The microsnail genera Clostophis and Acinolaemus (Eupulmonata, Pupilloidea, Hypselostomatidae) from central Thailand, with description of three new species

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
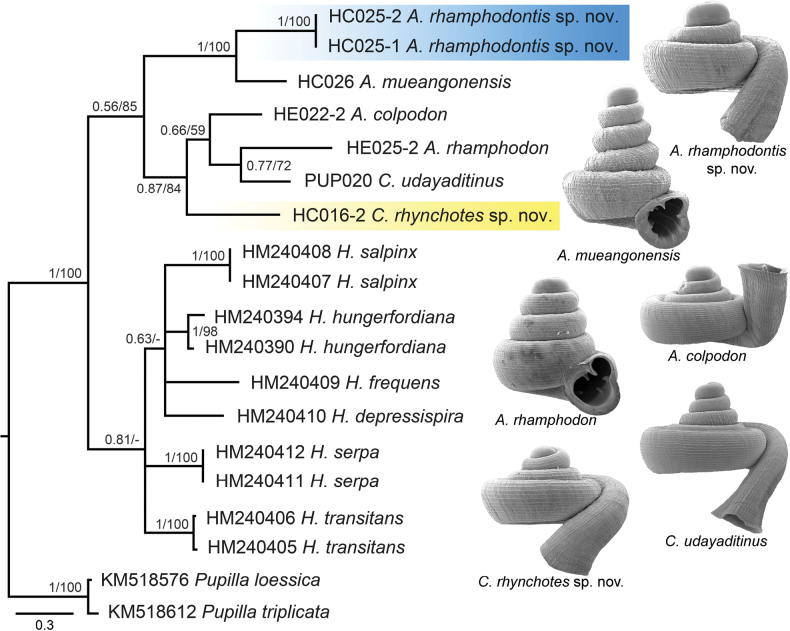 Figure 1
Figure 1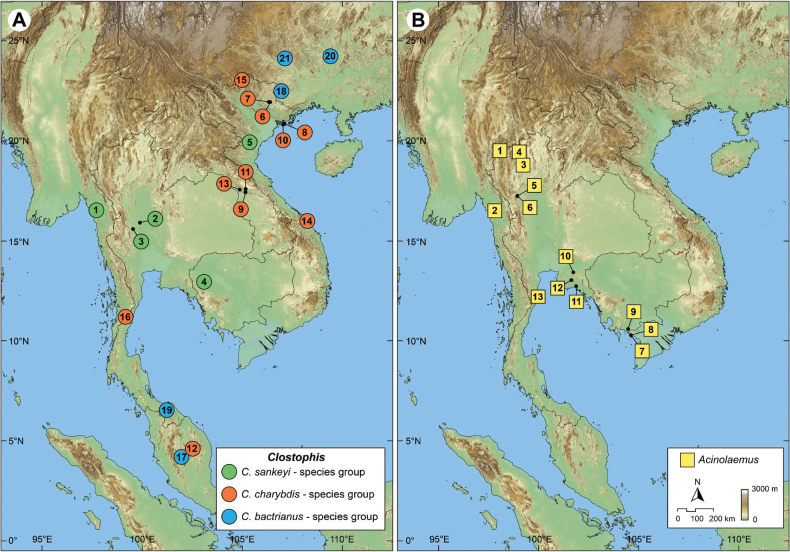 Figure 2
Figure 2 Figure 3
Figure 3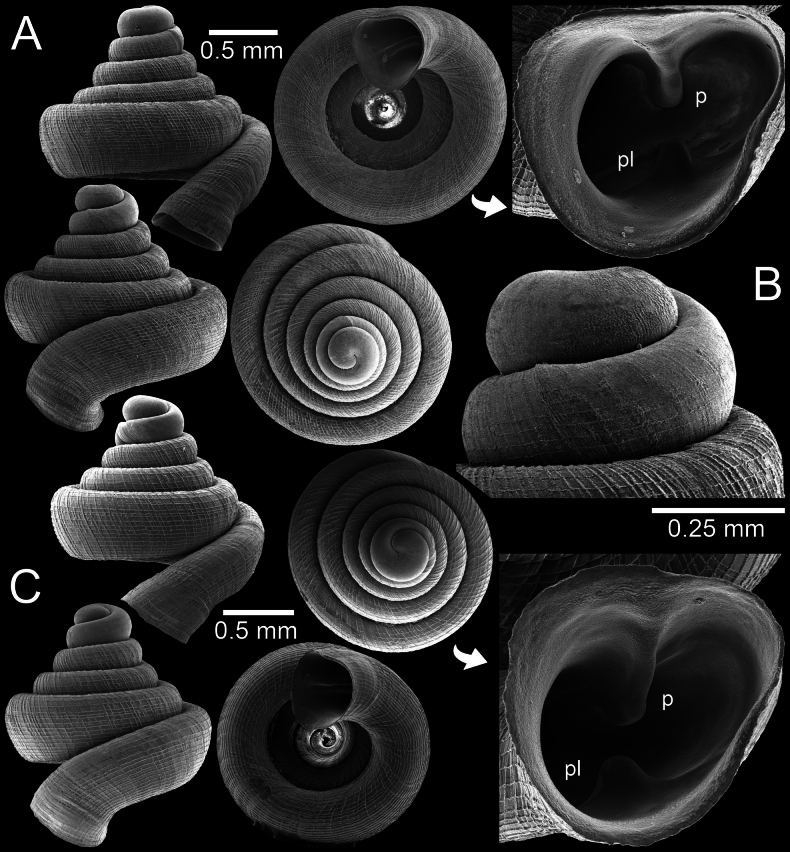 Figure 4
Figure 4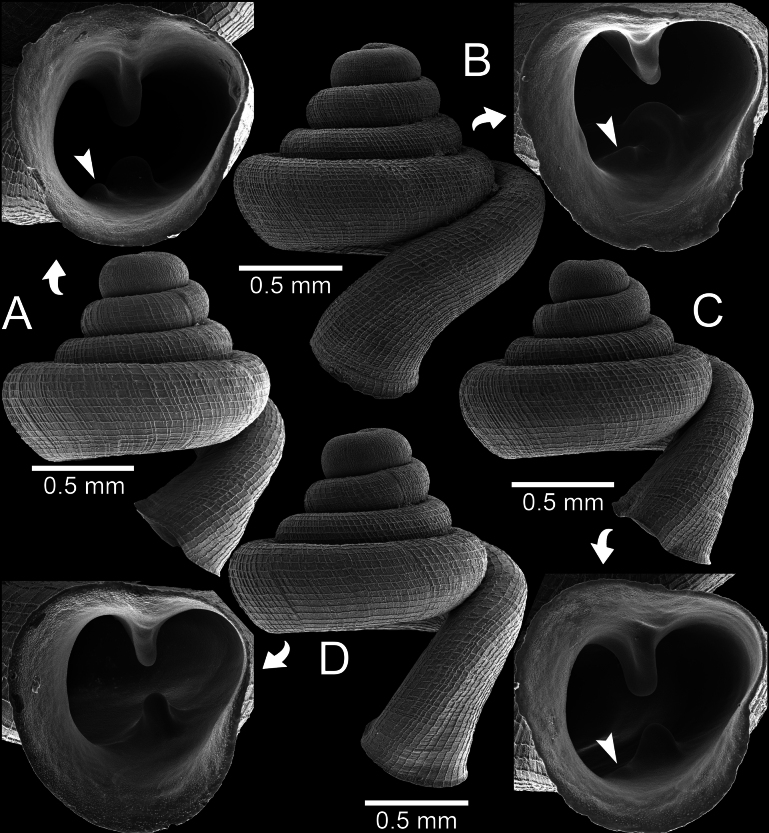 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7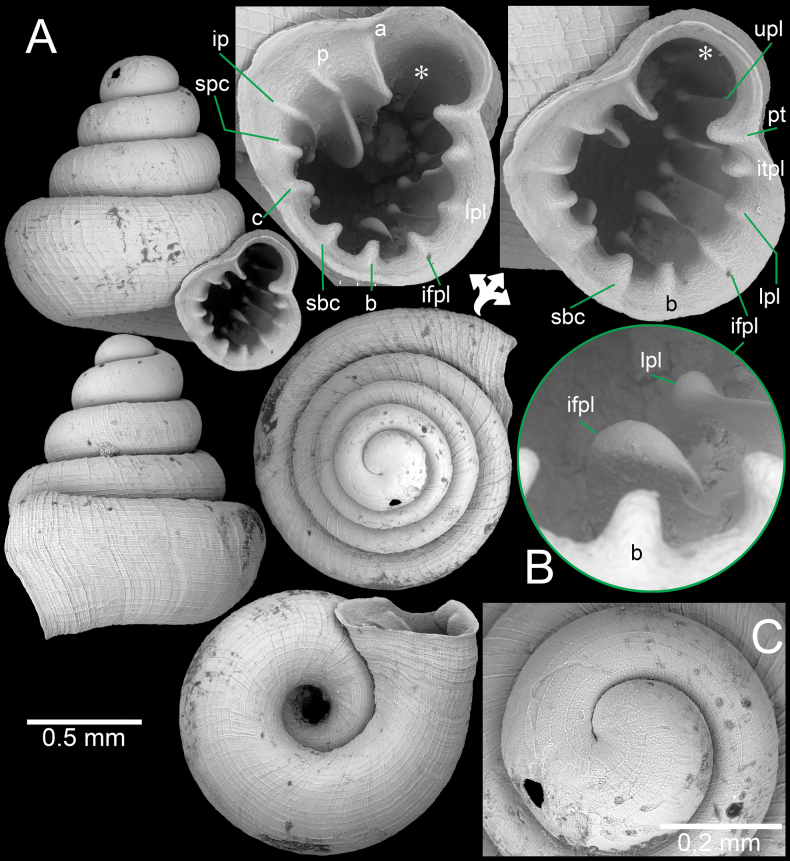 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12 Figure 13
Figure 13 Figure 14
Figure 14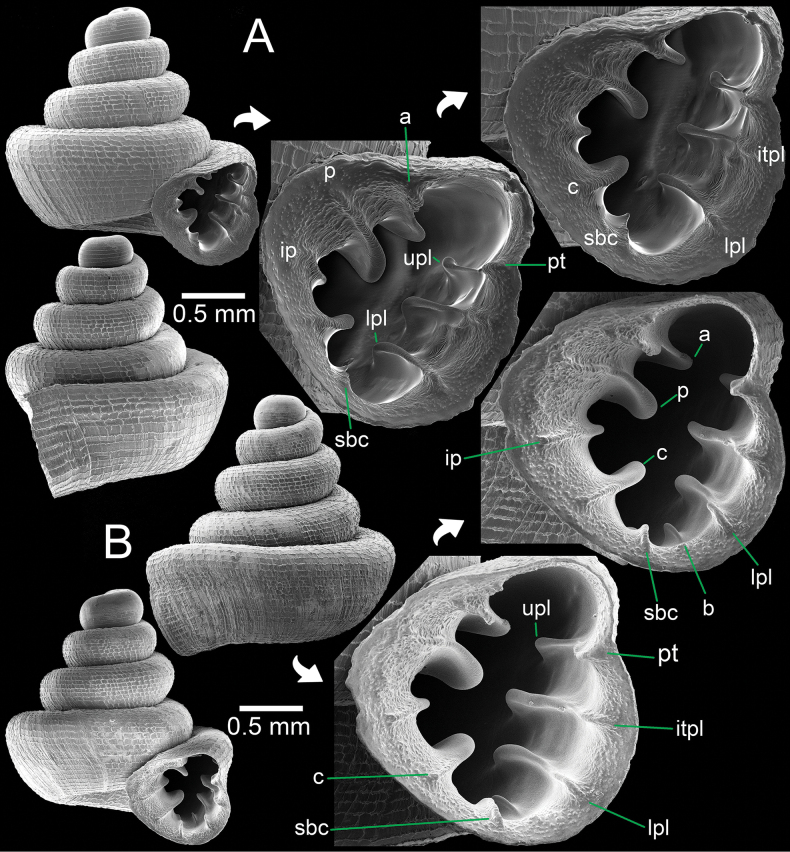 Figure 15
Figure 15 Figure 16
Figure 16 Figure 17
Figure 17| Taxon | Code | Museum registration number | GenBank accession number (COI) | Locality | Reference |
|---|---|---|---|---|---|
| HC025-1 | Paratype |
| Tak, Thailand | this study | |
| HC025-2 | Paratype |
| Tak, Thailand | this study | |
|
| HC026 |
| Tak, Thailand | this study | |
| HE022-2 |
| Chon Buri, Thailand | this study | ||
| HE025-2 |
| Chon Buri, Thailand | this study | ||
| HC016-2 | Paratype |
| Nakhon Sawan, Thailand | this study | |
| PUP020 | Paratype |
| Battambang, Cambodia | this study | |
| - | - |
| Kelantan, Malaysia | 1 | |
| - | - |
| Perak, Malaysia | 1 | |
| - | - |
| Pahang, Malaysia | 1 | |
| - | - |
| Pahang, Malaysia | 1 | |
| - | - |
| Kelantan, Malaysia | 1 | |
| - | - |
| Kelantan, Malaysia | 1 | |
| - | - |
| Kedah, Malaysia | 1 | |
| - | - |
| Kedah, Malaysia | 1 | |
| - | - |
| Kedah, Malaysia | 1 | |
| - | - |
| Kedah, Malaysia | 1 | |
| - | - |
| Belyashi, Altai, Russia | 2 | |
| - | - |
| Hracholusky, Bohemia, Czech Republic | 2 |
| Taxa | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. | 0 | |||||||||||
| 2. | 0.184 | - | ||||||||||
| 3. | 0.167 | 0.144 | - | |||||||||
| 4. | 0.155 | 0.183 | 0.179 | - | ||||||||
| 5. | 0.190 | 0.163 | 0.134 | 0.175 | - | |||||||
| 6. | 0.179 | 0.131 | 0.131 | 0.184 | 0.156 | - | ||||||
| 7. | 0.210 | 0.187 | 0.182 | 0.196 | 0.192 | 0.197 | 0.031 | |||||
| 8. | 0.174 | 0.172 | 0.164 | 0.190 | 0.163 | 0.173 | 0.137 | 0.003 | ||||
| 9. | 0.204 | 0.192 | 0.168 | 0.209 | 0.185 | 0.197 | 0.167 | 0.147 | 0 | |||
| 10. | 0.182 | 0.176 | 0.154 | 0.179 | 0.170 | 0.181 | 0.139 | 0.137 | 0.151 | 0.067 | ||
| 11. | 0.219 | 0.218 | 0.192 | 0.212 | 0.202 | 0.214 | 0.179 | 0.166 | 0.169 | 0.147 | - | |
| 12. | 0.198 | 0.198 | 0.183 | 0.195 | 0.177 | 0.197 | 0.161 | 0.166 | 0.145 | 0.139 | 0.179 | - |
| Species | Shell shape | Tubular | Protoconch | Aperture opening | Apertural dentition | Type locality |
|---|---|---|---|---|---|---|
|
| ||||||
| 1 | concave-conical | long descending | pitted and spirally striated | ventral to subventral | 1 (parietal) or 2 (parietal, palatal) | Myanmar, Mon State |
| 2 | concave-conical | long and descending | pitted and spirally striated | ventral to subventral | 2 (parietal, palatal) | Thailand, Nakornsawan |
| 3 | concave-conical | long and descending | pitted and spirally striated | ventral to subventral | 1 parietal | Uthai Thani, Thailand |
| 4 | concave-conical | long and descending | pitted and spirally striated | ventral to subventral | 4 (parietal, infraparietal, palatal, columellar) | Cambodia, Battambang |
| 5 | concave-conical | long and descending | spiral striations | subventral | none | Vietnam, Thanh Hoa |
|
| ||||||
| 6 | concave-conical | very short to absent | pitted and spirally striated | lateral | 2 (parietal, palatal) | Vietnam, Lang Son |
| 7 | conical, slightly concave side | short | spirally striated | sublateral | none | Vietnam, Lang Son |
| 8 | conical, straight to slightly convex side | very short to absent | n/a | lateral | 2 (parietal, palatal) | Vietnam, Quang Ninh |
| 9 | conical, straight side | absent | pitted and spirally striated | lateral | none | Vietnam, Haiphong |
| 10 | low conical, concave side | very short to absent | spirally striated | lateral | 1 parietal | Laos, Khammouan |
| 11 | conical, straight side | short to slightly long | pitted and spirally striated | lateral or sublateral | 1 parietal | Laos, Khammouane |
| 12 | conical, straight side | very short to absent | spirally striated | lateral | 2 (parietal, palatal) | Malaysia, Pahang |
| 13 | conical, straight to slightly convex side | very short to absent | pitted and spirally striated | lateral | none | Laos, Khammouane |
| 14 | conical, slightly concave side | short and descending | spirally striated | sub lateral | 2 (parietal, palatal) | Vietnam, Da Nang |
| 15 | conical, slightly convex side | very short to absent | spirally striated | lateral | 2 (parietal, palatal) | Vietnam, Ha Giang |
