Bridging the gaps: advanced techniques to unlock lipid function in plant reproductive development
Ze-Hua Guo, Mee-Len Chye

TL;DR
This paper explores how lipids contribute to plant reproduction and highlights advanced techniques to study their functions.
Contribution
The paper introduces advanced lipidomic techniques to better understand lipid roles in plant reproductive development.
Findings
Lipids regulated by specific genes are crucial during floral development.
ACBPs and related enzymes are key in fatty acid production and seed oil accumulation.
Advanced techniques like lipidomic studies and mass spectrometry imaging are essential for analyzing lipid functions.
Abstract
In plant cells, lipids serve various roles facilitating membrane bilayer formation, energy storage and signaling molecules. Acyl lipids are the most common in distinct plant cell compartments. Lipids regulated by key genes encoding fatty acid desaturases, diacylglycerol acyltransferase, 3-ketoacyl-CoA synthase and acyl-CoA-binding proteins (ACBPs) are deemed crucial during floral development. ACBPs, along with long-chain acyl-CoA synthase, acetyl-CoA carboxylase, fatty acid synthase, acyl-acyl carrier protein desaturases, acyl-ACP thioesterases and the ATP-binding cassette transporter subfamily A, contribute to fatty acid (FA) production, lipid transport and seed oil accumulation, making them bioengineering targets. To investigate lipid function, it is important to use appropriate analytical strategies because different lipid classes contain distinct FA patterns. These well-developed…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
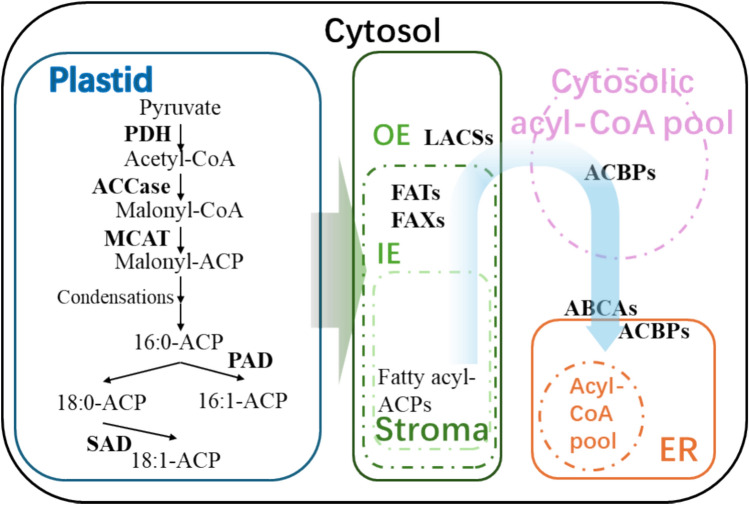 Figure 1
Figure 1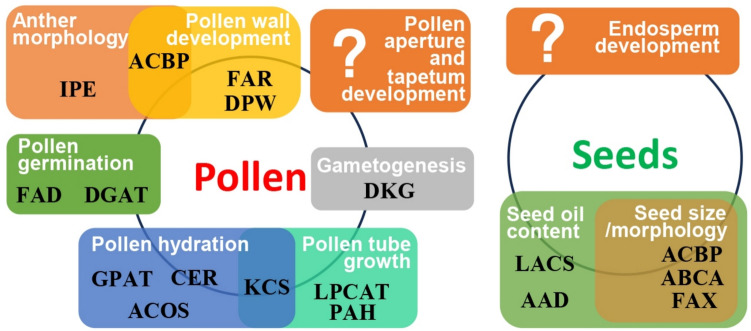 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLipid metabolism and biosynthesis · Plant Surface Properties and Treatments · Fatty Acid Research and Health
Introduction
Food security is a critical concern worldwide due to a growing population and an increased demand for sustainable agriculture (FAO 2024). In this context, research on plant reproduction plays a pivotal role in generating fruits and seeds, thereby directly impacting crop yield and quality. Some fruits and seeds produce oil, a vital nutrient as people worldwide consume over three quarters of their daily total lipid intake from plant-based oils to cover the essential fatty acids (FAs) that the human body cannot synthesize (Sumara et al. 2023). Also, plant oils support the production of pharmaceuticals and cosmetics (Munir et al. 2017; Raygan et al. 2019). In terms of biochemistry, these oils are extremely complex matrices because they contain chemical compounds that differ in concentration and origin (Sumara et al. 2023). For example, the discovery of bioactive compounds in oils has led to their applications for health benefits and industrial promotion of pressing oils from non-typical oil plants.
Besides its noteworthy role in agriculture, plant reproduction is important for both the persistence and propagation of plant species (Yadava et al. 2019). This stage in the life cycle of the plant is complex and the developmental processes determining seed yield are highly orchestrated (Pajoro et al. 2014). Tasked with a key role, lipid metabolism significantly affects the development and function of reproductive organs such as pollen, anther, stigma and seed (Fadhli Hamdan et al. 2021; Guo et al. 2022; Ortiz et al. 2020; Vallarino et al. 2023). Over the years, plant biologists have focused on these reproductive structures, employing various research techniques to gain a deeper perception of the underlying mechanisms (Borisjuk et al. 2023).
