: 2.0: 1.9: 2.8. Elytra 1.46–1.67× longer than wide; lateral margins rounded, widest at basal 1/5, apex truncate but divergent; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of extremely coarse punctures and with distinct longitudinal grooves along punctures, punctures and grooves apically abbreviated from apical 1/3. Gonocoxae (Fig. 22F) slender, connected at basal 1/5; each gonocoxa with seven long setae and one tiny seta from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 22E) weakly sclerotized apically, with several short setae at apical area, and some tiny setae at apical margin, spiculum extremely elongate. Spermathecal receptaculum (Fig. 22G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Food plants.
Melastomataceae: Otanthera scaberrima (Hayata) Ohwi (Fig. 9E, F); Rosaceae: Rubus formosensis Kuntze, R. morii Hayata, R. croceacanthus H. Lév., R. wallichianus Wight & Arn. (Fig. 9D).
Etymology.
This new species is named for Mei-Hua Tsou (曹美華), the first member of TCRT to collect specimens.
Distribution.
This species is widespread in mountainous areas in southern Taiwan (Fig. 7).
Batophila
wusheensis
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
713818CA-2B57-5CAF-850E-8074F52C3B89
https://zoobank.org/D26AAC6F-F64D-4394-8C84-85585A9F2AC6
Batophila yangweii : Chûjô 1937: 54 (part).
Type specimens examined (n = 201).
Holotype ♂ (TARI): Taiwan. Nantou: Nantou: Wushe (霧社), 30.VIII.–2.IX.1982, leg. L. Y. Chou & K. C. Chou. Paratypes. 88♂♂, 55♀♀ (TARI), same data as holotype; Nantou: 5♂♂, 5♀♀ (NHMUK), (Chingying, 精英), 24°02.530'N, 121°12.555'N, 1920 m, 6.VIII.2008, leg. M. V. L. Barclay, H. Mendel & R. Ewers; 3♀♀ (NMNS), Chunyang (春陽), 9.IV.–7.V.2002, leg. C. S. Lin & W. T. Yang, Malaise trap; 1♂, 1♀ (TARI), Hoshe (和社), 22.VII.1982, leg. L. Y. Chou & T. Lin; 2♀♀ (TARI), Musha (= Wushe, 霧社), 18.V.–15.VI.1919, leg. T. Okuni, both identified as B. yangweii by Chûjô (1937); 1♂ (TARI), same locality, 23–28.VI.1981, leg. K. S. Lin & W. S. Tang; 1♂ (TARI), same locality, 26–28.VIII.1981, leg. L. Y. Chou & S. C. Lin; 1♀ (TARI), same locality, 14.VII.1982, leg. S. C. Lin & C. N. Lin; 9♂♂, 7♀♀ (TARI), same locality, 7–8.X.1982, leg. K. C. Chou; 14♂♂, 13♀♀ (TARI), same locality, 19–22.IV.1983, leg. K. C. Chou & S. P. Huang; 2♀♀ (NMNS), same locality, 29–31.V.1996, leg. C. S. Lin; 1♂, 1♀ (TARI), Yu-shih (幼獅), 4.VIII.1981, leg. T. Lin & W. S. Tang.
Diagnosis.
Adults of B. wusheensis sp. nov. are not separable from those of B. houjayi sp. nov., B. yuae sp. nov., B. jungchani sp. nov., and B. huangi sp. nov. that are characterized by truncate elytral apices based on external morphology (Figs 10, 12) except for the aedeagus (see below). However, these species can be recognized by their allopatric distributions [B. wusheensis sp. nov. inhabits at lowlands in Nantou County, B. houjayi sp. nov. in high mountains in Chiayi, Ilan, Hualien, and Nantou counties, B. yuae sp. nov. in lowlands in Taipei and New Taipei Cities, and Ilan County, B. jungchani sp. nov. in high mountains in Taichung and Miaoli counties, B. huangi sp. nov. in lowlands in Miaoli County and high mountains in Hsinchu and Taoyuan counties (Fig. 13)]. Aedeagal shapes are diagnostic [apically tapering aedeagus from apical 1/5 in B. wusheensis sp. nov. (Fig. 23), widely rounded apex of aedeagus in B. houjayi sp. nov. (Fig. 11C), rounded apex of aedeagus with truncate process at middle of apical margin in B. yuae sp. nov. (Fig. 26C), subapically tapering apex of aedeagus in B. jungchani sp. nov. (Fig. 15C), and rounded apex of aedeagus with small, rounded process at middle of apical margin in B. huangi sp. nov. (Fig. 14C)].