| 16 | conical, straight side | short | n/a | sublateral | 1 parietal | Myanmar, Tanintharyi |
|
| ||||||
| 17 | conical, straight side | short | spirally striated | lateral | 4 (parietal, palatal, basal, columellar) | Malaysia, Pahang |
| 18 | concave-conical | very short to absent | pitted and spirally striated | lateral | 4 (parietal, palatal, basal, columellar) | China, Guangxi |
| 19 | conical, slightly concave side | very short to long and descending | spirally striated | lateral or sublateral | 5 (parietal, angular, palatal, basal, columellar) | Thailand, Yala |
| 20 | conical, slightly concave side | very short to absent | pitted and spirally striated | lateral | 4 (parietal, palatal, basal, columellar) | China, Guangxi |
| 21 | concave-conical | short | pitted and spirally striated | lateral | 4 (parietal, palatal, basal, columellar) | China, Guangxi |
| Species | Shell shape | Shell sculpture | Number of dentitions on | Palatal tubercle | Type locality | ||
|---|---|---|---|---|---|---|---|
| parietal wall | palatal wall | columellar wall | |||||
| 1. | conical, slightly concave side, no tuba | rectangular reticulation | 3 (parietal, angular, infraparietal) | 4 (3 palatals, basal) | 1 columella | present | Thailand, Mae Hong Son |
| 2. | concave-conical, no tuba | rectangular reticulation | 3 (parietal, angular, infraparietal) | 4 (3 palatals, basal) | 1 columella | present | Myanmar, Mon |
| 3. | conical, straight to concave side, no tuba | rectangular reticulation | 3 (parietal, angular, infraparietal) | 4 (3 palatals, basal) | 1 or 2 columellae | present | Thailand, Chiang Mai |
| 4. | conical, slightly concave side, no tuba | rectangular reticulation | 3 (parietal, angular, infraparietal) | 5 (4 palatals, basal) | 3 (columellae) | present | Thailand, Chiang Mai |
| 5. | conical, slightly concave side, no tuba | rectangular reticulation | 3 (parietal, angular, infraparietal) | 3 or 4 palatals | 2 (columellae) | present | Thailand, Tak |
| 6. | depressed conical, concave side, long and descending tuba | rectangular reticulation | 3 (parietal, angular, infraparietal) | 3 (2 palatals, basal) | 2 (columellae) | present | Thailand, Tak |
| 7. | depressed conical, curved side, short descending tuba | prominent spiral striations | 2 (parietal, angular) | 2 (parietal, basal) | absent | absent | Vietnam, Kien Giang |
| 8. | conical, slightly concave side, short and descending tuba | prominent spiral striations | 2 (parietal, angular) | 2 palatals | 1 columella or absent | absent | Vietnam, Kien Giang |
| 9. | conical, slightly concave side, short and ascending tuba | prominent spiral striations | 2 (parietal, angular) | 2 palatals | 1 columella | absent | Cambodia, Kampot |
| 10. | concave-conical, no tuba | prominent spiral striations | 2 (parietal, angular) | 2 palatals | 1 columella | weak | Thailand, Chachoengsao |
| 11. | conical, slightly concave side, no tuba | prominent spiral striations | 2 (parietal, angular) | 2 palatals | 1 columella | present | Thailand, Chanthaburi |
| 12. | conical, slightly concave side, long and ascending tuba | prominent spiral striations | 1 angular | 1 palatal | 1 columella | weak | Thailand, Rayong |
| 13. | conical, straight side, very short and ascending tuba | prominent spiral striations | 2 (angulo-parietal, infraparietal) | 2 palatals | 1 columella | absent | Thailand, Pra Chuap Khiri Khan |
- —Faculty of Science, Chulalongkorn University 501100015239 http://doi.org/10.13039/501100015239
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMollusks and Parasites Studies · Aquatic Invertebrate Ecology and Behavior · Freshwater macroinvertebrate diversity and ecology
Introduction
The terrestrial microsnails of Thailand belonging to the family Hypselostomatidae Zilch, 1959 (usually smaller than 5 mm) have been studied for more than 20 years (Panha 1998a, b; Panha and Burch 2005). Currently, this snail group comprises 13 nominal genera, and the species richness in each genus varies from nearly 100 species in Hypselostoma Benson, 1856 to one species in Boysia Pfeiffer, 1849 and two species in Pseudostreptaxis von Möllendorff, 1890 (Schileyko 1998; Páll-Gergely et al. 2019; Jirapatrasilp et al. 2024; Gojšina et al. 2025; MolluscaBase 2025). In the present paper, we focus on the genera Clostophis Benson, 1860 and Acinolaemus Thompson & Upatham, 1997, which are considered to be closely related and comprise morphologically similar species characterised by small (usually < 2 mm), colourless shells, with or without a detached last whorl (or tuba), and relatively variable apertural dentition (Thompson and Upatham 1997; Páll-Gergely et al. 2020; Páll-Gergely and Hunyadi 2022).