Through lipid biosynthesis, plants generate a diverse range of lipid species including phospholipids and triacylglycerol (TAG), to facilitate different functions (Ohlrogge and Browse 1995). The amphiphilic nature of phospholipids allows them to form fluid structures in cell membranes, serving as permeable barriers and maintaining membrane integrity (Li-Beisson et al. 2013; Ohlrogge and Browse 1995), as their composition is amenable under stress (Du et al. 2010; Oubohssaine et al. 2024). While various phospholipids maintain membrane fluidity and integrity, lipids within the cell membranes act as ligands to activate signal transduction and mediate signaling pathways (Eyster 2007), where lipid-derived second messengers affect cell metabolism and gene expression (Giusto et al. 2010; Newton et al. 2016). TAGs consist of three esterified FA molecules attached to a glycerol backbone, and represent the most abundant lipids in energy reserve during various biological processes (Li-Beisson et al. 2013), including seedling development (Graham 2008), pollen development (Zhang et al. 2009), pollen–stigma interaction (Wolters-Arts et al. 1998), pollen germination (Hernández et al. 2020; Zienkiewicz et al. 2013) and pollen tube elongation (Hernández et al. 2020). In Arabidopsis, TAG breakdown initiates with TAG lipase-mediated FA and glycerol release (Eastmond 2006), and the resultant free FAs (FFAs) are released into the peroxisome (Hayashi et al. 2002; Zolman et al. 2001) for β-oxidation (Graham 2008). Subsequently, FAs are converted into acetyl-CoA, and utilized for carbohydrate synthesis in seed germination and seedling development via the glyoxylate cycle and gluconeogenesis (Graham 2008).
Considering the importance of lipids, a broad range of methods and technologies have been developed to comprehend their function. Following lipid isolation from plants, various lipids including FAs can be derivatized to higher volatility, facilitating analysis by gas chromatography (GC) or GC-mass spectrometry (MS) (Brands et al. 2021). While direct infusion MS (shotgun lipidomics) and liquid chromatography-mass spectrometry (LC–MS) represent high-throughput lipid analyses (Gutbrod et al. 2021), imaging methods based on matrix-assisted laser desorption/ionization mass spectrometry imaging (MALDI-MS) techniques enable locating different lipids at the tissue level (Sturtevant et al. 2021). Also, investigations on lipid-protein interactions are indispensable in systems biology for assigning lipid function in a metabolic background (Guo et al. 2022).
Although the reproductive role of lipids have been previously reviewed in Wan et al. (2020), there is a need for an updated summary that incorporates recent findings and advancements in techniques. In this review, the lipid metabolic pathways in plant reproductive organs are discussed and recent techniques on lipid studies summarized to provide fresh insights..
Lipids in reproductive organs
Lipids function in pollen germination and pollination
For flowering plants, pollen lipids possess various functions including a crucial role during pollen germination. Various FA species including long-chain and very-long chain FAs, are abundant in pollen, and FA composition is known to vary among plants (Breygina et al. 2023; Zhang et al. 2016). Consisting of a mixture of 25 FAs including some omega acids, pollenkitt forms bridges between pollen grains but not between grains and stigma papillae, and performs a key role in pollen protection favoring pollination (Chichiriccò et al. 2019). Pollen development is accompanied by changes in the quantity of lipid metabolites (Rotsch et al. 2017), which are subject to gene regulation during lipid biosynthesis (Table 1). The coordinated action of olive fatty acid desaturases OeFAD2-3 and OeFAD3B led to increases in linoleic and alpha-linolenic acids as observed in germinating pollen, while the action of olive diacylglycerol acyltransferase1 (OeDGAT1) promoted TAG accumulation (Hernández et al. 2020). In Arabidopsis, 3-ketoacyl-CoA synthase4 (AtKCS4) plays a notable role in very-long-chain FA production as an atkcs4 mutation disrupted pollen tube elongation (Kim et al. 2021). In maize (Zea mays), aliphatic metabolism was greatly altered when the contents of lipid constituents especially C16/C18 FAs and their derivatives were significantly reduced in male-sterile mutant irregular pollen exine2 (ipe2), which displayed shrunken anthers and lacked starch accumulation in mature pollen grains (Huo et al. 2020). Two maize mutants (ms25-6065 and ms25-6057) disrupted in fatty acyl reductases (FARs) exhibited defective anther cuticles, abnormal Ubisch body formation, impaired pollen exine formation and total male sterility (Zhang et al. 2021a).Table 1. Genes related to lipid biosynthesis in flowers. The genes involved, the gene family to which each belongs, the organ of its expression, reported function, source and references are summarized in the following tableGenesGene familyOrganFunctionOrganismReferencesOeFAD2-3 and OeFAD3BFatty acid desaturasePollenLinoleic and alpha-linolenic acid biosynthesisOliveHernández et al. (2020)AtDGAT1Diacylglycerol acyltransferasePollenTAG biosynthesisArabidopsisHernández et al. (2020)AtKCS43-ketoacyl-CoA synthasePollenVery-long-chain FA biosynthesisArabidopsisKim et al. (2021)OsC6Lipid transfer proteinPollenExchanging lipids between membranesRiceZhang et al. (2010)ZmIPE2GDSL lipasePollenLipid breakdownmaizeHuo et al. (2020)ZmMs25Fatty acyl reductasePollenFA biosynthesismaizeZhang et al. (2021a)AtLPCATLysophosphatidylcholine