Batophila wusheensis sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Description.
Male. Length 1.57–1.74 mm, width 0.70–0.80 mm. General color metallic dark bronze; antennae yellowish brown but six apical antennomeres darker; legs yellowish but femora of hind legs darkened. Antenna (Fig. 23A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.6: 0.6: 0.6: 0.8: 0.7: 0.8: 0.8: 0.8: 0.7: 1.0; ratio of length to width of antennomeres I–XI 2.6: 2.3: 3.1: 2.7: 3.5: 2.7: 2.5: 2.6: 2.1: 2.0: 2.7. Pronotum 1.22–1.24× wider than long; lateral margins slightly rounded, disc with lateral depressions at basal 1/3, anterolateral angles separated from lateral margins by weak emarginations, slightly narrowed basally, distance between anterolateral angles 1.09–1.17× wider than basal margin. Elytra 1.40–1.46× longer than wide; lateral margins rounded, widest at basal 1/3, apex truncate; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of coarse punctures and with indistinct longitudinal grooves along lines, reduced in some individuals. Tarsomeres I of front and middle legs slightly swollen. Aedeagus (Fig. 23C, D) elongate, 5.8× longer than wide; parallel-sided, apically narrowed from apical 1/5, apex pointed; dorsal opening starting from apical 1/10 and basally membranous, tectum composed of three lobes, median lobe more ventral relative to lateral lobes and apical margin truncate, mostly membranous; moderately curved in lateral view; ventral surface with membranous area narrower than dorsal opening, starting from apical 1/20–1/2.
Female. Length 1.96–2.30 mm, width 0.87–1.00 mm. Antennae similar to males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 23B) 1.0: 0.6: 0.6: 0.6: 0.7: 0.6: 0.7: 0.7: 0.7: 0.6: 0.9; ratio of length to width of antennomeres I–XI 2.9: 2.2: 2.7: 2.7: 3.5: 2.8: 2.9: 2.4: 2.2: 2.1: 2.8. Elytra 1.42–1.51× longer than wide; lateral margins rounded, widest at basal 1/3, apex truncate; dorsoventrally convex elytral apex not visible in dorsal view; disc with longitudinal lines of coarse punctures and with indistinct longitudinal grooves along lines, reduced in some individuals. Gonocoxae (Fig. 23F) slender, connected at basal 1/5; each gonocoxa with seven long setae and one tiny seta from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 23E) weakly sclerotized apically, with several short setae at apical area, and some tiny setae at sides of apical margin, spiculum extremely elongate. Spermathecal receptaculum (Fig. 23G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Food plants.
Unknown.
Etymology.
This new species is named after its type locality, Wushe (霧社).
Distribution.
Only known from the abovementioned localities in central Taiwan (Fig. 13).
Batophila
yehi
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
07D72C33-5272-51A2-8DA9-AF1ACA2AD637
https://zoobank.org/690CABBE-A769-46DD-B9D5-D8F034D57B7B
Type specimens examined (n = 99).
Holotype ♂ (TARI). Taiwan • Taichung: Hsuehshan (雪山), 18.VI.2010, leg. W.-B. Yeh. Paratypes • 18♂♂, 12♀♀ (TARI), same data as holotype; • 15♂♂, 6♀♀ (TARI), same but with “3.V.2007”; • 1♀ (TARI), same but with “14.VI.2008”; • 5♂♂, 1♀ (TARI), same but with “4.VIII.2010”; • 1♀ (TARI), same but with “8.IV.2011”; • 12♂♂, 8♀♀ (TARI), same but with “10.VI.2011”; • 1♂ (TARI), same but with “3.VIII.1911”; • 1♀ (TARI), same locality, 29.IV.-28.VI.2012, leg. L.-P. Hsu; • Miaoli: 1♀ (TARI), Hsuehchien (雪見), 23.III.2014, leg. W.-B. Yeh; • Nantou: 6♂♂, 10♀♀ (TARI), Hehuanshan (合歡山), 23.VI.2018, leg. H.-F. Lu.