The genus Acinolaemus was originally created for eight species and was diagnosed by its colourless shell, its enlarged angular lamella forming a distinctly separated sinulus, and its spiral striation crossed by oblique radial growth lines on the protoconch (Thompson and Upatham 1997; Schileyko 1998). It is believed to be endemic to limestone hills from northern to peninsular Thailand as well as adjactent regions of Myanmar and Malaysia. Subsequently, several species have been added to this genus, so that its distribution range was expanded to include the limestone hills in the Salween Basin in Myanmar, and the Mekong Delta Karsts in Cambodia and Vietnam (Vermeulen et al. 2007, 2019; Changlom et al. 2019; Tongkerd et al. 2024). So far, seven of the 11 currently recognised species of Acinolaemus have been recorded in Thailand (Thompson and Upatham 1997; Changlom et al. 2019; MolluscaBase 2025).
The genus Clostophis was originally described as a monotypic genus (Benson 1860) that, for more than 150 years, was assigned to the caenogastropod family Diplommatinidae Pfeiffer, 1856, but since the work of Páll-Gergely et al. (2020) and Páll-Gergely and Hunyadi (2022) is now firmly classified as a speciose genus in the heterobranch family Hypselostomatidae. Currently, the genus comprises 20 nominal species which are mainly distributed in the Indo-Burma region, Peninsular Malaysia, and southern China, while only two species have been recorded in Thailand (Páll-Gergely et al. 2020; Páll-Gergely and Hunyadi 2022). Clostophis proboscideus (Panha & Burch, 2002) has a precise type locality in central Thailand (Panha and Burch 2002, 2005) while C. laidlawi (Collinge, 1902) is only known from the type locality as ‘Jalor’ (= Yala Province, Thailand), although several populations were reported from Malaysia (Collinge 1902; Páll-Gergely and Hunyadi 2022).
Chao Phraya River Basin in central Thailand plays a crucial role in the hydrology and economy of the country. The area is characterised by floodplains, highlands, and scattered limestone hills or karsts (Prasongtham and Kanjanapayont 2014; Singtuen and Phajuy 2020). Limestone hills are unique ecosystems that support significant, highly diverse, and unique biota (Clements et al. 2006). For this study, we surveyed the central Thailand limestone karsts for Clostophis and Acinolaemus microsnails and collected numerous specimens from multiple localities. Because this material clearly differed in shell shape, shell sculpture, and apertural dentition from all other known Clostophis and Acinolaemus species, we describe one new Clostophis and two new Acinolaemus species by means of shell morphology and DNA sequence data. In addition, to contribute to a better understanding of their taxonomy, C. proboscideus, A. ptychochilus Thompson & Upatham, 1997 (type species of Acinolaemus), A. cryptidentatusChanglom et al., 2019, and A. mueangonensisChanglom et al., 2019 are re-described based on the specimens collected from their type localities.
Materials and methods
Specimen sampling
The fieldwork was focused on limestone outcrops in the Chao Phraya River Basin in central Thailand. The topography in central Thailand consists of scattered small hills or karst towers above alluvial plains. The flat plains in this region are currently impacted by paddy fields and other agricultural uses, and quarrying.
Specimens were collected from limestone walls, soil, and litter debris at the base of limestone outcrops, and in rock crevices. Shells were soaked in water with detergent, and dirt or mud was removed manually using fine paintbrushes. Dry shells were examined and imaged with a Leica M205C microscope with a fusion optics and the Leica Application Suite Image System. Additional specimens were photographed by scanning electron microscopy (SEM; JEOL, JSM-6610 LV) for more detailed views of microscopic shell structures. Shell whorls were counted to the nearest quarter whorl (Kerney and Cameron 1979). Shell measurements were taken from digital images by Cell’D Imaging Software (Olympus). Identification of specimens and terminology used to describe the apertural dentition were based on Pilsbry (1918, 1948), Páll-Gergely et al. (2020), Páll-Gergely and Hunyadi (2022), Sutcharit et al. (2023), Gojšina et al. (2025), and Sutcharit et al. (2025).
DNA extraction, PCR amplification, and sequencing
Total genomic DNA was extracted from tissue samples using the G-spin Genomic DNA Extraction Kit following the manufacturer’s protocol. Approximately 655 bp of the mitochondrial cytochrome c oxidase subunit I (COI) gene was amplified by the polymerase chain reaction (PCR) using the primer pairs LCO1490 and HCO2198 (Folmer et al. 1994).
PCR reactions were performed with an initial denaturation step at 94 °C for 3 min, 35 PCR cycles (94 °C for 45 s, 45–46 °C for 45 s, 72 °C for 1 min) and a final extension step at 72 °C for 5 min. Both strands of the amplified products were purified using the DNA-spin Plasmid DNA Purification Kit and were sequenced at Bioneer Corporation, South Korea. Chromatograms were manually corrected for misreads, if necessary, and forward and reverse strands were merged into one sequence file using MEGA v. 7.0 (Kumar et al. 2016). New sequences have been deposited in GenBank (Table 1). Additionally, twelve COI sequences were downloaded from GenBank (Table 1). Ten sequences from six Hypselostoma species were used as part of the ingroup and two sequences from two Pupilla species were used as more distantly related outgroup.
Sequence alignment and phylogenetic analysis
Sequences were aligned using MEGA v. 7.0 with the default settings (Kumar et al. 2016). COI-sequence divergence was expressed as p-distances, calculated with MEGA v. 7.0 with the ‘pairwise deletion of gaps’ option (Kumar et al. 2016). Bayesian inference (BI) and maximum likelihood (ML) analyses were used to reconstruct phylogenetic relationships through the online CIPRES Science Gateway (Miller et al. 2010). The best-fit model of nucleotide substitution was identified for each sequence partition separately by means of the corrected Bayesian information criterion (BIC) using Kakusan4 (Tanabe 2011). As suggested by the Kakusan4 program, three partitions were designated: the HKY85 model with a gamma distribution for the first codon position of the COI fragment, and the GTR model with a gamma distribution for the second and the third codon positions of the COI fragment. Bayesian inference phylogenies were generated using MrBayes on ACCESS v. 3.2.7a (Ronquist et al. 2012), with two runs for 10 million generations, sampling every 1000 generations, and the first 25% of obtained trees being discarded as burn-in. Maximum likelihood phylogenetic trees were generated using RAxML-HPC2 on ACCESS v. 8.2.12 (Stamatakis 2014) using a GTRCAT model. Bootstrap support (BS) values were used for assessing branch support in the ML tree with 1000 bootstrap replicates and shown as a percentage. Branch support in the BI tree was evaluated by posterior probabilities (PP). PP values ≥ 0.95 and BS values ≥ 70% were interpreted as positive support for nodes, while lower values were interpreted as weak support. Both ML and BI phylogenetic trees were visualised and modified in FigTree v. 1.4.4 (Rambaut 2018), then visually processed in Adobe Illustrator 2021.
Institutional abbreviations
CUMZChulalongkorn University, Museum of Zoology, Bangkok;
NHMUKThe Natural History Museum, London;
SMFSenckenberg Forschungsinstitut und Naturmuseum, Frankfurt am Main;
UFFlorida Museum of Natural History, University of Florida.
Abbreviations
a, angular lamella; b, basal plica; c, columellar lamella; ifpl, infrapalatal plica; ip, infraparietal lamella; itpl, inter palatal plica; lpl, lower palatal plica; p, parietal lamella; pt, palatal tubercle; sbc, subcolumellar lamella; spc, supracolumellar lamella; upl, upper palatal plica.
Results
Molecular phylogenetic analyses
Our final dataset contained COI sequences from 17 specimens representing 12 taxa in the Hypselostomatidae. Seven species were represented by just a single specimen and five species were represented by two specimens. Additionally, two sequences from two pupillid taxa, Pupilla loessica Ložek, 1954 and P. triplicata (Studer, 1820), were included as outgroup. The final COI alignment had a total length of 655 bp, containing 264 variable sites, 391 invariant sites, 232 parsimony-informative sites, and 32 singleton sites.