acyltransferasePollenFA biosynthesisArabidopsisSong et al. (2024)AtPAHPhosphatidic acid phosphohydrolasePollenLipid breakdownArabidopsisSong et al. (2024)AtDGK2 and AtDGK4Diacylglycerol kinasePollenPhospholipid biosynthesisArabidopsisAngkawijaya et al. (2020)AtCER1, AtCER3, AtCER6 and AtCER10Enoyl-CoA reductaseStigmaLong-chain and very-long-chain FA biosynthesisArabidopsisWolters-Arts et al. (1998) and Zheng et al. (2005)AtKCS63-ketoacyl-CoA synthaseStigmaVery-long-chain FA biosynthesisArabidopsisQin et al. (2022)AtCER2-LIKEEnoyl-CoA reductaseAntherVery-long-chain FA biosynthesisArabidopsisQin et al. (2022)AtACBP3Acyl-CoA-binding proteinFloweracyl-lipid homeostasisArabidopsisGuo et al. (2024)AtACBP4, AtACBP5 and AtACBP6Acyl-CoA-binding proteinFlowerWax and cutin biosynthesisArabidopsisHsiao et al. (2015) and Ye et al. (2017b)OsGPAT3Glycerol-3-phosphate acyltransferaseAntherWax and cutin biosynthesisRiceMen et al. (2017)OsACOS12Acyl-CoA synthetaseAntherWax and cutin biosynthesisRiceYang et al. (2017)OsDPW2Acyl transferaseAntherWax and cutin biosynthesisRiceXu et al. (2017)ACBP, acyl-CoA-binding protein; ACOS, acyl-CoA synthetase; CER, enoyl-CoA reductase; DGAT, diacylglycerol acyltransferase; DGK, diacylglycerol kinase; DPW, defective pollen wall; FAD, fatty acid desaturase; GPAT, glycerol-3-phosphate acyltransferase; IPE, irregular pollen exine; KCS, 3-ketoacyl-CoA synthase; LPCAT, lysophosphatidylcholine acyltransferase; PAH, phosphatidic acid phosphohydrolase; At; Arabidopsis thaliana; Oe, Olea europaea; Os, Oryza sativa; Zm, Zea mays
Furthermore, lipids are essential for pollen tube penetration of the stigma and directing pollen-tube growth by regulating water flow to pollen in species with dry and wet stigmas (Wolters-Arts et al. 1998). Pollen tubes of Nicotiana tabacum (tobacco) can rapidly remodel their lipidome under heat stress likely by post-transcriptional and/or post-translational regulation (Krawczyk et al. 2022), while pollen sterol content is also subject to environmental conditions (Zu et al. 2021). The double mutant lpcat pah, defective in lysophosphatidylcholine acyltransferase (LPCAT) and phosphatidic acid phosphohydrolase (PAH), demonstrated decreased pollen tube growth in the pistil and reduced male transmission in Arabidopsis (Song et al. 2024). Lastly, two Arabidopsis diacylglycerol kinases, DGK2 and DGK4, were deemed crucial in gametogenesis and the biosynthesis of phosphatidylglycerol and phosphatidylinositol in the endoplasmic reticulum (ER) as the dgk2-1/– dgk4-1/– plants were gametophyte lethal (Angkawijaya et al. 2020). Overall, these findings highlight the interactive ability of pollen, the importance of lipids in the progamic phase, and their role in protecting pollen and promoting pollination.
Plant lipids in pollen development and fertilization
Fertilization, as a fundamental process in sexual reproduction of higher plants, relies on very-long-chain FAs (VLCFAs) and their derivatives for proper pollen hydration and germination (Batsale et al. 2021). While the mutation in the Arabidopsis gene encoding enoyl-CoA reductase, eceriferum10 (cer10), showed lower VLCFA content in shoots, disrupting shoot development and yielding shorter inflorescences and floral organ fusion (Zheng et al. 2005), other cer mutants atcer1, atcer3 and atcer6 displayed abnormal cuticles and male sterility under low-humidity conditions arising from impaired hydration on stigmas, thus highlighting the importance of lipids in plant reproduction (Wolters-Arts et al. 1998). Plant lipid transfer proteins (LTPs) are small and abundant lipid-binding proteins that have been demonstrated to facilitate lipid exchange across membranes (Zhang et al. 2019). OsC6, an LTP from Oryza sativa (rice), was shown to bind 1-pyrenedodecanoic acid, and osc6 mutant rice plants possessed defective orbicules and pollen exine accompanied by reduction in pollen fertility (Zhang et al. 2010).
Pollen coat lipids form a protective outer barrier and are essential for pollen-stigma interaction (Zhang et al. 2021b). In anthers, the expression of AtKCS6 and AtCER2-like genes in the anther endothelium was reported necessary for the production of ≥ C26 VLCFAs and derivatives which accumulate on mature pollen surfaces, serving as signaling molecules to activate water transfer from papilla cells for pollen hydration (Wattelet-Boyer et al. 2016). On the other hand, FA reduction during maturity is believed to discourage self-incompatible pollen mounting, because FA accumulation in stigmas was critical for compatible pollination (Qin et al. 2022). Interestingly, pollen coat VLCFA lipids, which are required for pollen hydration, were not reported to be strictly species-specific (Zhan et al. 2018).
Sporopollenin biosynthesis and lipid metabolism in pollen development
Sporopollenin is a highly durable and chemically resistant biopolymer that constitutes the outer walls of pollen grains, playing a crucial role in the protection of genetic material during pollination (Xiao et al. 2025). During sporopollenin biosynthesis, long-chain hydroxylated FAs (HFAs) represent the primary precursors (Ariizumi and Toriyama 2011) which are essential for Arabidopsis tapetal development (Ariizumi and Toriyama 2011; Quilichini et al. 2010). Oil bodies provide a rapid source for the biosynthesis of membrane lipids in mature pollen grains (Piffanelli et al. 1998) and their accumulation in the developing pollen grains initiates after the first pollen mitosis (Evans et al. 1991; Owen and Makaroff 1995; Piffanelli et al. 1997, 1998).