Diagnosis.
Adults of B. yehi sp. nov. are similar to those of B. taiwanica Döberl and B. meihuai sp. nov. in possessing convergent elytral apices. However, adults of B. yehi sp. nov. are recognized by their stout antennae, length of antennomeres VI–X 0.5× length of antennomere I (Fig. 25A, B) [> 0.5× in B. taiwanica (Fig. 20A, B) and B. meihuai sp. nov. (Fig. 18A, B)]. Adults of B. meihuai sp. nov. are characterized by their distinct and sexually dimorphic longitudinal ridges on the elytra (Fig. 17) [indistinct or reduced longitudinal ridges on the elytra in B. taiwanica (Fig. 19) and B. yehi sp. nov. (Fig. 24)]. The aedeagi of these species are diagnostic: truncate apex in B. meihuai sp. nov. (Fig. 18C), rounded apex with narrowly rounded process at middle of apical margin in B. taiwanica (Fig. 20C), and widely rounded apex in B. yehi sp. nov. (Fig. 25C).
Habitus of Batophila yehi sp. nov. A. Male, paratype, from Hsuehshan (雪山), dorsal view; B. Ditto, ventral view; C. Ditto, lateral view; D. Female, paratype, from Hehuanshan (合歡山), dorsal view; E. Ditto, ventral view; F. Ditto, lateral view.
Batophila yehi sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Description.
Male. Length 1.39–1.57 mm, width 0.69–0.78 mm. General color metallic dark bronze (Fig. 24A–C); antennae yellowish brown but six apical antennomeres darker; legs yellowish but femora of hind legs darkened. Antenna (Fig. 25A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.5: 0.4: 0.5: 0.5: 0.5: 0.5: 0.5: 0.5: 0.5: 0.7; ratio of length to width of antennomeres I–XI 2.9: 2.0: 1.8: 2.0: 2.3: 2.2: 2.0: 1.9: 1.5: 1.6: 2.3. Pronotum 1.22–1.25× wider than long; lateral margins slightly rounded, anterolateral angles separated from lateral margins by weak emarginations, widest at middle, slightly narrowed basally, distance between anterolateral angles 0.99–1.01× wider than basal margin. Elytra 1.28–1.33× longer than wide; lateral margins rounded, widest at basal 1/3, apex widely rounded and converge; dorsoventrally flattened, apex visible in dorsal view; disc with longitudinal lines of coarse punctures but lacking longitudinal grooves and ridges. Tarsomeres I of front and middle legs strongly swollen. Aedeagus (Fig. 25C, D) elongate, 5.3× longer than wide; lateral margins basally and slightly widened towards base, apex widely rounded; dorsal opening starting from apical 1/10 and basally membranous, tectum composed of three lobes, median lobe more ventral relative to lateral lobes and apical margin truncate, mostly membranous; moderately curved in lateral view; ventral surface with membranous area narrower than dorsal opening, starting from apical 1/20–2/5.
Female. Length 1.63–1.73 mm, width 0.81–0.89 mm. Antennae similar to males, but antennomeres VIII–X wider than those of males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 25B) 1.0: 0.6: 0.4: 0.4: 0.5: 0.5: 0.5: 0.5: 0.5: 0.5: 0.8; ratio of length to width of antennomeres I–XI 2.4: 2.0: 1.9: 1.7: 2.0: 1.8: 1.5: 1.3: 1.4: 1.4: 2.1. Elytra 1.28–1.32× longer than wide; lateral margins rounded, widest at basal 1/3, apex convergent; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of coarse punctures but lacking longitudinal grooves and ridges. Gonocoxae (Fig. 25F) slender, connected at basal 1/5; each gonocoxa with seven long setae and one tiny setae from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 25E) weakly sclerotized apically, with several short setae at sides of apex, and some tiny setae at sides of apical margin, spiculum extremely elongate. Spermathecal receptaculum (Fig. 25G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Variation.
