“Expanding the Lactococcal Cell Wall Polysaccharide Paradigm: Novel Structures and Metabolic Pathways in the Emerging Dairy Species Pseudolactococcus laudensis and Pseudolactococcus raffinolactis”
Axel Soto‐Serrano, Irina Sadovskaya, Evgeny Vinogradov, Wenwen Li, Jun‐Hyeok Yu, Kelsey White, Douwe van Sinderen, Lukasz Krych, Paulina Deptula, Jennifer Mahony

TL;DR
This study explores new polysaccharide structures and biosynthesis pathways in two dairy-related bacteria species, expanding knowledge of their role in dairy fermentation.
Contribution
The study identifies novel cell wall polysaccharide genotypes and structures in Pseudolactococcus species, revealing new biosynthetic pathways.
Findings
Eight novel CWPS genotypes (E–L) were identified, expanding diversity in dairy-associated (pseudo)lactococci.
Chemical analysis revealed unprecedented CWPS structures, including a homopolysaccharide and a novel 6-deoxy-α-l-talan polysaccharide.
EPS loci were found in 25 of 28 strains, showing evidence of horizontal gene transfer and genomic mobility.
Abstract
Cell surface‐associated polysaccharides, including cell wall polysaccharides (CWPSs), capsular polysaccharides (CPSs), and exopolysaccharides (EPSs), play vital roles in bacterial interactions with their environment, influencing critical aspects of dairy fermentations, such as phage–host dynamics. Pseudolactococcus laudensis and Pseudolactococcus raffinolactis (formerly Lactococcus laudensis and Lactococcus raffinolactis) are emerging dairy‐associated species whose CWPSs remain uncharacterized. This study analyzed the complete genomes of 21 P. laudensis and seven P. raffinolactis strains to investigate the genetic diversity underlying CWPS and EPS production. Eight novel cwps genotypes (E–L) were identified, significantly expanding the known diversity within the dairy‐associated (pseudo)lactococci. Notably, E and G genotypes diverge from the classical rhamnan‐PSP organization,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
