Multi-oocyte follicles in the bovine ovary: occurrence, activation, and growth during early in vitro folliculogenesis in a dynamic culture bioreactor
Andrea Candela, Vincenza De Gregorio, Vincenzo Genovese, Angela Travaglione, Mario Cimmino, Riccardo Talevi, Roberto Gualtieri

TL;DR
This study explores multi-oocyte follicles in cow ovaries, showing they can develop in vitro and likely originate before birth.
Contribution
The study provides evidence for the prenatal origin and functional capacity of multi-oocyte follicles in bovine ovaries.
Findings
MOFs can activate and develop to secondary stages in vitro, similar to single-oocyte follicles.
MOF frequency remains stable after in vitro culture, suggesting a prenatal origin.
Differences in activation rates and oocyte dynamics were observed between MOFs and SOFs.
Abstract
Multi-oocyte follicles (MOFs) have been identified in various mammals, including humans, yet their origin and function remain controversial. This study aimed to investigate the occurrence and early developmental dynamics of MOFs in bovine ovaries, comparing them to single-oocyte follicles (SOFs). Ovarian tissues from Bos taurus taurus individuals, including one case with an unusually high MOF incidence, were cultured in vitro under dynamic conditions. MOF frequency and their progression through early folliculogenesis stages were assessed histologically and via confocal microscopy. MOFs were observed at varying frequencies, with one individual showing a notably high incidence (15.8%). In vitro culture confirmed that MOFs can activate and progress to secondary stages, similar to SOFs. However, differences in activation rates and oocyte number dynamics were noted between MOFs and SOFs,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
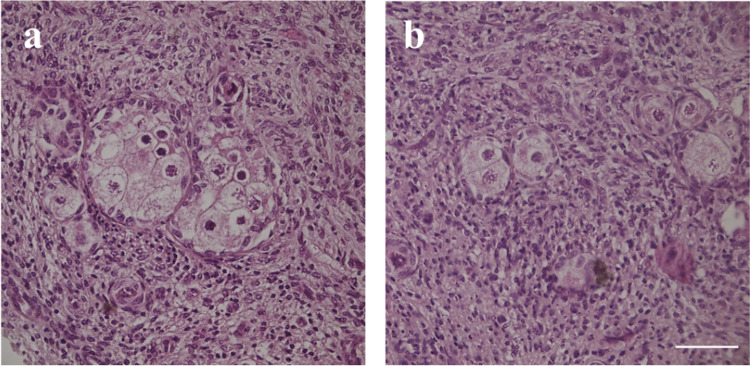 Figure 1
Figure 1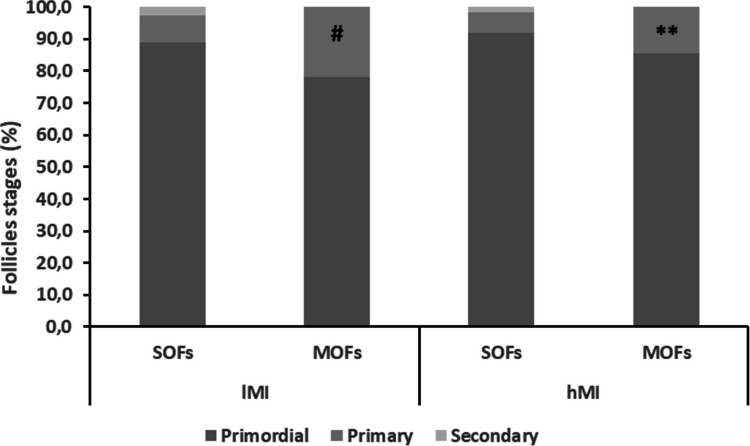 Figure 2
Figure 2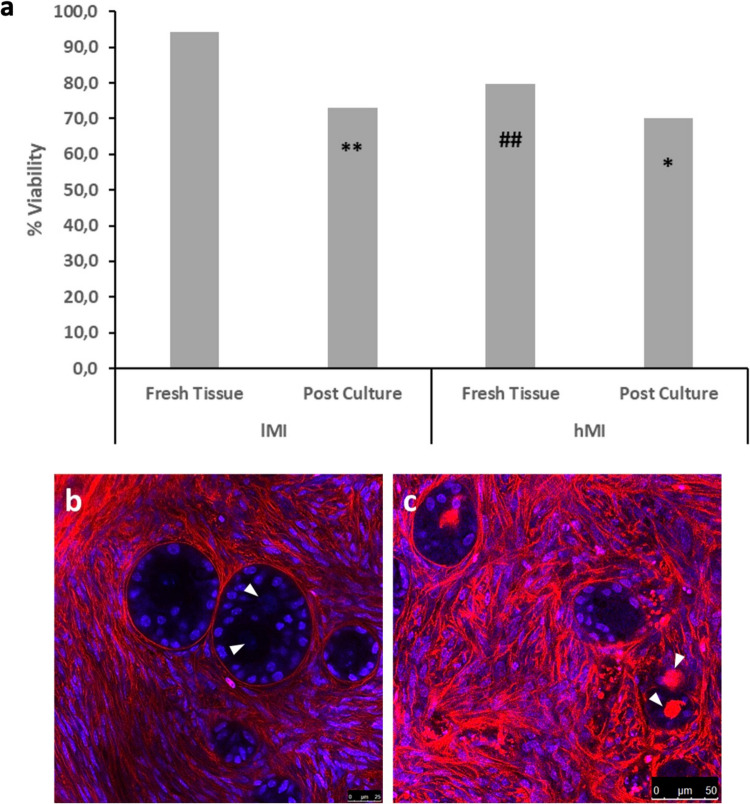 Figure 3
Figure 3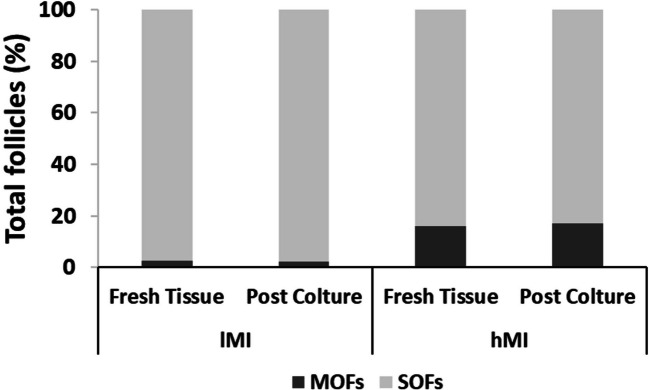 Figure 4
Figure 4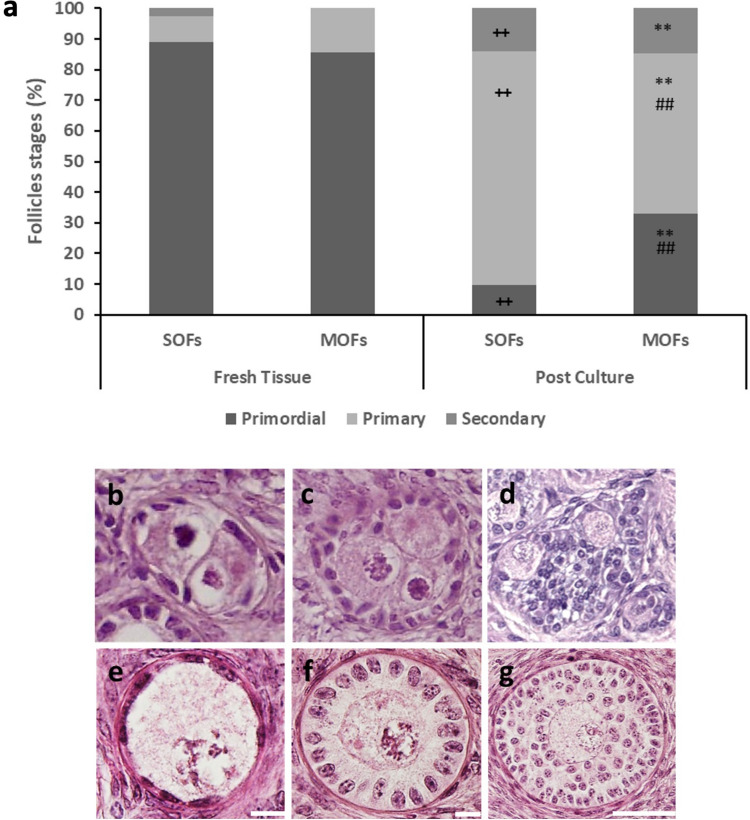 Figure 5