The BI and ML trees were largely consistent; therefore, only the BI tree is presented in Fig. 1. Both BI and ML analyses supported the monophyly of the family Hypselostomatidae (BB/PP: 1/100). However, the phylogenetic relationships within this family remain unresolved. Although six species of Hypselostoma formed a monophyletic group in the BI analysis with weak nodal support, they were not retrieved as a monophyletic group in the ML tree. Similarly, the monophyly of the genera Clostophis and Acinolaemus was not supported. Conversely, the sister group relationship of A. rhamphodontis sp. nov. and A. mueangonensis was well-supported (BB/PP: 1/100). In contrast, the remaining Acinolaemus species (A. rhamphodon and A. colpodon) were grouped together with two Clostophis species (C. rhynchotes sp. nov. and C. udayaditinus) in a separate clade (BB/PP: 0.87/84). This phylogenetic pattern suggests that current generic boundaries between Clostophis and Acinolaemus may not accurately reflect their evolutionary relationships and highlights the need for further taxonomic revision within the group.
Bayesian inference tree from analysis of the mitochondrial cytochrome c oxidase subunit I (COI) sequence data of species in Clostophis, Acinolaemus, and Hypselostoma. Numbers on branches indicate the Bayesian posterior probabilities (PP) and ML bootstrap (BS) support values. Blue colour represents Acinolaemus rhamphodontis sp. nov. clade. Yellow colour represents Clostophis rhynchotes sp. nov. clade. Representative shells of Clostophis and Acinolaemus are shown but not to scale.
The mean interspecific p-distance among the four Acinolaemus species varied from 14.4% to 18.4% and no intraspecific sequence divergence within our examined specimens of A. rhamphodontis sp. nov. (Table 2). The p-distance between C. rhynchotes sp. nov. and C. udayaditinus was 15.6% (Table 2). The average interspecific p-distance for the six Hypselostoma species varied from 13.7% to 21.9% and the average intraspecific sequence divergence within Hypselostoma varied from 0% to 6.7% (Table 2).
Systematics
Family Hypselostomatidae Zilch, 1959
Clostophis
Taxon classificationAnimaliaStylommatophoraHypselostomatidae
Benson, 1860
9BD9F1E4-62F7-57E8-ADBF-CA71858C7F37
Clostophis Benson, 1860: 95. Kobelt 1902: 484. Thiele 1929: 111. Páll-Gergely et al. 2020: 351, 352. Páll-Gergely and Hunyadi 2022: 419. Preece et al. 2022: 145, 256. Montapiculus Panha & Burch, 2002: 148. Type species: Montapiculus proboscidea Panha & Burch, 2002. Panha and Burch 2005: 38, 109.
Type species.
Clostophis sankeyi Benson, 1860, by monotypy.
Remarks.
The genus was recently revised by Páll-Gergely et al. (2020) and Páll-Gergely and Hunyadi (2022), who recorded two species from southeastern Myanmar, six species from Laos, six species from Vietnam, two species from Peninsular Malaysia, and two species from Thailand. All these species share strong spiral ridges, usually with one or many apertural dentitions (except for four species with no dentition), and a more or less detached last whorl (tuba). Other characters, such as shell shape and tuba length, show large intraspecific variation (although they can be useful in some cases to distinguish species). Based on its wide distribution and its substantial morphological variability, the genus can be divided into three phenotypic species groups:
Approximate type localities. A. All Clostophis species. Colours indicate different species groups; numbers correspond to the species listed in Table 3; B. All Acinolaemus species; numbers correspond to the species listed in Table 4.
Clostophis sankeyi species group: with 0–4 apertural dentitions, a long (~1/4 whorl or more) and descending tuba, and a conical shell shape with strongly concave sides. This group consists of five species (including Clostophis rhynchotes sp. nov.) distributed in Myanmar, Thailand, and northern Vietnam (Fig. 2A, Table 3). Clostophis charybdis species group: with 0–2 apertural dentitions, no to short tuba, moderately to strongly conical shell, concave sides, and aperture opened laterally to sublaterally. This species group comprises ten species, of which nine species are distributed mainly in the central to northern Annamite Ranges in Laos and central to northern Vietnam; one species is found in southern Myanmar (Fig. 2A, Table 3). Clostophis bactrianus species group: with four or five strong apertural dentitions, absent to short tuba, and conical to slightly concave sides. This group consists of five species with disjunct distributions. Three species from southern China tend to have long denticles deeper inside the aperture, whereas the two species from Peninsular Malaysia tend to have short denticles situated near the apertural lip (Fig. 2A, Table 3).
Clostophis
proboscideus
Taxon classificationAnimaliaStylommatophoraHypselostomatidae
(Panha & Burch, 2002)
523EAF63-BD27-5CC3-823F-0ECAC475A586
Montapiculus proboscidea Panha & Burch, 2002 [1999]: 148, figs 4–7. Type locality: Teppratan mountain, Nakornsawan Province. Panha and Burch 2005: 109, fig. 94. Clostophis proboscideus —Páll-Gergely et al. 2020: 364, figs 1j, 3b.
Material examined.
Holotype. Thailand • Wat Khao Huai Lung (former Teppratan mountain), Ban Daen, Banphot Phisai District, Nakhon Sawan Province; 15°55'35.4"N, 99°52'23.2"E; S. Panha leg.; CUMZ 14458 (former Ver-079). Paratypes. Thailand • 2 shells (Fig. 3A–C); same data as for holotype; CUMZ 14457 (former Ver-080).
Clostophis proboscideus, paratype CUMZ 14457 (former Ver-080) from the type locality. A. Shell from different angles and showing enlarged aperture; B. Protoconch and earlier whorl sculptures; C. Shell from different angles and showing enlarged aperture. Note: the aperture of this shell was broken during the SEM imaging process.
Other material.
Thailand • 2 shells (Fig. 4A–C); Wat Khao Huai Lung (former Teppratan mountain), Ban Daen, Banphot Phisai District, Nakhon Sawan Province; 15°55'35.4"N, 99°52'23.2"E; P. Tongkerd leg.; CUMZ 14456. • 2 shells + 2 juveniles; Wat Tham Bo Ya, Nong Krot, Mueang Nakhon Sawan District, Nakhon Sawan Province; 15°43'49.7"N, 99°56'45.7"E; P. Tongkerd leg.; CUMZ 14465. • 4 shells + 1 juvenile (Fig. 5A–D); Khao Pathawee, Taluk Du, Thap Than District, Uthai Thani Province; 15°28'25.2"N, 99°45'30.3"E; P. Tongkerd leg.; CUMZ 14429. • 105 shells + 8 juveniles; same collection data as preceding; CUMZ 14463. • 4 shells; Phu Toei, Huai Khamin, Dan Chang District, Suphan Buri Province; S. Panha leg.; CUMZ 15351.
Clostophis proboscideus, topotype CUMZ 14456 from the type locality showing variation in shell shape and sculpture. A. Shell with dense spiral striae sculpture; B. Protoconch and earlier whorl sculptures; C. Shell with conical spire with strong spiral striae and radial ridges.
Clostophis proboscideus, specimen CUMZ 14429 from Khao Pathawee, Uthai Thani Province, intra population variation in basal lamella. A. Specimen with prominent infrapalatal plica; B. Specimen with low but noticeable infrapalatal plica; C. Specimen with inconspicuous infrapalatal plica; D. Specimens without infrapalatal plica. White arrowhead indicates position of infrapalatal plica.
Diagnosis.
Shell concave-conical and with long and descending tuba. Apertural dentition with one parietal lamella and one weak palatal plica. Umbilicus wide.
Description.
Shell concave-conical, colourless; growing regularly; last whorl slightly expanded. Shell height 1.7–1.9 mm (including tuba) and shell width 1.6–1.7 mm. Apex large and rounded; protoconch ~1½ whorls, pitted and with very narrow spiral striae. Whorls ~4½–5 (excluding tuba), rounded and convex; suture wide and deep. Shell surface with strong, elevated and rather equidistant spiral striae (18–22 on body whorl in frontal view), and crossed with weak to strong and irregular radial growth lines. Sometimes growth lines on earlier whorls strong, thus making reticulated surface. Penultimate whorl seemingly sunken into last whorl. Last whorl slightly shouldered to rounded; tuba long, ~¼ whorl, strongly descending, curving and slightly twisted. Aperture subovate, open ventrally to subventrally; peristome thin, slightly expanded and with weak depression on parietal side. Apertural dentitions two: parietal lamella relatively strong, high, and situated near apertural lip; palatal plica moderate, low, and situated slightly deeper inside aperture (infrapalatal plica occasionally present). Umbilicus widely opened, occupies ~½ of shell width and showing all preceding whorls.
Differential diagnosis.
Clostophis proboscideus differs from C. sankeyi and C. yoga Páll-Gergely & Hunyadi, 2022 by having more numerous and more tightly coiled whorls, lower spire, and wider umbilicus. Meanwhile, C. sankeyi has weaker spiral striations, and C. yoga possesses no apertural dentition, has a narrower umbilicus (< 1/3 of shell width), and with spiral striations throughout the protoconch (Benson 1860; Páll-Gergely and Hunyadi 2022; Preece et al. 2022).
Clostophis proboscideus differs from the long and descending tuba morphs of C. multiformis Páll-Gergely & Reischütz, 2020 and C. laidlawi by having a concave-conical shape, wider umbilicus, aperture opening ventrally to subventrally, spiral striations appearing at late stage of protoconch, and with two apertural dentitions (parietal and palatal). In comparison, these two latter species possess conical shells with straight sides, apertures opened sublaterally, narrow umbilicus, and with spiral striations throughout the protoconch. In addition, C. multiformis has only a parietal lamella, while C. laidlawi possesses five apertural dentitions (parietal, angular, upper palatal, lower palatal, and columellar) (Collinge 1902; van Benthem Jutting 1961; Páll-Gergely et al. 2020).