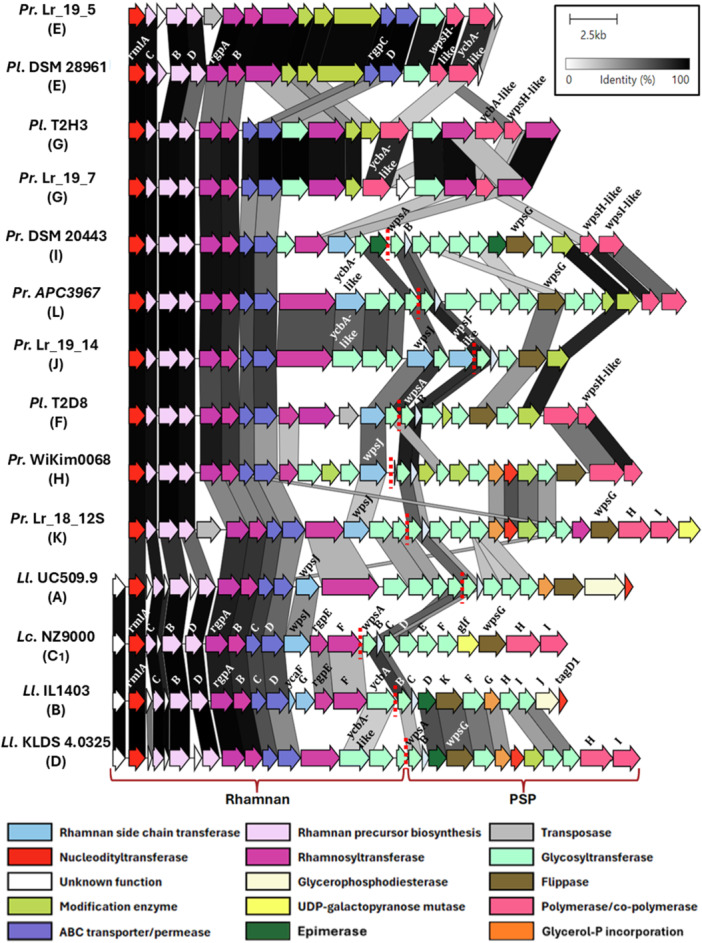 Figure 1
Figure 1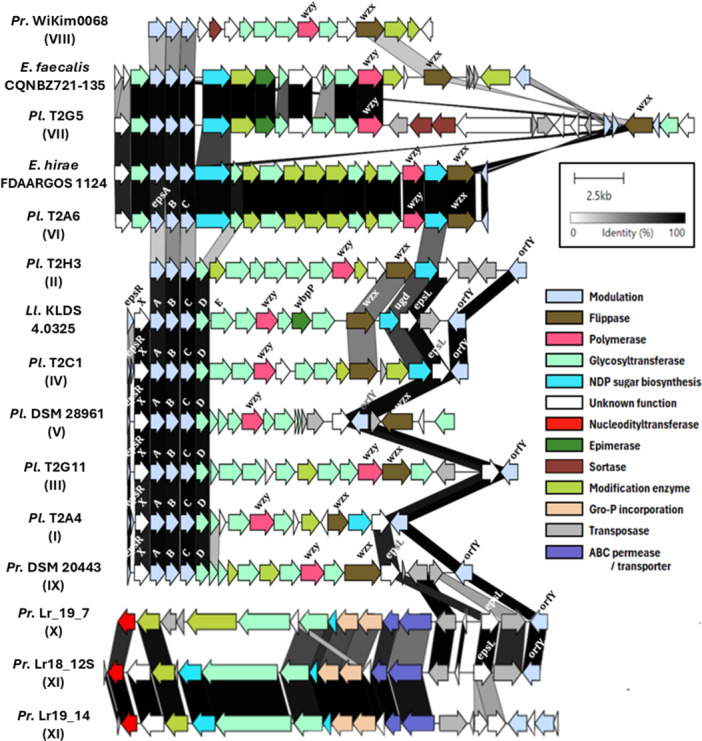 Figure 2
Figure 2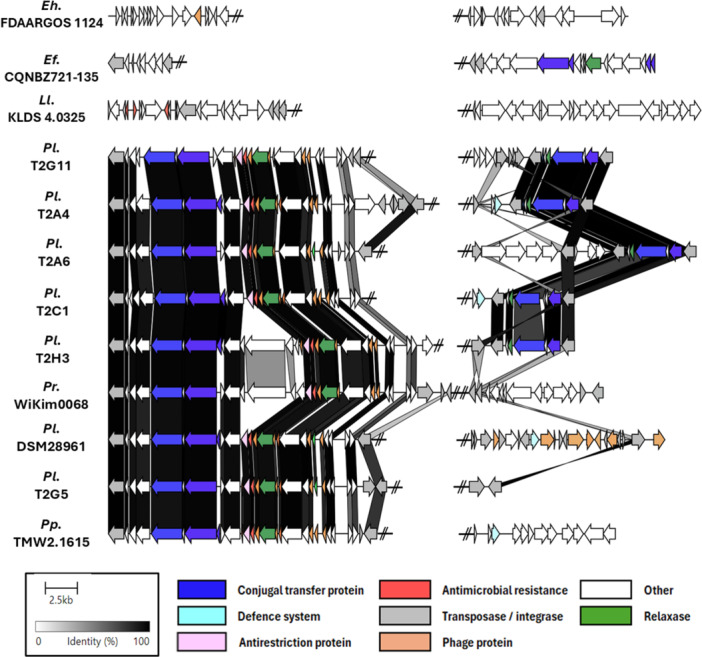 Figure 3
Figure 3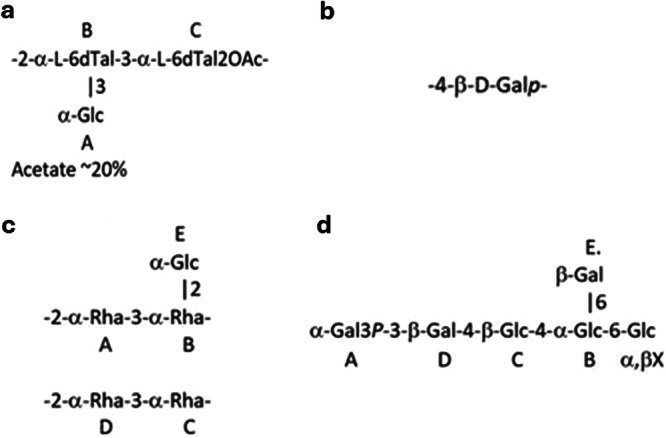 Figure 4
Figure 4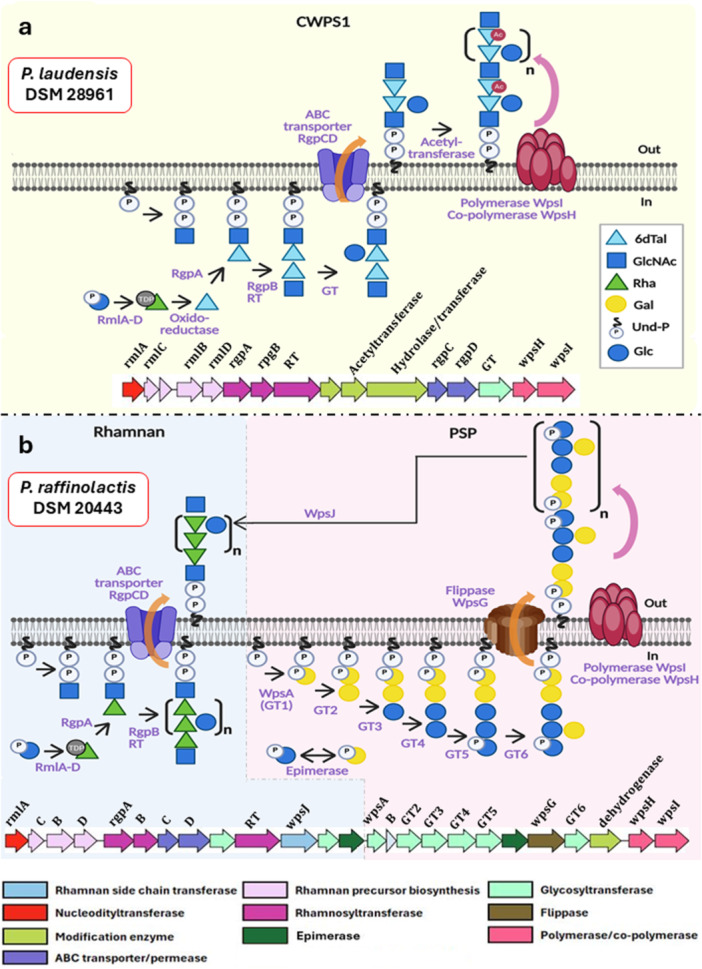 Figure 5
Figure 5| CWPS | EPS | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Species | Strain |
| Cluster start (bp) | Cluster end (bp) | Cluster length (bp) |
| Cluster start (bp) | Cluster end (bp) | Cluster length (bp) |
|
| DSM28961 | E | 182887 | 201332 | 18445 | V | 74320 | 90617 | 16297 |
| T2A4 | F | 1999196 | 2023901 | 24705 | I | 79289 | 93569 | 14280 | |
| T2A6 | F | 1946019 | 1970724 | 24705 | VI | 75286 | 94233 | 18947 | |
| T2A7 | F | 2085468 | 2110175 | 24707 | I | 107107 | 121389 | 14282 | |
| T2A8 | F | 2017410 | 2042116 | 24706 | I | 79288 | 93568 | 14280 | |
| T2C1 | F | 2022619 | 2047325 | 24706 | IV | 74431 | 91778 | 17347 | |
| T2C6 | F | 1944846 | 1969552 | 24706 | VI | 75286 | 94233 | 18947 | |
| T2C9 | F | 2042130 | 2066835 | 24705 | I | 79288 | 93568 | 14280 | |
| T2D8 | F | 2054139 | 2078844 | 24705 | I | 79288 | 93568 | 14280 | |
| T2E11 | F | 150314 | 175019 | 24705 | — | — | — | — | |
| T2E12 | F | 1944854 | 1969559 | 24705 | VI | 75286 | 94233 | 18947 | |
| T2E8 | F | 2054342 | 2079047 | 24705 | I | 79289 | 93569 | 14280 | |
| T2F10 | F | 1944795 | 1969499 | 24704 | VI | 75286 | 94233 | 18947 | |
| T2F2 | F | 1944847 | 1969552 | 24705 | VI | 75286 | 94233 | 18947 | |