Figure 5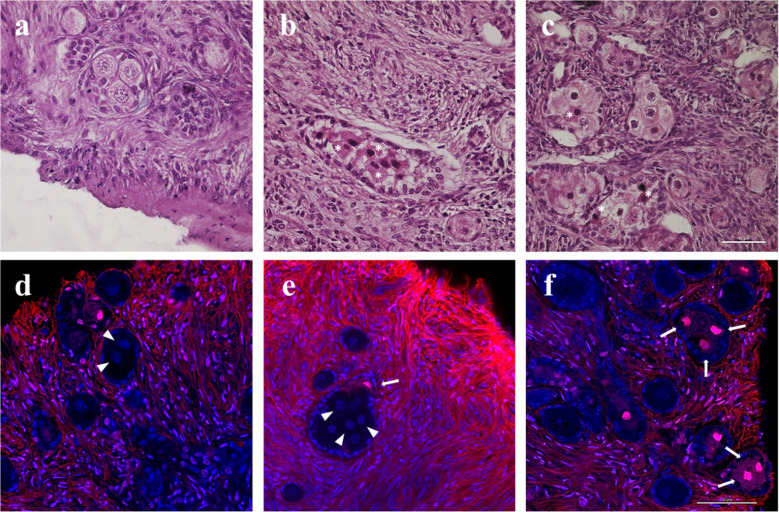 Figure 6
Figure 6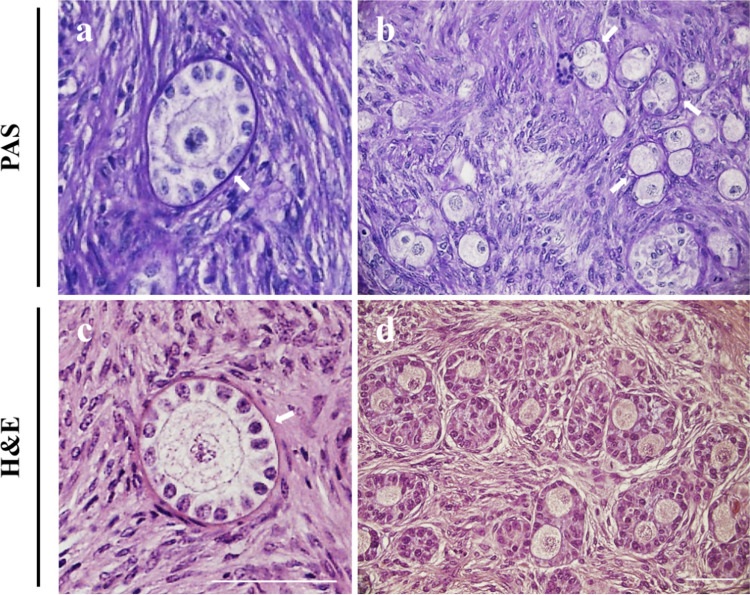 Figure 7
Figure 7- —http://dx.doi.org/10.13039/100020956Merck Healthcare KGaA
- —Università degli Studi di Napoli Federico II
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive Biology and Fertility · Ovarian function and disorders · Sperm and Testicular Function
Introduction
During female sex differentiation, oogonia undergo mitosis followed by incomplete cytokinesis giving rise to nests of germ cells connected through intercellular bridges. After initiation of meiosis and arrest at the prophase diplotene stage, during the second trimester in humans, and after birth in mice, germ-cell nest break down and flattened pregranulosa cells surround each oocyte to form primordial follicles [1, 2]. However, primordial follicles consisting of two or more oocytes enveloped by granulosa cells and a single follicle basal lamina, termed multiple-oocyte follicles (MOFs), have been reported in several mammals [3, 4]. Three main hypotheses have been proposed to explain the origin of MOFs. They could generate as consequence of (1) fusion of individual follicles during folliculogenesis due to ruptures of follicle basal lamina and the invasiveness of granulosa cells [5–7], (2) failure of germ-cell nest breakdown [8, 9], or (3) MOFs could represent a natural polymorphism, stemming from a range of potential numerical combinations of oocytes and pregranulosa cells [10]. The occurrence of a low number of MOFs is a common feature in humans [4], mice [9], rabbits [11], pigs [12], sheeps [13], and cattle [14], while in other mammals such as cotton rats [15] and dogs [16], MOFs can account up to the 40% of the ovarian follicle population. In Bos taurus taurus females, the prevalence of MOFs ranges from 0.3 to 5.4% [17]. Although MOFs have often been regarded as pathological entities, several reports in the human indicate that MOFs can successfully complete their growth and meiotic maturation giving rise to normal metaphase II oocytes. Mature oocytes enclosed