It has been reported that acyl-CoA-binding proteins (ACBPs) are important in lipid biosynthesis and transport during anther cuticle and pollen wall development (Hsiao et al. 2015; Ye et al. 2017b), implying their significance in pollen development (Shi et al. 2015). Null mutations of atacbp4 and atacbp5 contain more floral wax and cutin than the wild type (WT), while atacbp4 and atacbp4atacbp5 pollen exhibited a decrease in stearic acid but an increase in linoleic acid (Ye et al. 2017b). Pollen in the atacbp4atacbp5atacbp6 triple mutant was deformed, with fewer and smaller oil bodies (Hsiao et al. 2015; Ye et al. 2017b), suggesting that lipid metabolism was abrogated in the atacbp4atacbp5atacbp6 triple mutant. These studies showed AtACBP4, AtACBP5 and AtACBP6 play a combinatory role in pollen development, potentially affecting acyl-lipid transport during sporopollenin biosynthesis. Moreover, the recent findings on AtACBP3 conveyed its role in anther development and revealed the potential correlation between lipid content and anther morphology (Guo et al. 2024).
In rice, glycerol-3-phosphate acyltransferase3 (OsGPAT3) (Men et al. 2017) and acyl-CoA synthetase12 (OsACOS12) (Yang et al. 2017) were found to affect wax and cutin biosynthesis and were both required for anther development and male fertility. Also, rice DEFECTIVE POLLEN WALL2 (OsDPW2) encoding an acyl transferase was deemed necessary for rice pollen development, as its mutant (osdpw2) had increased amounts of cutin and waxes (Xu et al. 2017). While numerous functions of lipid-related genes in pollen have been reported (Fig. 2), there are more to be explored for potential roles in tapetum development (Yao et al. 2022) and pollen aperture formation (Zhao et al. 2023), which represent the less understood aspects in pollen development.
Fatty acid biosynthesis pathways in plant seed oils
Plant seed oils can confer beneficial health properties (Yang et al. 2018) given their significant phytochemical content including carotenoids, tocopherols, sterols, phenolic compounds, vitamins and minerals (Erkan et al. 2008). In plant seeds, de novo FA biosynthesis occurs in plastids (Fig. 1), which is catalyzed by the type II fatty acid synthase (FAS) with cooperation with other enzymes and enzymatic complexes such as the pyruvate dehydrogenase complex (PDC), acetyl-CoA carboxylase (ACCase) or the FAS complex (Troncoso-Ponce et al. 2016). At the first step of FA synthesis, ACCase transforms acetyl-CoA and bicarbonate into malonyl-CoA (Ohlrogge and Browse 1995). The malonyl group from CoA is then transferred to acyl carrier protein (ACP) by a malonyl-CoA: acyl carrier protein S-malonyltransferase (MCAT) to assemble FAs (Ohlrogge and Browse 1995). Through a series of condensation with acyl-CoA catalyzed by FAS enzymes, the malonyl-ACP is elongated to saturated FA 16:0-ACP (Ohlrogge and Browse 1995). The fate of 16:0-ACP is either desaturation to 16:1-ACP or elongation to 18:0-ACP, which eventually undergoes desaturation to form 18:1-ACP (Ohlrogge and Browse 1995).Fig. 1A diagram on de novo fatty acid biosynthesis in seeds. Pathways of lipid biosynthesis are indicated using arrows. The blue box is a representative plastid. The green box indicates a zoom-in view of the plastid stroma, while green boxes within (with dashed borders) represent the inner envelope (IE) and outer envelope (OE) of the stroma. The thick grey arrow indicates transfer of fatty acyl-ACPs to the stroma. The pink circle with dashed outline indicates the cytosolic acyl-CoA pool. The orange box represents the endoplasmic reticulum (ER), and orange circle within (with dashed border) denotes the acyl-CoA pool in the ER. ABCA, ATP-binding cassette transporter subfamily A; ACBP, acyl-CoA-binding protein; ACCase, acetyl-CoA carboxylase; ACP, acyl carrier protein; FAT, fatty acyl-ACP thioesterases; FAX, FATTY ACID EXPORT; LACS, long-chain acyl-CoA synthase; MCAT, malonyl-CoA:ACBP transacylase; PAD, palmitoyl-ACP desaturases; PDH, pyruvate dehydrogenase complex; SAD, stearoyl-ACP desaturase. Proteins including enzymes are bolded
Lipid biosynthesis in seeds
Many genes expressed in seeds have been reported to modulate lipid biosynthesis (Table 2). This process is primarily governed by the coordinated action of multiple enzymes in the plastidial FAS pathway, including ACCase which catalyzes the initial carboxylation step, and the Kennedy pathway which is responsible for triacylglycerol assembly in the endoplasmic reticulum (Ohlrogge and Browse 1995). For instance, the soluble acyl-acyl carrier protein desaturases (AADs) in stroma determines the proportion of newly-synthesized saturated versus monounsaturated fatty acids (Kachroo et al. 2007). There are seven genes encoding AADs in the Arabidopsis genome including FATTY ACID BIOSYNTHESIS2 (AtFAB2) and SUPPRESSOR OF SALICYLIC ACID INSENSITIVE2 (AtSSI2) (Kachroo et al. 2007). Depending on their regio- and substrate-specificity, AADs are characterized as stearoyl-ACP desaturases (SADs) and palmitoyl-ACP desaturases (PADs) (Kachroo et al. 2007). While SADs are responsible for desaturation of 18:0-ACP to 18:1-ACP, PADs catalyze the transition of 16:0-ACP to 16:1-ACP (Kachroo et al. 2007). As the single Arabidopsis aad mutation barely affected seed FA composition, it appears that the functions of AADs/SADs are redundant (Kachroo et al. 2007). This is further supported by recent studies on four SAD desaturases (AtFAB2, AtAAD1, AtAAD5 and AtAAD6) which were shown to play collaborative roles in oleic acid biosynthesis in developing Arabidopsis seeds (Kazaz et al. 2020).Table 2. Key gene families associated with lipid biosynthesis in seeds. The genes involved, the gene family to which each belongs, the organ of its expression, reported function, source and references are summarized in the following tableGenesGene familyFunctionOrganismReferenceAtFAB2 and* AtSSI2Acyl-acyl carrier protein desaturaseExchanging saturated and unsaturated FAsArabidopsisKachroo et al. (2007)AtAAD1, AtAAD5 and AtADD6Acyl-acyl carrier protein desaturaseOleic acid biosynthesisArabidopsisKazaz et al. (2020)AtFATAcyl-ACP thioesteraseBiosynthesis of saturated FAsArabidopsisBonaventure et al. (2003)CsFATAcyl-ACP thioesteraseBiosynthesis of saturated FAsC. sativaOzseyhan et al. (2018)AtLACS1, AtLACS4 and AtLACS9Long-chain acyl-CoA synthaseTAG biosynthesisArabidopsisZhao et al. (2010) and Jessen et al. (2015)BnLACS2Long-chain acyl-CoA synthaseTAG biosynthesisB. napusDing et al. (2020)GmDGATGlycerol-3-phosphate acyltransferaseTAG biosynthesisGlycine maxZhao et al. (2019)AtACBP1Acyl-CoA-binding proteinphytosterol biosynthesisArabidopsisLung et al. (2017 and 2018)BnACBP2Acyl-CoA-binding proteinTAG biosynthesisB. napusLiao et al. (2019)OsACBP2Acyl-CoA-binding proteinLong-chain FA accumulationRiceGuo et al. (2019b)VfACBP3A and VfACBP3BAcyl-CoA-binding proteinFA biosynthesisTung treePastor et al. (2013)EgACBP1 and EgACBP3Acyl-CoA-binding proteinEndosperm oil accumulationOil palmAmiruddin et al. (2020)AtABCA9ATP-binding cassette transporter subfamily ATransportation of FAs into the ERArabidopsisKim et al. (2013)AtABCA9ATP-binding cassette transporter subfamily ASeed oil productionC. sativa*Cai et al. (2021)AAD, acyl-acyl carrier protein desaturase; ABCA, ATP-binding cassette transporter subfamily A; ACBP, acyl-CoA-binding protein; FAB2, FATTY ACID BIOSYNTHESIS2 (FAB2); FAT, fatty acyl-ACP thioesterase; FAX, fatty acid export; LACS, long-chain acyl-CoA synthase; SSI2, SUPPRESSOR OF SALICYLIC ACID INSENSITIVE2; At; Arabidopsis thaliana; Bn, Brassica napus; Cs, Camelina sativa; Eg, Elaeis guineensis; Os, Oryza sativa; Vf, Vernicia fordii; Zm, Zea mays
Fatty acid biosynthesis and transportation in seeds
While less than 40% of FAs are trafficked into the plastidic prokaryotic pathway, the majority of de novo synthesized FAs need to be transported to the ER for lipid synthesis via the eukaryotic pathway (Li et al. 2016). Firstly, acyl-ACPs are hydrolyzed by fatty acyl-ACP thioesterases (FAT) at the inner plastid envelope membrane (IE) to FFAs. In Arabidopsis (Bonaventure et al. 2003) and Camelina sativa oilseeds (Ozseyhan et al. 2018), it was reported that the loss of function in FAT proteins led to a reduction in saturated FAs. The FATTY ACID EXPORT (FAX) proteins are also FFA transporters anchored in the IE (Li et al. 2015). In Arabidopsis, the atfax1 mutant possessed lower leaf and floral TAG content (Li et al. 2015), while atfax2atfax4 seeds contain 30% less TAGs than the WT (Li et al. 2020), providing evidence that loss of AtFAX2 and AtFAX4 function affected FFA transport and TAG biosynthesis during seed development. Following the transportation by FAX, FFAs are mobilized across the plastid outer envelope (OE) via vectorial acylation by long-chain acyl-CoA synthase (LACS), which incorporates FFAs into acyl-CoA thioesters (Fig. 1). In Arabidopsis, nine AtLACS proteins are known to function in lipid biosynthesis. Double mutants of Arabidopsis atlacs1atlacs9 and atlacs4atlacs9 showed decreased FA content in seed TAGs (Jessen et al. 2015; Zhao et al. 2010), while the overexpression of Brassica napus BnLACS2 in rapeseed resulted in higher seed oil content (Ding et al. 2020).
Seed acyl-lipid homeostasis and oil accumulation
FFAs are likely transported to the ER by ACBPs (Fig. 1). In Arabidopsis, three cytosolic ACBPs (AtACBP4, AtACBP5 and AtACBP6) have overlapping roles in seed acyl-lipid homeostasis (Hsiao et al. 2014). The single and double mutants of these cytosolic AtACBPs showed altered seed oil composition in comparison to the WT affecting FA and lysophosphatidylcholine (LPC) composition, implying their combinatory function in seed development (Guo et al. 2019a). AtACBP1 negatively affects sterol synthesis, as demonstrated by an increase in total phytosterols in atacbp1 mutant seeds (Lung et al. 2017, 2018). AtACBP1 has also been suggested to transport membrane-associated acyl-CoAs from the ER to the plasma membrane (Raboanatahiry et al. 2018). In Brassica, the high expression of BnACBP2 during rapid seed oil accumulation implies its potential role in TAG biosynthesis (Liao et al. 2019), and the Class I BnACBP not only modulates C18:1-CoA content but also influences TAG content (Yurchenko et al. 2014). In rice, Class I OsACBP2 is highly expressed in seed embryos, the scutellum and the aleurone layer, with OsACBP2 overexpression in transgenic rice causing accumulation of long-chain FAs in mature embryos, and enhanced grain size and bran oil content (Guo et al. 2019b). In Vernicia fordii (tung tree), both VfACBP3A and VfACBP3B were reported to be expressed throughout seed development affecting seed FA metabolism (Pastor et al. 2013). In Elaeis guineensis (oil palm), EgACBP1 and EgACBP3, also displayed marked increased expression during the active period of endosperm oil accumulation (Amiruddin et al. 2020).