| T2F8 | F | 2052966 | 2077672 | 24706 | I | 79288 | 93568 | 14280 | |
| T2G11 | F | 2048104 | 2072809 | 24705 | III | 74437 | 90237 | 15800 | |
| T2G3 | F | 2020537 | 2045242 | 24705 | IV | 74430 | 91777 | 17347 | |
| T2G5 | F | 2104178 | 2117025 | 12847 | VII | 76449 | 90201 | 13752 | |
| 295724 | 307558 | 11834 | |||||||
| T2H1 | F | 1999389 | 2024094 | 24705 | VI | 75287 | 94234 | 18947 | |
| T2H3 | G | 2031233 | 2054047 | 22814 | II | 78886 | 98046 | 19160 | |
| T2H4 | F | 2104194 | 2117041 | 12847 | VII | 76449 | 90202 | 13753 | |
| 295725 | 307559 | 11834 | |||||||
|
| WiKim0068 | H | 183549 | 210693 | 27144 | VIII | 80195 | 94639 | 14444 |
| DSM 20443 | I | 167655 | 193821 | 26166 | IX | 61407 | 78976 | 17569 | |
| Lr_19_7 | G | 231981 | 253318 | 21337 | X | 102387 | 125579 | 23192 | |
| Lr_19_5 | E | 374429 | 394173 | 19744 | — | — | — | — | |
| Lr_19_14 | J | 173936 | 197235 | 23299 | XI | 1151222 | 1173701 | 22479 | |
| Lr_18_12S | K | 168009 | 198226 | 30217 | XI | 1851815 | 1874084 | 22269 | |
| APC3967 | L | 152269 | 181745 | 29476 | — | — | — | — | |
- —This research was supported by the Milk Levy Fund and co‐financed by Arla Foods as part of the project “Cassandra: Quality Modelling Using Detailed Genomics.” J.M. is the recipient of a Frontiers of t
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Gut microbiota and health · Infant Nutrition and Health
Introduction
1
Bacteria produce a range of intracellular and extracellular glycoconjugates, often exhibiting significant structural and functional diversity (Tytgat and Lebeer 2014). Among these, cell surface‐associated polysaccharides play critical roles in bacterial interactions with their environment. These polysaccharides are generally categorized