in the same zona pellucida, termed conjoined oocytes, have been rarely retrieved by binovular MOFs in women undergoing assisted reproduction [18–20] and were capable of giving rise to live births [21–23]. In most cases, the size and maturation stage of oocytes in binovular follicles were asynchronous, though Magdi et al. (2020) reported a case of dizygotic twinning live births from two mature conjoined oocytes after intracytoplasmic sperm injection (ICSI) [24]. Far less is known on the dynamic of early folliculogenesis in MOFs compared to the population of SOFs. In the human, MOFs containing 2 or 3 oocytes were observed in 98% of 117 ovaries of women aged 18–52 years, and their frequency (0.06–2.44%) was not age-dependent [25]. Moreover, though MOFs were reported to undergo atresia before puberty, they were suggested to do so at rates similar to SOFs during the adult life-span [25–28]. In bitches, cows, and rabbits, MOFs have been observed in all stages of folliculogenesis, from primordial to peri-ovulatory follicles [11, 17]. The main aims of the present study were to determine the ability of primordial MOFs to activate and successfully progress to the primary and secondary follicle stages compared to the activation and growth dynamics of SOFs. We recently established a dynamic in vitro culture system for bovine and human ovarian tissue in a perifusion bioreactor (PB) that significantly improves follicle quality and viability, percentage and health of secondary follicles, overall tissue health, and steroid secretion compared to conventional static culture [29, 30]. Herein, we adopted such an in vitro culture system to study the incidence of MOFs and SOFs in fresh and cultured bovine ovarian cortical tissue and followed follicle activation, growth, and viability in tissues recovered from different individuals, including one subject with an unusually high occurrence of MOFs, which has not been previously observed in Bos taurus taurus. Our findings shed light on the dynamic and physiological function of MOFs during early folliculogenesis in the bovine animal model.