Besides ACBPs, the Arabidopsis ATP-binding cassette transporter subfamily A (ABCA) member, AtABCA9, mediates FA transport into the ER (Kim et al. 2013). AtABCA9 was demonstrated to be ER-localized and was highly expressed during seed development (Kim et al. 2013). The atabca9 mutant seeds were smaller, deformed and contain lower amount of TAGs (Kim et al. 2013). In contrast, TAGs had accumulated in transgenic Arabidopsis AtABCA9-overexpressing seeds (Kim et al. 2013). As the developing atabca9 seeds incorporated less ^14^C-oleoyl-CoA into TAG than the WT seeds (Kim et al. 2013), AtABCA9 was proposed to transport acyl-CoAs into the ER to provide substrates for TAG synthesis. Also, the overexpression of Arabidopsis AtFAX1 and ABCA9 promoted oil production in C. sativa seeds (Cai et al. 2021). Given that the overexpression of lipid-transporting proteins frequently impacted seed oil content, this strategy can be applied to bioengineer and boost seed oil accumulation in transgenic plants. Evolutionary studies have shown that genes involved in seed germination and lipid metabolism undergo strong positive selection, suggesting a potential reservoir of candidate genes for enhancing seed oil content and germination through targeted directional selection (Parakkunnel et al. 2024). The versatility of lipid-related genes suggests that additional functions of these genes have yet to be addressed in other tissues, for instance in seed endosperm (Fig. 2).Fig. 2A summary of the proteins in lipid metabolism within reproductive organs. Protein functions in pollen and seeds are depicted. Proteins including enzymes are bolded in black. White letters refer to various events in plant reproduction. The circle on the right indicates biological events in seeds, while that on the left, pollen. The white question marks identify stages for future work on mapping proteins in lipid metabolism to specific functions. AAD, acyl-acyl carrier protein desaturase; ABCA, ATP-binding cassette transporter subfamily A; ACBP, acyl-CoA-binding protein; ACOS, acyl-CoA synthetase; CER, enoyl-CoA reductase; DGAT, diacylglycerol acyltransferase; DGK, diacylglycerol kinases; DPW, defective pollen wall; FAD, fatty acid desaturase; FAR, fatty acyl reductases; FAX, fatty acid export; GPAT, glycerol-3-phosphate acyltransferase; IPE, irregular pollen exine; KCS, 3-ketoacyl-CoA synthase; LACS, long-chain acyl-CoA synthase; LPCAT, lysophosphatidylcholine acyltransferase; PAH, phosphatidic acid phosphohydrolase
Building on the evolutionary insights and tissue-specific potential of lipid-related genes as highlighted in Fig. 2, the next sections explore advanced methodologies to dissect lipid-protein interactions, metabolic pathways, and spatial lipid dynamics. These techniques bridge evolutionary biology with molecular precision: isothermal titration calorimetry (ITC) quantifies lipid-protein binding affinities, X-ray crystallography resolves structural interactions of acyl-CoA-binding proteins (ACBPs) with thioesters, and lipidomic analyses—including optimized extraction protocols and gas chromatography—map lipid composition and flux. Further, enzyme assays and chromatography-mass spectrometry platforms decode acyl-CoA thioester metabolism, while matrix-assisted laser desorption/ionization mass spectrometry imaging (MALDI-MSI) spatially profiles lipids in tissues like the endosperm. Collectively, these approaches illustrate the technology available for enhancing seed germination efficiency and seed oil accumulation, and for translating evolutionary signatures into strategies in bioengineer oil crops in sustainable lipid production.
Techniques in studying lipids
Isothermal titration calorimetry offers a robust approach in lipid-protein interaction analysis
To investigate the many functions of newly characterized lipid-related genes in reproduction, several methodologies have gained popularity. In investigations involving the interaction of lipids with proteins, researchers typically employ binding assays. However, traditional methods such as the Lipidex assays (Glatz and Veerkamp 1983) and filter-binding assays (Dowler et al. 2002) have limitations in terms of both efficiency and data accuracy, rendering them more suitable for qualitative analysis of lipid-protein interactions. Isothermal titration calorimetry (ITC) that can measure heat changes generated by binding and record the energetics associated with reactions or processes occurring at a constant temperature (Wiseman et al. 1989), is deemed more appropriate for studying biochemical and molecular interactions (Freire et al. 1990; Jiang et al. 2019). ITC provides information on the thermodynamics of a variety of biophysical process including reactions catalyzed by enzymes and those involving ligand binding that occur between macromolecules, as well as reactions involving ligand- or pH-induced macromolecular conformational changes (Freire et al. 1990). Very early on in the 1970s, ITC was utilized to investigate binding of nucleotides to proteins, antibody to antigen, ligands to human hemoglobin, toxins to proteins as well as peptide-lipid association (Freire et al. 1990), making it fitting for studying protein-lipid binding.