Individuals collected from Hehuanshan (合歡山) (Fig. 24D–F) have longitudinal grooves connected with lines of coarse punctures on the elytra.
Food plants.
Rosaceae: Fragaria hayatai Makino.
Etymology.
This new species is named for Dr. Wen-Bin Yeh (葉文斌), who worked as professor at the National Chung Hsing University and collected most of the type series.
Distribution.
This species is found in alpine areas of central Taiwan (Fig. 13).
Batophila
yuae
sp. nov.
Taxon classificationAnimaliaColeopteraChrysomelidae
5DFD2583-0143-544C-AF10-EAD69149DE9C
https://zoobank.org/F0E8D7D7-5893-4A66-B1C8-7CDA46803066
Batophila yangweii : Chûjô 1937: 54 (part).
Type specimens examined (n = 20).
Holotype ♂ (TARI). Taiwan • Taipei: Lengshuikeng (冷水坑), 29.VI.2008, leg. M.-H. Tsao (sic!). Paratypes • 2♀ (TARI), same data as holotype; • 1♀ (TARI), same locality, 22.II.2015, leg. M.-H. Tsou; • Ilan: 1♀ (NMNS), Chilan (棲蘭), 15.V.1999, leg. S. Halbert, C. W. & L. B. O’Brien; • 1♂ (TARI), Fushan Botanical Park (福山植物園), 19–26.VI.2006, leg. C.-S. Tung; • 1♀ (TARI), same but with “14–21.VII.2007”; • 2♂♂, 2♀♀ (TARI), Mingchi (明池), 5.IV.2009, leg. M.-H. Tsou; • 1♀ (TARI), Nishimura (= Hsitsun, 西村), 24.VII.1919, leg. Y. Miwa, identified as B. yangweii by Chûjô (1937); • 2♂ (NMNS), Wufangchi (sic!) (五峰旗), 16.X.1990, leg. C. C. Chiang; • New Taipei City: 1♂, 1♀ (TARI), Hinokiyama (= Kueishan, 檜山), 22.VII.1929, leg. Y. Miwa, both identified as B. yangweii by Chûjô (1937); • 1♀ (TARI), Wulai (烏來), 26.X.2006, leg. S.-F. Yu; • 1♀ (TARI), Yingtzuling (鶯子嶺), 9.V.2010, leg. M.-H. Tsou; • Taipei City: 1♀ (TARI), Sôzan (= Yangmingshan, 陽明山), 28.IV.1940, leg. S. Miyamoto; • 1♀ (TARI), Tatunshan (大屯山), 26.V.2010, leg. S.-F. Yu.
Diagnosis.
Adults of B. yuae sp. nov. are not separable from those of B. houjayi sp. nov., B. wusheensis sp. nov., B. jungchani sp. nov., and B. huangi sp. nov. that are characterized by truncate elytral apices based on external morphology (Figs 10, 12) except for the aedeagus (see below). However, these species can be recognized by their allopatric distributions [B. yuae sp. nov. inhabits lowlands in Taipei and New Taipei Cities, and Ilan County, B. wusheensis sp. nov. in lowlands of Nantou County, B. houjayi sp. nov. in high mountains of Chiayi, Ilan, Hualien, and Nantou counties, B. jungchani sp. nov. in high mountains of Taichung and Miaoli counties, B. huangi sp. nov. in lowlands of Miaoli County and high mountains of Hsinchu and Taoyuan counties (Fig. 13)]. Aedeagal shapes are diagnostic [rounded apex of aedeagus with truncate process at middle of apical margin in B. yuae sp. nov. (Fig. 26C), apically tapering aedeagus from apical 1/5 in B. wusheensis sp. nov. (Fig. 23C), widely rounded apex of aedeagus in B. houjayi sp. nov. (Fig. 11C), subapically tapering apex of aedeagus in B. jungchani sp. nov. (Fig. 15C), and rounded apex of aedeagus with a small, rounded process at middle of apical margin in B. huangi sp. nov. (Fig. 14C)].
Batophila yuae sp. nov. A. Antenna, male; B. Antenna, female; C. Aedeagus, dorsal view; D. Aedeagus, lateral view; E. Abdominal ventrite VIII, female; F. Spermatheca; G. Gonocoxae.