Materials and methods
Chemicals and consumables
Leibovitz’s L-15 medium, α-MEM GlutaMAX medium, insulin transferrin selenium (ITS) 100 ×, Live/dead Fixable Far-Red Stain were purchased from Invitrogen (Milan, Italy). Penicillin streptomycin 100 ×, amphotericin B 250 μg/mL, bovine serum albumin (BSA), L-ascorbic acid, L-glutamine 200 mM, propidium iodide, methanol, benzyl alcohol/benzyl benzoate (BABB), periodic acid Shiff (PAS), and eosin-Y were purchased from Sigma-Aldrich (Milan, Italy). Mayer’s haematoxylin and paraffin wax were from Carlo Erba (Milan, Italy). The tissue chopper was purchased from McIlwain, Mickle Laboratory Engineering Company Ltd. (Surrey, UK). Pump tubing was purchased from Cole-Parmer (Milan, Italy) and the peristaltic pump from Ismatec (Enco, Venice, Italy).
Collection and preparation of ovarian tissue and dynamic culture in PB
Bovine ovaries collected at the time of slaughter (Slaughterhouse Di Tella, San Marcellino, Caserta, Italy; CEE accreditation number 1403/M) from 4 individuals (n = 4; age 8–24 months) were transported within 2 h to the lab in Leibovitz’s L-15 supplemented with 1% penicillin streptomycin (P/S) and 1 μg/mL amphotericin-B at 4 °C. Ovaries were cut into cortical slices (~ 0.6-mm thick) using a tissue slicer and further dissected in cortical strips 1 mm × 1 mm × 0.5 mm through the use of a tissue chopper (McIlwain, Mickle Laboratory Engineering Company, Ltd., Surrey, UK). After two washes in handling medium, groups of 10 ovarian cortical strips from each individual were randomly distributed into a sterile perifusion bioreactor (PB) for dynamic culture at a flow rate of 4 mL/min as previously reported [29, 30]. The cortical strips were cultured for 14 days, replacing half the volume with fresh medium every 2 days. At the end of the dynamic culture, 6 cortical strips from each PB were collected and processed for histology and 4 strips for viability assessments. Fresh uncultured strips were processed for histology and viability assessment as day 0 controls.