Researchers have employed ITC to analyze the affinity between acyl-CoA thioesters and ACBPs in Arabidopsis (Hsiao et al. 2014; Hu et al. 2018; Xue et al. 2014), rice (Guo and Chye 2021; Guo et al. 2017, 2021) and soybean (Lung et al. 2022). This method has also been applied to address protein–protein interactions between ACBPs and their protein partners (Miao et al. 2019; Ye et al. 2017a). Although traditional binding assays such as the Lipidex assays (Rasmussen et al. 1990) and filter-binding assays (Stevenson et al. 1998) were utilized to study binding involving ACBPs earlier (Chye 1998; Chye et al. 2000; Gao et al. 2010, 2009; Leung et al. 2006; Li et al. 2008; Meng et al. 2011; Xiao and Chye 2011), ITC offers more comprehensive and reliable information by documenting the thermodynamics of binding parameters (Guo et al. 2017, 2021; Hsiao et al. 2014; Hu et al. 2018; Lung et al. 2022; Miao et al. 2019; Xue et al. 2014; Ye et al. 2017a). ITC allows for the accurate determination of thermodynamic parameters, such as changes in Gibbs free energy (ΔG), enthalpy (ΔH) and entropy (ΔS), by using curve-fitting parameters from a selected binding site model (Ladbury et al. 2010). The ΔG value indicates the affinity of binding between the ligand and the protein, while the ΔH value represents the heat energy changes that occur during complex formation at a constant temperature (Ladbury et al. 2010). Furthermore, the equilibrium binding constant (K) can be calculated based on the amount of free or bound ligand at any point during the titration process (Ladbury et al. 2010). These parameters enable the comparison of binding affinities among different proteins and ligands, providing biochemical evidence to support biological functional studies.
X-ray crystallography: Unveiling ACBP and acyl-CoA thioester interactions
X-ray crystallography has also been used to study protein–ligand interactions, such as those between ACBPs and acyl-CoA thioesters. Using this technique, the acyl-CoA-binding (ACB) domain was shown to be most structurally conserved (Burton et al. 2005), featuring four alpha helices in an up-down-down-up arrangement (Andersen and Poulsen 1993). This unique helical scaffold is preserved across ACBPs from various organisms, including yeast (Teilum et al. 2005), man (Taskinen et al. 2007), armadillo (Costabel et al. 2006), Plasmodium falciparum (van Aalten et al. 2001), the plant fungus Moniliophthora perniciosa (Monzani et al. 2010) and rice (Guo et al. 2017; Jin et al. 2020).
Moreover, ACBPs have been co-crystallized to demonstrate their interaction with acyl-CoA thioesters (Jin et al. 2020; Kragelund et al. 1993; Monzani et al. 2010; Taskinen et al. 2007). The liganded ACBP modes exhibit diversity: bovine ACBP binding C16:0-CoA esters in a 1 to 1 ratio (Kragelund et al. 1993), human liver ACBPs undergo dimerization when interacting with C14:0-CoA, and M. perniciosa ACBPs form dimers when binding C12:0-, C14:0-, C16:0-, C18:0- and C20:0-CoAs (Monzani et al. 2010; Taskinen et al. 2007). Rice OsACBP2 also forms a dimer when binding to its ligand C18:3-CoA, but the spatial conformation differed from human liver ACBPs and the dimerization is suggested to be interchangeable from monomer from size exclusion chromatography-coupled multiangle light scattering analysis (Jin et al. 2020). In summary, x-ray crystallography offers a more detailed perspective on protein-lipid interactions, paving the way for a new field in protein-lipid research.
Lipidomic analysis: Extraction methods, derivatization for gas chromatography, and advanced mass spectrometry workflow
Lipidomic studies refer to analyses of the lipidome of a tissue, organelle or cell. Their complete profile of lipid species can be accurately derived by modern multi-dimensional LC–MS-based lipidomics (Han 2016). Identifying the lipid alterations can reveal the cellular homeostasis (Harayama and Riezman 2018). The extraction of lipids from tissues is a crucial step for successful lipidomic analysis. The Folch method (Folch et al. 1957) and Bligh and Dyer method (Bligh and Dyer 1959) are classical techniques for lipid extraction, with the former predominantly used for solid tissue and the latter more effective for biological fluids (Pati et al. 2016). Both methods utilize chloroform/methanol in extracting major lipid classes, and lipid extraction from plants mostly rely on the Bligh and Dyer method (Saini et al. 2021). In vegetative tissue, phospholipases can quickly change lipid content, thus it is critical to inactivate this potential lipase activity to ensure successful extraction of plant lipids (Bengtsson et al. 2021).
After derivatization, lipid extracts are analyzed by GC to separate and identify individual compounds in mixtures using a selected column phase based on analyte affinity (Brands et al. 2021; McNair et al. 2019). Recorded by a detector utilizing flame ionization or mass spectrometric methods, the detected data from the column can reveal the composition of the flow-through gas and retention time, defining individual compounds (Bartle and Myers 2002). Absolute quantification of analytes is accomplished by incorporating defined internal standards, aiding in the determination of compound abundance (Brands et al. 2021). Recently, there have been significant advancements in MS techniques, particularly in the areas of data acquisition (Yuan et al. 2024) and MS imaging (Dong and Aharoni 2022; Horn and Chapman 2024; Ma et al. 2023; Yin et al. 2023). These innovations have promoted the application of MS to an even wider range (Yin et al., 2024).
Acyl-CoA thioester analysis: Enzyme assays, chromatography and mass spectrometry techniques
Acyl-CoA thioesters play essential roles in energy metabolism, the biosynthesis and recycling of complex lipids, posttranslational modification of proteins and the regulation of gene expression (Haslam and Larson 2021). The central role of acyl-CoA thioesters in cellular metabolism is widely known, and many techniques have been developed for the measurement of acyl-CoA content (Haslam and Larson 2021). Analyses are usually carried out by using either enzyme assays of Coenzyme A from hydrolyzed acyl-CoA thioesters, or chromatographic assays of released FAs, or the separation and UV detection of acyl-CoA thioesters by reversed-phase high-performance liquid chromatography (HPLC) (Haslam and Larson 2021).