Distribution.
This species is currently known from several limestone outcrops in central Thailand.
Remarks.
The correct publication date of C. proboscideus has been specified in Jirapatrasilp et al. (2023: 24). This species was described based on three specimens: the holotype and two paratypes. The paratypes are photographed herein, one with a broken palatal wall and the other still intact (Fig. 3A–C). However, when we revisited the type locality, we found only two empty shells (Fig. 4A–C).
Shell variations were observed from specimens from Khao Patawi, Uthai Thani Province. These specimens possessed a general shell form similar to the type specimens but tended to have wider spaces between radial striations on the last whorl (~14–20), and the infrapalatal plica may be present. When an infrapalatal is present, it is located close to a larger plica, and the infrapalatal varies from a noticeable but low ridge (Fig. 5A) to weak and inconspicuous (Fig. 5B, C) or without an infrapalatal plica (Fig. 5D). These variations occur syntopically, and we consider this as intrapopulation variation.
Clostophis
rhynchotes
Taxon classificationAnimaliaStylommatophoraHypselostomatidae
Tongkerd & Panha sp. nov.
FC546542-DCD6-53CB-948C-EC093483CA20
https://zoobank.org/B7590776-7C97-4372-AC9E-CE1B98BCBC04
Type material.
Holotype. Thailand • height 1.3 mm (including tuba), width 1.5 mm (Fig. 6A, B); Wat Khao Chakkachan Wanaram, Chum Ta Bong District, Nakhon Sawan Province; 15°35'44.4"N, 99°32'39.3"E; P. Tongkerd leg.; CUMZ 14460. Paratypes. Thailand • 1 shell (Fig. 6C); same data as for holotype; CUMZ 14459. • 1 shell (Fig. 7B); same data as for holotype; CUMZ 14436. • 1 shell (Fig. 7A); same data as for holotype; CUMZ 14437. • 29 specimens in ethanol (Fig. 7C; COI accession no. PV698339); same data as for holotype; CUMZ 14464 (COI accession number PV698339). • 2 shells; same data as for holotype; NHMUK 20250359. • 2 shells; same data as for holotype; SMF.
Clostophis rhynchotes sp. nov. from Nakhon Sawan Province. A, B. Holotype CUMZ 14460 with enlarged aperture: B. Protoconch and earlier whorl sculptures; C. Paratype CUMZ 14459 from the type locality.
Clostophis rhynchotes sp. nov. from Nakhon Sawan Province. A. Paratype CUMZ 14437 with enlarged aperture; B. Paratype CUMZ 14436 from the type locality; C. Paratype CUMZ 14464 under light microscope showing colourless and translucent shell and with enlarged aperture.
Diagnosis.
Shell depressed concave-conical, penultimate whorl slightly sunken into last whorl, long and descending tuba, 12–14 spiral striations, peristome weakly expanded, with only parietal lamella, and wide umbilicus.
Description.
Shell depressed, concave-conical, colourless; spire growing regularly and last whorl broadly expanded. Shell height 1.3–1.4 mm (including tuba) and shell width 1.4–1.5 mm. Apex large and rounded; protoconch ~2 whorls, pitted and sculptured with prominent spiral striae. Whorls ~4–4¾ (excluding tuba) weakly shouldered and convex; suture wide and deep. Shell surface with strong, elevated, continuous, equidistant spiral striae (12–14 on body whorl in frontal view), and crossed with weaker and irregular radial growth lines. Growth lines on earlier whorls rather strong, making reticulated surface. Penultimate whorl slightly sunken into last whorl. Last whorl with slight indication of blunt shoulder; tuba short, < ¼ whorl, strongly descending and twisted. Aperture subovate, open ventrally to subventrally; peristome thin and slightly expanded on columellar side and with weak depression on parietal side. Apertural dentition one: parietal lamella strong and tall with low ridge near peristome edge then gradually taller deep inside aperture. Umbilicus widely opened, occupies ~½ of shell width and showing all preceding whorls.
Differential diagnosis.
Clostophis rhynchotes sp. nov. differs from C. proboscideus, C. sankeyi, and C. yoga by having a depressed conical spire, strong parietal lamella, and 12–14 spiral striae on last whorl. In comparison, these other three species possess conical spires with 18–20 spiral striae on last whorl. In addition, C. proboscideus has a parietal lamella and a strong palatal plica, while C. sankeyi and C. yoga generally have no dentition, but parietal lamella and palatal plica may be weakly present in C. sankeyi (Páll-Gergely et al. 2020).
Clostophis rhynchotes sp. nov. also differs from C. udayaditinus Sutcharit & Panha, 2025 from Cambodia by having slightly concave-sided shell, penultimate whorl slightly sunken into last whorl, 12–14 spiral striae on last whorl, and only one parietal lamella, while C. udayaditinus has a strongly concave shell, the penultimate whorl sunken into last whorl, 18–24 spiral striae on last whorl, and four dentitions (hooked parietal, infraparietal, palatal, and hooked columellar) (Sutcharit et al. 2025).
Distribution.
This new species is currently known only from the type locality. There is a small limestone outcrop (700 m long and 250 m wide), and a hill covered with low vegetation, surrounded by a housing area and temple, and with low disturbance.
Etymology.
The specific name rhynchotes is from the Greek word rhynchos meaning ‘snout’ and the suffix –otes; it refers to the tuba of the last whorl, which resembles the snout of a tapir.
Remarks.
Although C. rhynchotes sp. nov. shares a depressed shell, a long descending tuba, and spirally striated protoconch with A. rhamphodontis sp. nov., it is distinguished from the latter by the prominent spiral striae crossed by less prominent growth lines without forming a rectangular pattern, and with very few dentitions. These characteristics indicate the placement of this new species within the genus Clostophis.
Acinolaemus
Taxon classificationAnimaliaStylommatophoraHypselostomatidae
Thompson & Upatham, 1997
19803BBA-1E93-52D1-AC6D-8935AF7197A3
Acinolaemus Thompson & Upatham, 1997: 223, 224. Schileyko 1998: 255. Panha and Burch 2005: 39. Vermeulen et al. 2007: 86.
Type species.
Acinolaemus ptychochilus Thompson & Upatham, 1997, by original designation.
Remarks.
The genus currently contains 11 species (MolluscaBase 2025) with few to many apertural dentitions, a shell with ascending to descending tuba or without tuba, and a shell surface with rectangular reticulations to prominent spiral striations (Table 4; Thompson and Upatham 1997). These variations are possibly the cause that rendered the generic boundary ambiguous and necessitates intensive systematic revision.
Acinolaemus
ptychochilus
Taxon classificationAnimaliaStylommatophoraHypselostomatidae
Thompson & Upatham, 1997
84775BC6-930E-5846-B095-4E48015FC6FD
Acinolaemus ptychochilus Thompson & Upatham, 1997: 225, 226, figs 7–11. Type locality: Ban Prang Ma-O, Doi Pha San Sao (Mountain), Chiang Mae [= Chiang Mai] Province, Thailand. Acinolaemus ptychochilus —Schileyko 1998: 255, fig. 316.
Type material examined.
Holotype. Thailand • height 1.5 mm, width 1.3 mm (Fig. 8); Doi Pha San Sao, Ban Prang Ma-O, Chiang Mai Province; F.G. Thompson leg.; UF 113502.
Acinolaemus ptychochilus from Chiang Mai Province. A. Holotype UF 113502 with enlarged aperture from three different angles; B. Inset of hooked infrapalatal plica; C. Protoconch and earlier whorl sculptures. An asterisk () indicates small lamella and plica situated in posterior chamber. Photo credit: B. Páll-Gergely.*
Other material.
Thailand • 4 shells (Fig. 9); limestone near Chai Prakan Highway Division (~43 km to Chieng Dao District), Chai Prakan District, Chiang Mai Province; S. Panha leg.; CUMZ 15360.
Acinolaemus ptychochilus from Chiang Mai Province. A. Specimen CUMZ 15360 with enlarged aperture from three different angles; B. Protoconch and earlier whorl sculptures. An asterisk () indicates small lamella and plica situated in posterior chamber.*
Diagnosis.
Shell concave-conical, peristome weakly expanded. Apertural dentitions eleven or more, long and reaching peristome edge in form of prominent knobs: three on parietal wall, palatal tubercle, four on palatal wall, one basal, and two on columellar wall.
Re-description.
Shell conical, with concave sides, colourless; spire low to high and growing regularly; last whorl expanded. Shell height 1.3–1.4 mm and shell width 1.4–1.5 mm. Apex blunt; protoconch ~1½–2 whorls, with narrow and fine spiral striae. Whorls ~4–5, rounded and convex; suture wide, well impressed, and deep. Shell surface sculptured with equidistantly spaced spiral striae, crossed with irregular radial growth lines making a rectangular reticulated sculpture throughout shell. Penultimate whorl regularly coiled; last whorl rounded. Aperture subovate; peristome thickened, slightly expanded and with weak depression on parietal side. Aperture with approx. ten or eleven dentitions and with strong knobs reaching peristome edge. Parietal wall with three lamellae: parietal lamella large, outer part low, inner part long with tall ridge and located deeper inside aperture; infraparietal lamella long and low; angular lamella long, consisting of two peaks of prominent tall ridges which are interrupted by a low wide ridge. Palatal tubercle rectangular, situated at peristome edge and continuous with upper palatal plica. Three tiny low plicae present (not reaching peristome edge) in sinulus. Palatal wall with four plicae: upper-, inter-, lower-, and infra-palatal plicae connecting peristome edge with prominent tubercles, then continuing with narrow, low ridge, and becoming a stronger fold deep inside aperture. Folding of inter- and lower-palatal plicae larger than upper- and infra-palatal plicae. Infrapalatal plica hooked (in holotype). Basal plica a low ridge and similar to infrapalatal plica. Columellar wall with three or four lamellae: columellar lamella strong, distinct, and tall ridge; supracolumellar and one or two subcolumellar lamellae present as low ridges. Umbilicus widely perforated, ~⅓ of shell width, rounded and deep.