Description.
Male. Length 1.53–1.67 mm, width 0.76–0.80 mm. General color metallic dark bronze; antennae yellowish brown but six apical antennomeres darker; legs yellowish but femora of hind legs darkened. Antenna (Fig. 26A) filiform and antennomeres VIII–X wide, ratio of length of antennomeres I–XI to length of antennomere I 1.0: 0.6: 0.5: 0.5: 0.6: 0.6: 0.7: 0.7: 0.7: 0.6: 0.8; ratio of length to width of antennomeres I–XI 2.6: 2.1: 2.2: 2.4: 2.6: 2.5: 2.3: 2.0: 2.1: 1.8: 2.4. Pronotum 1.24–1.25× wider than long; lateral margins slightly rounded, anterolateral angles separated from lateral margins by weak emarginations, slightly narrowed basally, distance between anterolateral angles widest, 1.10–1.15× wider than basal margin. Elytra 1.31–1.34× longer than wide; lateral margins rounded, widest at basal 1/5, apex truncate; dorsoventrally flattened, apex visible in dorsal view; disc with longitudinal lines of coarse punctures, and indistinct longitudinal grooves along punctures present only near base, no ridges present between longitudinal grooves. Tarsomeres I of front and middle legs slightly swollen. Aedeagus (Fig. 26C, D) elongate, 6.6× longer than wide; parallel-sided, apically and strongly tapering near apex, apex with one transverse process, apical margin truncate; dorsal opening starting from apical 1/7 and basally membranous, tectum composed of three lobes, median lobe more ventral relative to lateral lobes and apical margin truncate, mostly membranous; moderately curved in lateral view; ventral surface with membranous area slender and small, starting from apical 1/8–1/3.
Female. Length 1.74–1.78 mm, width 0.86–0.92 mm. Antennae similar to males, ratio of length of antennomeres I–XI to length of antennomere I (Fig. 26B) 1.0: 0.6: 0.5: 0.6: 0.7: 0.6: 0.7: 0.6: 0.6: 0.6: 0.8; ratio of length to width of antennomeres I–XI 2.8: 2.5: 2.9: 2.9: 3.4: 2.5: 2.5: 2.3: 2.2: 2.0: 2.5. Elytra 1.31–1.38× longer than wide; lateral margins rounded, widest at basal 1/3, apex truncate; dorsoventrally convex, apex not visible in dorsal view; disc with longitudinal lines of coarse punctures, and indistinct longitudinal grooves along punctures present only near base, no ridges present between longitudinal grooves. Gonocoxae (Fig. 26F) slender, connected from basal 1/5 to base; each gonocoxa with seven long and one tiny setae from apical 1/5 to apex, subapically slightly curved. Ventrite VIII (Fig. 26E) weakly sclerotized apically, with several short setae at sides of apex, spiculum extremely elongate. Spermathecal receptaculum (Fig. 26G) strongly swollen, with transverse wrinkles at basal 1/2; pump wide and curved, with transverse wrinkles at apical 2/3; sclerotized spermathecal canal moderately long before base of spermathecal gland.
Food plants.
Rosaceae: Rubus corchorifolius L. f.
Etymology.
This new species is named for Su-Fang Yu (余素芳), the first member of TCRT to collect specimens.
Distribution.
This species is widespread at lowlands of northern Taiwan (Fig. 13).
Key to Taiwanese species of Batophila
**: **
Discussion
Species diversity of Batophila in Taiwan is unexpectedly high by comparison with ten species in mainland China (Bezděk and Konstantinov 2024), probably because taxonomic studies have been hampered by the small body sizes. Specimens smaller than 2.0 mm can be damaged during aedeagal dissections. Moreover, diagnostic characters were difficult to observe historically due to the absence of modern optics and insufficient numbers of specimens. Species are adapted to various microhabitats ranging from alpine areas to lowlands, and adults are wingless. Although high species numbers occur in several genera, including Paraplotes Laboissière (Lee 2015) Agetocera Hope (Lee et al. 2010), Sikkimia Duvivier (Lee and Bezděk 2016), Siemssenius Weise (Lee 2016), Shairella Chûjô (Lee and Beenen 2017), Lochmaea Weise (Lee 2018), and Furusawaia Chûjô (Lee and Bezděk 2021), only Paraplotes have similar high species diversity (> 10 species) as Batophila. In addition to allopatric speciation, two species in the same areas also occur in both genera.