Histology
Cortical strips fixed in Bouin’s and embedded in paraffin were cut into 5-μm-thick serial sections and stained with haematoxylin and eosin or with periodic acid Schiff (PAS). Follicle staging was evaluated by two blind expert observers and scored according to Gougeon’s criteria [31] as follows: primordial follicles have a single layer of flat GCs, primary follicles have a complete single layer of cuboidal GCs, and secondary follicles have two or more complete layers of cuboidal GCs. For PAS staining, sections were treated with periodic acid solution for 10 min at room temperature (RT), rinsed in distilled water, and immersed in Schiff’s reagent for 35 min in the dark. Slides were washed in sulfuruos acid solution (NaSO_3_ 10% w/v in 0.05 M HCl), rinsed for 5 min in tap water, counterstained in haematoxylin solution for 50 s, rinsed, dehydrated, cleared, and mounted in xylene-based mounting media.
Viability assessment
Strips were incubated, under shaking, for 3 h at 4 °C, in Dulbecco’s PBS (phosphate buffered saline) with 1 μg/mL Live/Dead Fixable Far-Red Stain, washed in fresh dPBS for 30 min, fixed in 4% paraformaldehyde for 2 h at RT, washed in fresh PBS, and incubated at 4 °C overnight in a permeabilization solution consisting of triton 2% in PBS supplemented with propidium iodide 10 μg/mL. The live/dead probe is resistant to fixation and reacts with free amines and is excluded by cells with intact membranes. Strips were then optically cleared using BABB clearing protocol [32]. Briefly, samples were serially incubated in solutions of 50%, 70%, 80%, 90%, 100%, and 100% methanol, each for 30 min subsequently in a 1:1 solution composed of methanol and BABB for 4 h, and finally in absolute BABB for 24 h. To avoid compression, strips were mounted in BABB solution on a glass slide with 3 spacer coverslips (0.17 mm) placed on each side of the samples and covered with a coverslip. Analysis was carried out with a Leica TCS SP5 confocal scanning laser microscope (Leica Microsystems, Wetzlar, Germany) using a 496-nm Argon laser for visualizing the nuclear label (propidium iodide) and a 633-nm helium–neon laser for the live/dead probe. Each strip was traversed using the z-position control, and fields to a depth of 300 μm from the tissue surface were imaged using a 63 × glycerol immersion objective.
Statistical analysis
For each experiment, data were reported as cumulative percentages. Statistical analysis was performed by Fisher’s exact test for pairwise comparisons when overall significance was detected.
Results
Histological analysis of fresh and cultured ovarian tissues of the four individuals analysed showed the presence of MOFs in all samples. In our experience on early folliculogenesis in vitro, MOFs are usually retrieved at a low frequency in bovine ovaries. One individual had a markedly high frequency of MOFs and has been henceforth termed as high MOF individual (hMI), compared to the remaining three individuals, termed low MOF individuals (lMI). A total number of 656 follicles was scored in fresh tissue of hMI, and 15.8% of the total follicle population was accounted by MOFs containing 2 to 10 oocytes (Table 1, Fig. 1a). A total number of 1162 follicles were scored in the three lMI fresh tissues, and MOFs, containing 2 to 3 oocytes accounted for the 2.7% of the total follicle population (Table 1, Fig. 1b). Table 1. Histological characterization of MOFs in hMI and lMI: MOF numbers, stages, and numbers of oocytes enclosed in folliclesMOFs/total folliclesFollicle stageNo. of oocyte per follicle2345678910hMI Fresh tissue104/656Primordial601973-----Primary251-411-1Secondary--------- Postculture82/485Primordial18711-----Primary261331-----Secondary84-------IMI Fresh tissue32/1162Primordial223-------Primary7--------Secondary--------- Postculture15/638Primordial43-------Primary71-------Secondary---------Fig. 1. Representative images of hMI (a) and 1MI (b) in fresh tissue. Bar = 50 µm
Histological analysis of fresh tissue samples demonstrated a similar high proportion of MOFs and SOFs at the primordial stage in hMI and lMI ovarian tissues (ns). However, the percentages of primary MOFs significantly exceeded those of primary SOFs in both lMI and hMI (Fig. 2).Fig. 2. Follicle stages in fresh tissue of 1MI and hMI. #, P < 00.05 versus primary SOFs of 1MI; **, P < 00.01 versus primary SOFs of hMI
In fresh hMI tissue, primordial MOFs (n = 89) contained 2–5 oocytes (Table 1). The percentages of primordial MOFs enclosing 2 to 5 oocytes were: 2, 67.4%; 3, 21.3%; 4, 7.9%; 5, 3.4%; mean primordial MOF oocyte number, 2.47. Primary MOFs (n = 15) contained 2 to 10 oocytes (Table 1), and the oocyte number per follicle was significantly higher compared to primordial follicles (P < 0.05). Percentages of primary MOFs enclosing 2 to 10 oocytes were: 2, 13.3%; 3, 33.3%; 4, 6.7%; 5, 0%; 6, 26.6%; 7, 6.7%; 8, 6.7%; 9, 0%; 10, 6.7%; mean primary MOF oocyte number, 4.80. Secondary MOFs were absent in fresh hMI tissue.