Distribution.
Acinolaemus ptychochilus is known from the type locality in Chiang Dao District, Chiang Mai Province in northern Thailand (Thompson and Upatham 1997).
Remarks.
The specimens from Chai Prakan District, which is ca 30 km north of the type locality, have only faint protoconch and teleoconch sculptures due to shell weathering. However, the original sculpture remains near the suture and around the umbilical area. These specimens differ slightly from the holotype (Fig. 8A) in having a slightly more depressed shell, a less expanded last whorl, infrapalatal plica that is not hooked (hooked in the type), and two subcolumellar lamellae (one in the type); however, the remaining apertural dentitions are identical to those in the holotype.
Acinolaemus
cryptidentatus
Taxon classificationAnimaliaStylommatophoraHypselostomatidae
Changlom, Chan-ard & Dumrongrojwattana, 2019
9ACF3662-3683-5345-BF24-8E1F6266AFEE
Acinolaemus cryptidentatus Changlom, Chan-ard & Dumrongrojwattana, 2019: 158, 159, fig. 2. Type locality: Tham Wua (Wua Cave), Mueang District, Mae Hong Son Province, Thailand. Tongkerd et al. 2024: 164, figs 1b, 2b.
Material examined.
Thailand • 3 shells (Fig. 10A, B); Mae La Na Cave, Pang Mapha District, Mae Hong Son Province; 19°34'30.8"N, 98°12'56.3"E; S. Panha leg.; CUMZ 15362.
Acinolaemus cryptidentatus from Mae Hong Son Province. A. Specimen CUMZ 15362 with enlarged aperture from three different angles; B. Protoconch and earlier whorl sculptures. An asterisk () indicates small lamella and plica situated in posterior chamber.*
Diagnosis.
Shell concave-conical, peristome weakly expanded, rectangular reticulated sculpture present on shell surface. Apertural dentitions nine: three on parietal wall, three on palatal wall, one basal, and two on columellar wall. Palatal tubercle prominent and located between upper- and inter-palatal plicae.
Re-description.
Shell weakly concave-conical, colourless; spire high and growing regularly; last whorl expanded. Shell height 1.6–1.8 mm and shell width 1.5–1.6 mm. Apex blunt; protoconch ~2 whorls with conspicuous spiral striae. Whorls ~4–5, bluntly shouldered, and convex; suture wide, well impressed, and deep. Shell surface sculptured with equidistantly spaced spiral striae, crossed with discontinuous and irregular radial ridge-like growth lines making rectangular reticulated sculpture throughout shell. Penultimate whorl regularly coiled to slightly sunken into last whorl. Last whorl rounded to weakly shouldered and flattened below periphery. Aperture subovate; peristome weakly expanded. Aperture with eight dentitions. Parietal wall with three lamellae: parietal lamella large, outer part low and reaching peristome edge, inner part strongly developed with long tall ridge located deeper inside aperture; infraparietal lamella prominent with a high ridge; angular lamella slightly curved, reaching peristome edge, a strong, tall ridge, long deeper inside aperture and with deep incision medially. Palatal tubercle strong, triangular, and situated on peristome edge between upper- and inter-palatal plicae. Three tiny and low plicae present in sinulus. Palatal wall with three plicae: upper-, inter-, and lower-palatal plicae with tall ridges and situated slightly inside aperture. Lower palatal plica tall and more prominent than upper- and inter-palatal plicae. Basal plica prominent with tall ridge. Columellar wall with two lamellae: columellar lamella is a very tall and distinct ridge; subcolumellar lamella a strong ridge and almost same size as basal plica. Umbilicus widely perforated, ~⅓ of shell width, rounded and deep.
Distribution.
Acinolaemus cryptidentatus is known from the type locality at Tham Wua Cave, Mae Hong Son Province in northern Thailand (Changlom et al. 2019), and subsequently reported from Shan State, Myanmar (Tongkerd et al. 2024).
Remarks.
The specimens from Mae La Na Cave in Pang Mapha District, which is ca 15 km east of the type locality are almost identical to the type specimen in shell shape, sculpture, and most of the apertural dentitions. However, these specimens have a prominent subcolumellar lamella, which was probably overlooked in the original description.
Acinolaemus
mueangonensis
Taxon classificationAnimaliaStylommatophoraHypselostomatidae
Changlom, Chan-ard & Dumrongrojwattana, 2019
6482881F-8AD2-5706-BCF8-AA1AB433E805
Acinolaemus mueangonensis Changlom, Chan-ard & Dumrongrojwattana, 2019: 159–161, fig. 3. Type locality: Tham Mueang On [Mueang On Cave], Mae On District, Chiang Mai Province, Thailand. Acinolaemus muangonensis [sic]—Changlom et al. 2019: 155 (abstract), 160 (figure caption).
Material examined.
Thailand • 8 shells (Fig. 11A, B); Mueang On Cave, Ban Sa Ha Khon, Mae On District, Chiang Mai Province; 18°47'13.2"N, 99°14'16.6"E; S. Panha leg.; CUMZ 15361 [type locality]. • 53 shells (Figs 12A, 13B, C); Phra Phutthabat Doi Khao Nam, Ban Na Subdistrict, Sam Ngao District, Tak Province; 17°14'54.7"N, 98°56'17.1"E; P. Tongkerd leg.; CUMZ 14455.1. • 1 shell (Fig. 12B); same data as preceding; CUMZ 14455.3. • 10 specimens in ethanol (Fig. 12C, D); same data as preceding; CUMZ 14455.4.
Acinolaemus mueangonensis from the type locality in Chiang Mai Province. A. Specimen CUMZ 15361 with enlarged aperture from three different angles; B. Protoconch and earlier whorl sculptures. An asterisk () indicates small lamella and plica situated in posterior chamber.*
Acinolaemus mueangonensis from Tak Province. A. Specimen CUMZ 14455.1 under light microscope showing brownish shell and plicae; B. Specimen CUMZ 14455.3 with enlarged aperture from two different angles; C, D. Living snails CUMZ 14455.4: C. Snail crawling on Hypselostoma pendulum; D. Crawling on wet paper towel (shell width ~1.8 mm). An asterisk () indicates small lamella and plica situated in posterior chamber.*
Diagnosis.
Shell concave-conical, brownish and with rectangular reticulated sculpture. Apertural dentitions nine, long and reaching peristome edge: three on parietal wall, three on palatal wall, one basal and one on columella wall. Palatal tubercle continuous with upper palatal plica.
Re-description.
Shell conical, concave-sided, brownish to pale brown; spire low to high, growing regularly, and sometimes slightly bent; last whorl expanded. Shell height 1.8–1.9 mm and shell width 1.7–1.8 mm. Apex blunt; protoconch spirally striated, ~2. Whorls ~5–6, rounded and convex; suture wide and well impressed. Shell surface sculptured with equidistantly spaced spiral striae, crossed with discontinuous and irregular radial growth lines making rectangular reticulated sculpture throughout shell. Penultimate whorl regularly coiled or sunken into last whorl. Last whorl bluntly shouldered and flattened below periphery. Aperture subcircular; peristome thickened and slightly expanded. Aperture with eight dentitions and with strong knobs reaching peristome edge. Parietal wall with three lamellae: parietal lamella large, outer part low, and inner part strongly developed and high, long deeper inside aperture; infraparietal lamella long and low; angular lamella prominent, high, long deeper inside aperture, and with slightly narrow and low ridge in middle. Palatal tubercle triangularly shaped, situated on peristome edge and connected to upper palatal plica. One or two tiny and low plicae (not reaching peristome edge) may be present in sinulus. Palatal wall with three plicae: upper-, inter-, and lower-palatal plicae ending on peristome edge as prominent tubercles, then continuing as narrow and low, but becoming strong deeper inside aperture. Basal plica low and long deeper inside aperture. Columellar lamella strong, high, and continuing deep inside aperture. Umbilicus widely perforate, ~⅓ of shell width, rounded and deep.
Living animal.
Snails are typically stylommatophoran with two pairs of tentacles. Upper tentacles are long, stout, cylindrical tubes, dark greyish, and with dark eye spots on the tip (Fig. 12C, D). Lower pairs are very short to knob-shaped and can be seen clearly in full extension in moving snails. Animal with short body, anterior-dorsal side with grey to blackish pigmentation, and posterior body and foot pale greyish to semi-translucent. The snails tend to cover their shell with soil, mud, or dirt.
Distribution.
Acinolaemus mueangonensis has a distribution beyond its type locality in northern Thailand (Changlom et al. 2019), since during this study several specimens were collected in soil samples taken at the base of limestone cliffs and outcrops in Tak Province, ca 170 km south of the type locality.
Remarks.
The specimens from the type locality examined herein have shell sculpture and apertural dentition similar to the original description and the type specimens (Changlom et al. 2019), except for the presence of a tiny plica in the sinulus, which was not mentioned in the original description.
The specimens from central Thailand (Tak population) show the same distinguishing characters as the type specimens: a brownish shell, rectangular reticulated sculpture, and long denticles deep inside the aperture and becoming strong knobs when reaching the peristome edge. In addition, the parietal lamella has a low ridge near the peristome that becomes a tall ridge inside the aperture, and the angular lamella has a high ridge with a narrow and low ridged incision in the middle. However, the central Thailand population (Fig. 12A, B) differed from typical specimens (Fig. 11A, B) in having a more concave-sided shell, a more elevated and slender spire (height 2.1–2.2 mm and width 1.7–1.8 mm), and the penultimate whorl slightly sunken into the last whorl. Additionally, the central Thailand population has a basal plica instead of a subcolumellar lamella, while a typical shell has a subcolumellar lamella instead of a basal plica. However, the distinction between the subcolumellar lamella and the basal plica is sometimes difficult and not a reliable character for species distinction and is subject to intraspecific variability. Therefore, we provisionally recognise the central population as conspecific with A. mueangonensis, since no concrete difference in morphology could be observed. DNA sequence data are needed to resolve this ambiguity.