At many localities two species of Batophila occur sympatrically. Finding more than two species of Batophila at some localities was unexpected. These included Alishan (阿里山), Sungkang (松崗), and Tsuifeng (翠峰). Based on recent field collections, specimens collected from a single host plant belonged to one Batophila-species. Further investigations are needed to clarify host specificity in different localities.
In this study the aedeagus is found to have more diagnostic value than previously recognized, especially in the position of the dorsal opening and membranous area on the ventral surface. However, diagnostic value of female genitalia is very limited in this genus by comparison with those of Argopistes Motschulsky (Lee et al. 2024). Some diagnostic external morphological characters were found, such as shape of elytra and antenna, and punctuation on the elytra. Although some species are not externally distinguishable, they can be identified by their allopatric distributions and aedeagi.
Various species of Rubus (Rosaceae) were confirmed as dominant host plants for members of Batophila in Taiwan. Species of this genus in Europe are restricted to species of Rosaceae (Jolivet and Hawkeswood 1995). Interestingly, some Taiwanese species feed on plants in other families. For example, adults of B. houjayi sp. nov. feed on leaves of Persicaria thunbergia and P. chinense (Polygonaceae), and those of B. tsoui sp. nov. feed on leaves of Otanthera scaberrima (Melastomataceae). Thus, species diversity of Batophila of the world is likely still underestimated.
Supplementary Material
XML Treatment for Batophila acutangula
XML Treatment for Batophila alishanensis
XML Treatment for Batophila choui
XML Treatment for Batophila chungi
XML Treatment for Batophila houjayi
XML Treatment for Batophila huangi
XML Treatment for Batophila jungchani
XML Treatment for Batophila meihuai
XML Treatment for Batophila taiwanica
XML Treatment for Batophila tsoui
XML Treatment for Batophila wusheensis
XML Treatment for Batophila yehi
XML Treatment for Batophila yuae
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bezděk J Konstantinov AS (2024) Supertribe Alticitae. In: Bezděk J Sekerka L (Eds) Catalogue of Palaearctic Coleoptera.Volum 6/2. Updated and Revised Second Edition. Chrysomeloidea (Orsodacnidae, Megalopodidae, Chrysomelidae). Brill, Leiden/Boston, 468–580. 10.1163/9789004707283 · doi ↗
- 2Chen S-H (1933) Study of Chinese Halticinae beetles with descriptions of some exotic new species.Sinensia 3: 211–254.
- 3ChûjôM (1937) Studies on the Chrysomelidae in the Japanese Empire (VIII). Subfamily Halticinae (8).Transactions of the Natural History Society of Formosa 27: 52–58.
- 4Döberl M (2010) Two new Alticinae from Taiwan (Coleoptera: Chrysomelidae: Alticinae).Acta Coleopterologica 26: 65–67.
- 5Duckett CN Daza JD (2004) A new species of flea beetle in the genus Alagoasa Bechyné (Coleoptera: Chrysomelidae: Galerucinae) from highland Costa Rica.Coleopterists Bulletin 58(1): 84–96. 10.1649/608 · doi ↗
- 6Heikertinger F (1921) Die paläarktischen Arten der Halticinengattung Batophila Foudr. (Glyptina Lec.). (54. Beitrag zur Kenntnis der Halticinen).Koleopterologische Rundschau 9: 89–98.
- 7Heikertinger F (1948) Bestimmungstabellen europäischer Käfer (10. Stück). LXXII. Fam. Chrysomelidae. 5. Subfam. Halticinae. 11.-30. Gattung: Crepidodera-Verwandtschaft weitesten Sinnes.Koleopterologische Rundschau 31: 15–80.
- 8Jolivet P Hawkeswood PJ (1995) Host-plants of Chrysomelidae of the world.Backhuys Publishers, Leiden, 281 pp.