In lMI tissues, primordial MOFs (n = 25) contained 2–3 oocytes (Table 1; percentages of primordial MOFs enclosing 2 to 3 oocytes: 2, 88%; 3, 12%), and the mean primordial MOF oocyte number was 2.12. Primary MOFs (n = 7) enclosed 2 oocytes, and the mean primary MOF oocyte number was 2. Secondary MOFs were absent in fresh lMI tissues.
Confocal analysis of live-dead staining showed that 94.3% and 79.7% (P < 00.01) of follicles were viable in lMI and hMI tissues, respectively (Fig. 3a). Percentages of viable MOFs and SOFs were not significantly different in both hMI and lMI tissue samples as shown by representative confocal images (Fig. 3b, c).Fig. 3. Viability assessment. a Percentages of viable follicles before and after 14 days of dynamic culture in 1MI and hMI. *, P < 00.05 and **, P < 00.01 versus corresponding fresh tissue; ##, P < 00.01 versus 1MI|fresh tissue. b, c Representative confocal images of live (b) and dead (c) SOFs and MOFs. Blue, propidium iodide-stained nuclei; red, extracellular matrix, basal lamina, and dead oocytes stained by live-dead far-red; white arrowheads, germinal vesicles of two live (b) or dead (c) oocytes in MOFs. Bar = 25 µm (b). Bar = 50 µm (c)
Ovarian cortical strips were analysed after 14 days of dynamic culture in PB. The percentage of MOFs in lMI and hMI remained constant before and after culture (Fig. 4), suggesting that 1) no fusions among follicles occurred during activation and growth in vitro, and 2) MOFs did not have an increased tendency to undergo atresia compared to SOFs.Fig. 4SOFs and MOF percentages before and after culture in 1MI and hMI
In hMI cultured tissues, a significantly higher percentage of primordial and a decreased percentage of primary MOFs, compared to the proportions found in SOFs, were retrieved (Fig. 5a).Fig. 5a Follicle stages in hMI before and after culture. **, P < 00.1 versus corresponding MOF stages in fresh tissue; + +, P < 00.01 versus corresponding SOF stages in fresh tissue; ##, P < 00.01 versus corresponding SOF stages after culture. b–g Representative images of primordial (b, e), primary (c, f), and secondary (d, g) MOF and SOF, respectively. Bar = 10 µm (b, c, e, f). Bar = 50 µm (d, g)
Histological analysis of follicle stages (Fig. 5b–d, MOFs; Fig. 5e–g, SOFs) in fresh and cultured hMI tissue showed that the proportion of primordial SOFs dropped from 89 to 10%, whereas the percentage of primordial MOFs dropped from 80 to 35% after culture. This indicates that, although both follicle types activate their growth during culture, a significantly lower proportion of primordial MOFs (56.2%) were able to do so compared to primordial SOFs (88.8%) over the 14-day culture period. Although the percentage of primary SOFs significantly exceeded that of primary MOFs, the proportions of secondary SOFs and MOFs were similar suggesting a similar competence of the two follicle populations to progress to the secondary stage during culture (Fig. 5a). The dynamic of activation and growth in SOFs of lMI was similar to that shown in hMI. The dynamics of activation and progression in MOFs of lMI were not shown given the very low numbers of MOFs retrieved.
In hMI cultured tissue, primordial MOFs (n = 27) contained 2–5 oocytes (percentages of primordial MOFs enclosing 2 to 5 oocytes: 2, 66.7%; 3, 25.9%; 4, 3.7%; 5, 3.7%; mean primordial MOF oocyte number, 2.44), primary MOFs (n = 43) had 2–5 oocytes (percentages of primary MOFs enclosing 2 to 5 oocytes:2, 60.5%; 3, 30.2%; 4, 7%; 5, 2.3%; mean primary MOF oocyte number, 2.51), secondary MOFs (n = 12) had 2–3 oocytes (percentages of secondary MOFs enclosing 2 or 3 oocytes: 2, 66.6%; 3, 33.4%; mean secondary MOF oocyte number, 2.33). Numbers of oocytes in MOFs of lMI were not shown given the very low numbers of MOFs retrieved.