Acinolaemus
corusticorus
Taxon classificationAnimaliaStylommatophoraHypselostomatidae
Tongkerd & Panha sp. nov.
B875F10A-A735-5F09-93EA-1DD2F53525CA
https://zoobank.org/78BD0129-3EAB-4593-AA77-8FCED5DCD0DA
Type material.
Holotype. Thailand • height 2.2 mm, width 1.9 mm (Fig. 14A, B); Phra Phutthabat Doi Khao Nam, Ban Na, Sam Ngao District, Tak Province; 17°14'54.7"N, 98°56'17.1"E; P. Tongkerd leg.; CUMZ 15363.1. Paratypes. Thailand • 2 shells (Fig. 15A, B); same data as for holotype; CUMZ 15363.2. • 45 shells (Fig. 13D, E); same data as for holotype; CUMZ 14455.2. • 2 shells; same data as for holotype; NHMUK 20250360. • 2 shells; same data as for holotype; SMF.
A. Synoptic photo of sympatric Acinolaemus species, specimens sorted from soil sample collected at Tak Province; B, C. Acinolaemus mueangonensis, specimen CUMZ 14455.1 with brownish shell in lateral and apertural views showing long plicae visible throughout the translucent shell; D, E. Acinolaemus corusticorus sp. nov., paratype CUMZ 14455.2 with colourless shell in lateral and apertural views showing short plicae visible throughout the translucent shell.
Acinolaemus corusticorus sp. nov. from Tak Province. A, B. Holotype CUMZ 15363.1 with enlarged aperture from three different angles and B. Protoconch and earlier whorl sculptures.
Acinolaemus corusticorus sp. nov. from the type locality. A. Paratype CUMZ 15363.2 with enlarged aperture from two different angles (without basal plica); B. Paratype CUMZ 15363.2 with enlarged aperture from two different angles (with basal plica).
Diagnosis.
Shell concave-conical, colourless, and with rectangular reticulations. Nine apertural dentitions, reaching peristome edge: three on parietal wall, palatal tubercle, four on palatal wall, and two on columellar wall. Palatal tubercle continuous with upper palatal plica.
Description.
Shell concave-conical, colourless; spire high and growing regularly; last whorl expanded. Shell height 2.1–2.3 mm and shell width 1.8–1.9 mm. Apex blunt; protoconch ~2 whorls with fine spiral striae. Whorls 4–5, rounded and convex; suture wide and well impressed. Shell surface sculptured with equidistantly spaced spiral striae, crossed with irregular radial growth lines making a rectangular reticulated sculpture throughout shell. Penultimate whorl regularly coiled; last whorl bluntly shouldered and flattened below periphery. Aperture subovate; peristome thickened and slightly expanded. Aperture with nine or ten dentitions and with more or less strong knobs reaching peristome edge. Parietal wall with three lamellae: parietal lamella large, strongly developed, and high and long deeper inside aperture; infraparietal lamella long and low; angular lamella relatively smaller than parietal lamella, long, low near peristome edge, somewhat sinuous, and higher deeper inside aperture. Palatal tubercle strongly developed with triangular shape and continuous with upper palatal plica. Palatal wall with four plicae: upper-, inter-, lower-, and infra-palatal plicae connecting with peristome in the form of prominent tubercles then continuing as narrow and low, becoming strong and high inside aperture. Inter- and lower-palatal plicae much larger than upper palatal plica, infrapalatal plica smallest or may be absent (Fig. 15A). Basal plica may be present with small and low ridge. Columellar wall with two lamellae: columellar lamella strong and distinct, continuing deep inside aperture; subcolumellar lamella small, low. Umbilicus widely perforate, ~⅓ of shell width, rounded and deep.
Differential diagnosis.
This new species can be distinguished from A. cryptidentatus from northern Thailand by having major dentitions (on parietal, palatal, and columellar walls) that become strong knobs when reaching peristome edge, palatal tubercle continuous with upper palatal plica, and without tiny plicae inside the sinulus. In comparison, A. cryptidentatus possesses dentitions that do not reach the peristome edge, a palatal tubercle situated between upper- and inter-palatal plicae, and with two tiny plicae inside the sinulus.
Acinolaemus corusticorus sp. nov. is similar to A. dayanum (Stoliczka, 1871) from Myanmar and A. mueangonensis from northern Thailand in shell form and sculpture. It differs by having a colourless shell without cervical crest (a swelling or convex ridge on the last whorl behind the expanded lip); parietal lamella has a thick and high ridge; angular lamella has a continuously high ridge and without incision; palatal plicae short (<½ of last whorl length when seen from lateral view; Fig. 13D, E), palatal plicae continue from knobs on peristome edge with short, narrow, and low ridges, and then becoming high inside the aperture. For comparison, A. dayanum and A. mueangonensis have palatal plicae continuing from peristome knobs, which are long and narrow but low ridges before becoming folds inside aperture. Acinolaemus mueangonensis possesses a brownish and conical to elevated conical shell; parietal lamella has a low ridge near the peristome then becoming a tall ridge inside; angular lamella has a high ridge with narrow and low ridge in middle; palatal plicae long (>½ of last whorl length when seen from lateral view; Fig. 13B, C). Acinolaemus dayanum has a conical low spire with a cervical crest (a swelling or convex ridge on the last whorl behind the expanded lip); parietal lamella has a low ridge near peristome edge then gradually becoming a tall ridge inside; angular lamella has a high ridge near peristome edge and then gradually becoming a low ridge inside aperture.
Acinolaemus ptychochilus from northern Thailand also clearly differs from A. corusticorus sp. nov. in shape of parietal and angular lamellae, and in having four palatal plicae (upper-, inter-, hooked lower-, and infra-), a small basal plica, and three columellar lamellae; palatal tubercle situated between upper- and inter- palatal plicae. Acinolaemus rhamphodon Thompson & Upatham, 1997 and A. stenopus Thompson & Upatham, 1997 differ from A. corusticorus sp. nov. in having fewer dentitions, a very strong angular lamella, and a weak parietal lamella. Both species also possess two palatal plicae (upper- and lower-), and a columellar lamella. Finally, A. rhamphodon has a hooked columellar lamella, while A. stenopus has an elevated shell with a high spire, and an enlarged angular lamella and palatal plica that nearly enclose the sinulus.
Distribution.
Acinolaemus corusticorus sp. nov. is known only from the type locality. At this locality, the species is sympatric with five other hypselostomatid species: Hypselostoma pendulum (Panha & Burch, 2002), H. khaowongensis Panha, 1998, A. mueangonensis, A. rhamphodontis sp. nov., and Krobylos takensis Panha & Burch, 2004 (Panha 1998b; Panha et al 2004).
Etymology.
The specific name corusticorus is from two Latin words co- meaning ‘together or with’ and rusticor meaning ‘living in the country’, referring to the new species being sympatric with two other congeners.
Remarks.
Although this new species occurs sympatrically with A. mueangonensis, under the microscope it is evident that they are two distinct species. Acinolaemus corusticorus sp. nov. differs from sympatric congeners by having a colourless, short, and stout shell (Fig. 13A), while A. mueangonensis has a much smaller and more slender shell, and with pale brownish to brownish shell colour. In addition, these two species are clearly distinct in number and morphology of apertural dentitions.
Acinolaemus
rhamphodontis
Taxon classificationAnimaliaStylommatophoraHypselostomatidae
Tongkerd & Panha sp. nov.
93B43A14-CFBA-50E4-B61C-F690F5141DB2
https://zoobank.org/848D50A4-5B1B-457A-988A-0D07445ECDDC
Type material examined.
HoIotype. Thailand • height 1.3 mm (including tuba), width 1.4 mm (Fig. 16A, B); Phra Phutthabat Doi Khao Nam, Ban Na subdistrict, Sam Ngao District, Tak Province; 17°14'56.4"N, 98°56'16.3"E; Tongkerd leg.; CUMZ 14449. Paratypes. Thailand • 3 shells (Fig. 17A–C); same data as for holotype; CUMZ 14450. • 1 adult + 1 juvenile in ethanol (Fig. 17D–F); same data as for holotype; CUMZ 14451 (COI accession number PV698334–PV698335). • 37 adults + 3 juveniles (COI accession nos. PV698334, PV698335); same data as for holotype; CUMZ 14452. • 2 shells; same data as for holotype; NHMUK 20250361. • 2 shells; same data as for holotype; SMF.
Acinolaemus rhamphodontis sp. nov., holotype CUMZ 14449 from Tak Province. A. Shell with enlarged aperture from two different angles; B. Protoconch and earlier whorl sculptures.
Acinolaemus rhamphodontis sp. nov., from Tak Province. A. Paratype CUMZ 14450 with enlarged aperture; B. Juvenile paratype CUMZ 14450 from the type locality; C. Natural broken shell with dentition on parietal and palatal walls as seen from inside the shell; D–F. Specimen CUMZ 14451 of living snails crawling on wet paper towel (different angles of same individual; shell width ~1.4 mm).
Diagnosis.
Shell concave-conical, with long and descending tuba, peristome expanded. Apertural dentitions eight, all longer inside aperture: infra parietal, parietal, angular, upper- and lower-palatal, palatal tubercle, basal, subcolumellar lamella, and columellar lamella. Umbilicus wide.
Description.
Shell concave-conical, colourless; spire depressed and growing regularly; last whorl broadly expanded. Shell height 1.3–1.4 mm (including tuba) and shell width 1.3–1.5 mm. Apex large and rounded; protoconch ~2 whorls and with prominent spiral striae. Whorls ~4 (excluding tuba), rounded and convex; suture wide, well impressed, and deep. Shell surface sculptured with equidistantly spaced spiral striae (14–16 on body whorl in frontal view), crossed with discontinuous narrow radial growth lines making a rectangular reticulated sculpture throughout shell. Penultimate whorl regularly coiled; last whorl rounded; tuba long, ~¼ whorl or less, strongly descending and curving. Aperture subovate, open ventrally to subventrally; peristome thin, weakly expanded, and with thin depression area on parietal side. Apertural dentitions eight and all knob-shaped when reaching peristome edge. Parietal wall with three lamellae: parietal lamella long, outer part low, and then gradually becoming strong, thickened, and high inside aperture; infraparietal lamella evenly low; angular lamella strong. Palatal tubercle prominent, triangular, situated at peristome lip, and continuous with upper palatal plica. Palatal wall with two plicae: upper- and lower-palatal plicae connecting with peristome edge in form of prominent knobs, then continuing as narrow and low deeper inside aperture, eventually becoming stronger and higher at innermost ends. Basal plica narrow, and low. Columellar wall with two lamellae: columellar lamella prominent with tall ridge; subcolumellar lamella very small and low. Umbilicus widely opened, occupying ~½ of shell width and showing all preceding whorls.