Specifically for HPLC, the use of solid-phase extraction has improved recovery rates (Golovko and Murphy 2004) and hydrophilic interaction liquid chromatography has been utilized to reduce peak tailing (Abrankó et al. 2018). Moreover, a matrix assisted laser desorption ionization/time-of-flight (MALDI/TOF) mass spectrometer has been reported for complete structural identification of individual acyl-CoAs, including the location of unsaturated bonds in the FA chains (Wang and Hsu 2020). The method developed by Haslam and Larson (2021) represents an invaluable contribution, as it enables the measurement of acyl-CoA content from minute amounts of tissue. This advancement expands the scope of studies on acyl-CoA metabolism to include smaller organs, such as anthers and stigmas, which were previously difficult to analyze due to the limited tissue sample size available. Recent studies in Leishmania infantum (Carnielli et al. 2022), Brassica (Zhu et al. 2023) and mice (Talamonti et al. 2020) have employed this method to analyze lipid content and composition.
Matrix-assisted laser desorption/ionization mass spectrometry imaging (MALDI-MSI)
While lipid profiling usually takes homogenized samples, MALDI-MSI can demonstrate spatial localization of lipid molecules, visualizing the unequal or heterogeneously distribution of lipid molecules in different tissues and cell types (Chen et al. 2024b; Sturtevant et al. 2021). In various types of plant tissues, MALDI-MSI has been reported to analyze seed lipid distribution (Lu et al. 2018; Marmon et al. 2017; Sturtevant et al. 2017a), and this technique has been applied for analysis of seeds from Arabidopsis (Sturtevant et al. 2017a), Gossypium barbadense (Sturtevant et al. 2017b), B. napus (Woodfield et al. 2017), C. sativa (Usher et al. 2017), sorghum (Montini et al. 2020) and peanut (Wang et al. 2022). In a standard MALDI-MSI experiment, an intact tissue or tissue section is coated with a chemical matrix, and then subject to the mass spectrometer for a laser beam to pulse over the tissue (Sturtevant et al. 2021). A plume of ions is generated and then examined for their physical properties, resulting in data on physical distribution of ions within a sample (Sturtevant et al. 2021).
The process of preparing plant tissues for MALDI-MSI experiments follows a similar protocol to mammalian tissues (Sturtevant et al. 2021). Despite extra challenges in handling plant tissues, careful handling and preparation can successfully address most technical issues during MALDI-MSI analysis (Sturtevant et al. 2021). Different computing platforms such as MassImage, pyBASIS, and M2aia have enhanced clustering outcomes (Cordes et al. 2021), allowing visualization of sample relationships in MSI research (Zamora Obando et al. 2021). The novel matrix 2,4-dihydroxy-5-nitrobenzoic acid (DHNBA) overcomes previous limitations imposed by MALDI-MSI in plant phytohormone studies such as low hormone abundance and suboptimal matrix performance. DHNBA provides enhanced sensitivity, uniform deposition, and minimal background interference, enabling simultaneous detection, imaging, and precise tracking of multiple hormones (e.g., cytokinins, ABA, IAA, ACC) across diverse tissues to elucidate biosynthesis pathways and distinct functions (Chen et al. 2024a).
Conclusion and future perspectives
In summary, lipids play a variety of roles in plant cells, including in the formation of the membrane bilayer and acting as energy storage besides signaling molecules (Ohlrogge and Browse 1995). Acyl lipids are the most prevalent group of lipids, representing a wide range of structurally diverse compounds (Li-Beisson et al. 2013; Ohlrogge and Browse 1995). Different lipid classes are present in distinct compartments of plant cells, with each plant tissue displaying a unique lipid profile (Breygina et al. 2023; Erkan et al. 2008).
Lipids play a vital role in floral development, which is regulated by key genes associated with lipid function in flowers include fatty acid desaturases AtFAD2-3, AtFAD3B, AtDGAT1 (Hernández et al. 2020), AtKCS4 (Kim et al. 2021), AtKCS6 and AtCER2-like genes (Qin et al. 2022), as well as ACBPs (Hsiao et al. 2015; Shi et al. 2015; Ye et al. 2017b). Meanwhile, ACBPs, together with LACS (Guo et al. 2022; Jessen et al. 2015; Zhao et al. 2010), ACCase, FAS, AADs (Kachroo et al. 2007; Kazaz et al. 2020; Troncoso-Ponce et al. 2016), FAT, FAX (Bonaventure et al. 2003; Li et al. 2016; Ozseyhan et al. 2018) and ABCA (Kim et al. 2013), play crucial roles in FA production, lipid transport and seed oil accumulation, making them potential targets for bioengineering strategies to increase or modify seed oil content (Cai et al. 2021).
The amount of lipids within the cell can vary depending on their functions, and their quantity can range from highly abundant structural and storage lipids to smaller amounts for signaling lipids (Saini et al. 2021). Numerous molecular species with distinct FA patterns make up various lipid classes, including membrane glycerolipids and TAGs (Ohlrogge and Browse 1995). The abundance of these molecular species within a lipid class is an important factor when determining the analytical strategy for their examination (Saini et al. 2021). These strategies involve advanced lipidomic studies using modern multi-dimensional LC–MS techniques (Han 2016) as well as MALDI-MSI which allows for the spatial localization of lipid molecules (Sturtevant et al. 2021). Additionally, lipid-protein interactions can be investigated through lipid-binding assays and x-ray crystallography techniques (Guo et al. 2022). Given the importance of lipid function, scientists have utilized well-established methods such as strong anion exchange (SAX)-HPLC (Laha et al. 2021) and nuclear magnetic resonance (NMR) spectroscopy (Knaack et al. 2021). With the evolution of these techniques, the functions of lipids are anticipated to be rapidly elucidated.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1FAO (2024) FAO Cereal Supply and Demand Brief. https://www.fao.org/worldfoodsituation/csdb/en