Total follicle viability significantly decreased after culture in both lMI (P < 00.01) and hMI (P < 00.05) tissues (Fig. 3a). Loss of viability of one or more oocytes in hMI MOFs was occasionally observed after culture both in histological (Fig. 6a–c) and confocal (Fig. 6d–f) assessments.Fig. 6. Representative histological and confocal images showing the health (a–c) and viability (d–f) of MOF oocytes in hMI. *, Eosinophilic, damaged oocytes; white arrowheads = germinal vesicles of live oocytes; white arrows, dead oocytes. Bar = 50 µm
No figures of basal lamina breakdown or invasion of granulosa cells from one follicle to the neighboring follicles were detected in confocal optical sections (Fig. 6d–f) and histological sections (Fig. 7a–d), confirming that the number of MOFs is kept constant during culture.Fig. 7. Representative images of SOFs and MOFs in hMI. White arrows, basal laminae. Bar = 50 µm
Discussion
During female sex differentiation, the germ-cell nest breakdown is a key early event in mammalian folliculogenesis, in which physiologically a single layer of pregranulosa cells surrounds each oocyte resulting in the formation of primordial follicles [1, 2]. However, the presence of MOFs in the ovary is relatively frequent in various species [4, 9, 11–16, 25], including Bos taurus taurus with an occurrence ranging from 0.3 to 5.4% of the total follicle population [17]. Our findings showed a high incidence of MOFs in one individual (15.8%) compared to the other three individuals in which MOF frequency is within the range reported in literature in this species (2.7%).
We are unable to provide a clear explanation for the unusually high occurrence of MOFs observed in this individual. Several studies across different species reported a higher frequency of MOFs in prepubertal subjects compared to postpubertal ones [12, 14, 33, 34]. Although we have previously analysed a considerable number of ovaries from Bos taurus taurus individuals aged 8 to 24 months—corresponding to pubertal or adult stages [29, 30, 35]—this represents, in our experience as well as to the best of our knowledge in the literature, the first documented case of such a high occurrence of MOFs in this species.
Several factors reportedly contribute to MOF origin, including genetic, chemical, and physical influences that may interfere with the breakdown of germ-cell nests. Foetal or neonatal exposure to environmental endocrine disruptors in various species [9, 35–38], along with altered gene expression in transgenic or knockout mice [6, 39, 40], and foetal irradiation in rats [41], have been reported to increase the frequency of MOFs. We cannot rule out the possibility that the hMI was exposed to unknown factors inducing this unusually high incidence of MOFs at the time of germinal nest breakdown.
Regardless of the underlying causes of the high frequency of MOFs in the hMI individual, this occurrence provided a valuable opportunity to assess their activation and growth in comparison with coexisting SOFs from the same individual.
The role and the functionality of MOFs in reproductive physiology remain a matter of debate. Several studies consider MOFs as abnormalities in terms of formation, activation, and progression through the stages of follicular development [5, 6, 39]. Conversely, although they occur at a very low frequency, MOFs could normally proceed through folliculogenesis as they have been observed from primordial to periovulatory stages in different species [4, 11, 17, 34]. In addition, the high incidence of MOFs in mouse lines selected for high fertility suggested a potential association between the MOF occurrence and increased reproductive capability [42]. A further direct evidence of MOF developmental competence is the retrieval of conjoined oocytes in ART patients, which have led to full-term births [20–24].
Herein, several findings differed between fresh and in vitro cultured tissue. In uncultured tissue of hMI and lMI, the proportions of primary MOFs significantly exceeded those of primary SOFs. This could suggest either that primordial MOFs have a higher activation ability, or that MOFs spend a longer time at the primary stage compared to their SOFs counterparts. In cultured tissues, there is a remarkable drop of primordial follicles and a concomitant increase of primary follicles as mechanical fragmentation of ovarian cortical tissue followed by in vitro culture is known to promote dormant follicle activation through the Hippo pathway disruption [43–45]. Although data showed that MOFs are capable to activate and progress through the early stages of folliculogenesis, the frequencies of primordial and primary MOFs were higher and lower compared to the corresponding SOF stages, respectively. This observation, contrary to the in vivo findings, may suggest a lower activation tendency of the MOFs following in vitro culture. Nevertheless, the presence of a comparable proportion of secondary MOFs and SOFs in the hMI individual indicates that MOFs retain a normal capacity for follicular progression. Given the low frequency of MOFs, the lack of secondary MOFs in lMI cultured tissues was not unexpected, as after dynamic culture, the yield of secondary follicles is usually limited to 15–20% of the total follicle population [29, 30]. This is consistent with studies suggesting that the rarity of MOFs at advanced follicular stages is primarily due to their already low incidence at the primordial stage [46].