Living animal.
Snail typically stylommatophoran with two pairs of tentacles. Upper tentacles are long, stout, cylindrical tubes, colourless to translucent, with dark eye spots on the tip. Lower pairs are very short to knob-shaped (difficult to observe in living snails). Animal with short body, anterior-dorsal side brownish while posterior body and foot paler to translucent. The snails tend to cover their shell with soil, mud, or dirt (Fig. 17D–F).
Distribution.
This new species is known only from the type locality, where the empty shells and one living snail were found in the soil, leaves, and twig litter at the base of a limestone cliff. The type locality is an island located in the reservoir of Bhumibol Dam, ca 45 km northwest of Tak Town. This island is ca 400 m long and 200 m wide and aligned north-south; the dry shells and the specimen were collected from the eastern slope of the island. The hills are low, with scattered land used by temples and with some exposed limestone rocks and cliffs. The vegetation on this island is dominated by low, dry, dipterocarp forest.
Differential diagnosis.
The shell of A. rhamphodontis sp. nov. is most similar to A. dayanum from Myanmar, and A. cryptidentatus, A. mueangonensis and A. ptychochilus from Thailand. They all share a rectangular reticulated shell sculpture, many long dentitions that reach the peristome edge where they form small denticles (except in A. cryptidentatus) and have a palatal tubercle (Thompson and Upatham 1997; Changlom et al. 2019; Tongkerd et al. 2024). The differences are that A. rhamphodontis sp. nov. has a long and descending tuba and a spire sunken into the last whorl. In contrast, the other four species have no tuba, and have a conical spire with straight to curved sides.
Acinolaemus rhamphodontis sp. nov. differs from A. carcharodonVermeulen et al., 2007 and A. pyramidalis (Vermeulen et al., 2007) from the Mekong Delta limestone hills in Vietnam, and A. rectusVermeulen et al., 2019 from Cambodia in having a long and descending tuba, long dentitions that reach the peristome edge, and a rectangular reticulated shell surface. In comparison, these three species possess a very short tuba, prominent radial ridges, and dentitions that are short and situated inside to deep inside the aperture. Additionally, A. carcharodon has a sunken spire and four dentitions (parietal, hooked angular, palatal, and basal), A. pyramidalis has a conical spire and four to five dentitions (parietal, angular, two palatals, and a very inconspicuous columellar), and A. rectus has a conical spire, last whorl rounded, tuba slightly ascending and five dentitions (parietal, angular, two palatals, and columellar) (Vermeulen et al. 2007, 2019).
This new species has a long descending tuba similar to several species in the Clostophis sankeyi species group. It differs by having rectangular reticulated shell sculpture, with eight apertural dentitions (parietal, infra parietal, angular, two palatals, basal, and two columellar) reaching the peristome edge, and with a palatal tubercle present. Furthermore, A. rhamphodontis sp. nov. has 14–16 dash-like spiral striae on the last whorl and the penultimate whorl not sunken, whereas C. sankeyi, C. proboscideus, C. yoga, and C. udayaditinus have 18–20 or more continuous spiral striae on the last whorl, and the penultimate whorl sunken into the last whorl. Clostophis yoga also has a narrower umbilicus, <1/3 of the shell width, than the new species (Benson 1860; Páll-Gergely et al. 2020; Páll-Gergely and Hunyadi 2022; Sutcharit et al. 2025). Finally, A. rhamphodontis sp. nov. also differs from C. rhynchotes sp. nov. by having a rectangular reticulated shell surface and dash-like spiral striae on the last whorl, while C. rhynchotes sp. nov. has a weak reticulated shell surface and continuous spiral striae on the last whorl.
Etymology.
The specific name rhamphodontis is from two Greek words: rhamphos meaning ‘curving beak’ and odontos meaning ‘tooth’. Together they refer to the tuba that curves downward and the many dentitions in the aperture.
Remarks.
The presence of a rectangular reticulated shell sculpture and the many dentitions reaching the peristome edge where they form small denticles and extend inside the aperture, clearly position A. rhamphodontis sp. nov. within the genus Acinolaemus (Thompson and Upatham 1997).
Discussion
The report of these three new species from central Thailand raises some interesting questions regarding the generic boundaries and relationships between Clostophis and Acinolaemus. In the original description, Acinolaemus was characterised by an auriculate-shaped aperture, 1–3 dentitions on the parietal wall, an enlarged and conspicuous angular lamella, a well-developed sinulus (formed by angular lamella and palatal plica), and a spirally striated protoconch (Table 4; Thompson and Upatham 1997; Schileyko 1998). However, these diagnostic characters are not consistently expressed across all species assigned to the genus, casting doubt on their reliability for the diagnosis of the genus. For example, six species, i.e., A. cryptidentatus, A. dayanum, A. mueangonensis, A. ptychochilus (type species), A. rhamphodontis sp. nov., and A. corusticorus sp. nov. exhibit many dentitions that reach the peristome edge and extend deeper inside the aperture as long dentitions, a well-developed palatal tubercle, and with distinct rectangular reticulated shell sculptures (Thompson and Upatham 1997; Changlom et al. 2019; Tongkerd et al. 2024). In the present study, A. mueangonensis and A. rhamphodontis sp. nov. appeared as well-supported sister taxa, while A. sphinctinion Thompson & Upatham, 1997, A. colpodon Thompson & Upatham, 1997, A. stenopus, and A. rhamphodon possess only a few apertural dentitions and a palatal wall with one or two lamellae. Our phylogeny indicates that A. rhamphodon and A. colpodon form a clade with C. rhynchotes sp. nov. and C. udayaditinus, with support from the ML analysis. Interestingly, these four species share certain morphological characters such as prominent spiral striations relative to the growth lines and few apertural dentitions, which are the distinguishing characters of the Clostophis (Páll-Gergely et al. 2020). Moreover, three other species, namely A. carcharodon, A. pyramidalis and A. rectus, possess a short to long tuba, four or five apertural dentitions, and prominent spiral striations (Vermeulen et al. 2007, 2019) and these conchological characters suggest a close relationship between Acinolaemus and Clostophis, with this latter genus being defined by a colourless shell, short to long tuba, prominent spiral striations, and typically with one or more apertural dentitions (Páll-Gergely et al. 2020). Previous DNA studies by Tongkerd et al. (2004) suggest that the shell surface sculpture provides a more informative and reliable phylogenetic signal than shell shape or dentition, and this was taken into consideration in the Gojšina et al. (2025) revision. However, due to the limited number of genera and species examined in the Hypselostomatidae, it remains premature to expand further on the relationship between Clostophis and Acinolaemus.
Supplementary Material
XML Treatment for Clostophis
XML Treatment for Clostophis proboscideus
XML Treatment for Clostophis rhynchotes
XML Treatment for Acinolaemus
XML Treatment for Acinolaemus ptychochilus
XML Treatment for Acinolaemus cryptidentatus
XML Treatment for Acinolaemus mueangonensis
XML Treatment for Acinolaemus corusticorus
XML Treatment for Acinolaemus rhamphodontis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Benson WH (1860) On Clostophis and Rhiostoma, new Burmese genera of land-shells.Annals & Magazine of Natural History 5(26): 95–97. 10.1080/00222936008697183 · doi ↗
- 2Changlom B Chan-ard T Dumrongrojwattana P (2019) Two new species of the genus Acinolaemus (Pulmonata: Pupillidae) from Thailand.Thailand Natural History Museum Journal 13(2): 155–161. https://journal.nsm.or.th/en/node/175
- 3Clements R Sodhi NS Thuizen MS Ng PKL (2006) Limestone Karsts of Southeast Asia: Imperiled arks of biodiversity. Bioscience 56(9): 733–742. 10.1641/0006-3568(2006)56[733:LKOSAI]2.0.CO;2 · doi ↗
- 4Collinge WE (1902) On the non-operculated land- and freshwater molluscs collected by the members of the “Skeat Expedition” in the Malay Peninsula, 1899–1900.Journal of Malacology 9(3): 71–95. 10.1111/j.1469-7998.1902.tb 08186.x · doi ↗
- 5Folmer O Black M Hoeh W Lutz R Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates.Molecular Marine Biology and Biotechnology 3: 294–299.7881515 · pubmed ↗
- 6Gojšina V Hunyadi A Sutcharit C Tongkerd P Auffenberg K Grego J Vermeulen JJ Reischütz APáll-Gergely B (2025) A new start? Revision of the genera Anauchen, Bensonella, Gyliotrachela and Hypselostoma (Gastropoda, Eupulmonata, Hypselostomatidae) of Southeast Asia with description of 46 new species.Zoo Keys 1235: 1–338. 10.3897/zookeys.1235.14528140321358 PMC 12044344 · doi ↗ · pubmed ↗
- 7Hoekstra P Schilthuizen M (2011) Phylogenetic relationships between isolated populations of the limestone-dwelling microsnail Gyliotrachela hungerfordiana (Gastropoda: Vertiginidae).Journal of Zoological Systematics and Evolutionary Research 49(4): 266–272. 10.1111/j.1439-0469.2011.00623.x · doi ↗
- 8Jirapatrasilp P Tongkerd P Sutcharit C (2023) Somsak Panha and his contribution to Southeast Asian biodiversity research.Tropical Natural History, Supplement 7: 1–30. 10.58837/tnh.23.7.257693 · doi ↗