A second aspect differing between the fresh and the cultured tissues was the number of oocytes in dormant versus growing MOFs. Specifically, only in fresh hMI tissue, the mean oocyte number in primary MOFs almost doubled the corresponding number in primordial MOFs (4.8 vs 2.4, P < 00.05). In agreement with other studies, this could suggest that primary MOFs could form postnatally due to the fusion of neighboring activated follicles, through the infiltrative ability of granulosa cells [5, 7]. However, we did not observe figures of granulosa cell infiltration or breakage of the follicle basal lamina in histological and confocal analysis. Conversely, in hMI cultured tissue, the mean MOF oocyte number was kept constant through the primordial, primary, and secondary stages (2.4, 2.5, 2.3). One of the main differences between the theory of prenatal MOFs formation as a result of “nest breakdown failure” and the postnatal formation according to the hypothesis of granulosa cells’ invasiveness concerns the number of MOFs found in the adult ovary. In fact, in the case of a prenatal formation, the MOF pool should remain unchanged or at most decrease in the adult ovary, whereas according to the postnatal hypothesis, a de novo formation of MOFs could occur. Our results in hMI and lMI demonstrate that the percentage of total MOFs remains unchanged after culture suggesting that de novo formation does not occur in vitro. We cannot rule out the possibility that unknown factors present only in the in vivo environment promote the postnatal formation of MOFs as a result of fusion of activated follicles.
The literature comparing the ability of MOFs to undergo folliculogenesis in vitro with that of SOFs is limited. Most studies have been focused on the formation, oocyte functionality, and morphological features of MOFs in vivo, rather than on a detailed functional comparison with SOFs in vitro. To our knowledge, this is the first study in which MOF development and viability were assessed, comparing them to SOFs within the same individual. Confocal analysis of follicle viability in fresh and cultured tissues indicates that MOFs did not have an increased tendency to undergo atresia compared to SOFs. However, MOFs containing at least one or more dead oocytes were observed in both histological and confocal images. The observation of apparently viable MOFs with one or a few dead oocytes could cause a progressive reduction of the oocyte number in more advanced stages of folliculogenesis [8, 10].
Several studies detected differences among the oocytes contained in advanced MOF stages in terms of size and meiotic condition [11]. We did not observe evident size differences among oocytes contained in MOFs from the primordial to the secondary stages. This could suggest that the reported asynchrony in growth and meiotic maturation among oocytes in single advanced MOF stages could arise later, during the advanced preantral or the antral phase of folliculogenesis.
In conclusion, the present findings indicate that MOFs do not substantially differ from SOFs in their ability to undergo early folliculogenesis in vitro. MOFs demonstrated the capacity to activate and progress through the primordial, primary, and secondary stages without evident signs of increased atresia or compromised viability. While certain differences emerged between fresh and cultured tissues—particularly regarding oocyte number dynamics and follicle activation potential—these did not appear to hinder MOF development. Importantly, the maintenance of a constant proportion of MOFs after culture supports the hypothesis that de novo MOF formation does not occur in vitro, reinforcing the idea of a predominantly prenatal origin. Further studies are needed to explore whether environmental or molecular factors might promote postnatal MOF formation in vivo and to determine the developmental competence of MOFs beyond the early stages. These findings offer new insight into the developmental plasticity of MOFs and support their potential relevance in reproductive physiology.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chaves MS, Azevedo HC, Luz VB, Ferreira-Silva JC, Barros I, Paiva SR, de Olivera Melo E, de Melo Magalhães Padilha D, de Figueirêdo Freitas VJ, Bartolomeu CC, Lemos Oliveira MA. Occurrence, morphology, and morphometry of follicles containing multiple oocytes in Fec GE mutant Santa Inês ewes. Anim Reprod Sci. 2021;226:106690. 10.1016/j.theriogenology.2019.06.033.10.1016/j.anireprosci.2021.10669033561808 · doi ↗ · pubmed ↗