into three types: exopolysaccharides (EPSs), which are loosely attached and released into the environment; capsular polysaccharides (CPSs), which are bound to the cell, forming a protective barrier; and cell wall polysaccharides (CWPSs), which may or may not be covalently linked to the cell wall but do not form a capsule (Chapot‐Chartier 2014). Among the industrially significant species Lactococcus lactis and Lactococcus cremoris, widely used in dairy fermentations, it is established that CWPSs serve as receptors for several lactococcal bacteriophage (phage) genera, including most commonly encountered Skunaviruses and the P335 group (Mahony et al. 2013, 2017). Given the economic impact of phage‐associated fermentation failure and delays (Guglielmotti et al. 2012), lactococcal CWPSs have been subject to extensive research. Consequently, the phage–host interactions of these two lactococcal species have, perhaps unsurprisingly, received the most significant research attention at the cost of other lactococcal species.
Recently, the genus Lactococcus was reclassified into two separate genera: Lactococcus and Pseudolactococcus (Abe et al. 2025). Currently, 26 species have been identified across these two genera, of which five are associated with milk or dairy environments: the well‐characterized L. lactis and L. cremoris, along with the emerging species Lactococcus hircilactis, Pseudolactococcus laudensis, and Pseudolactococcus raffinolactis (Abe et al. 2025; Mahony et al. 2023). In contrast to L. lactis and L. cremoris, little is known about these emerging species, despite their growing relevance in dairy microbiology (Mahony et al. 2023). Although some preliminary phenotypic characterizations exist (Jung et al. 2020; Klijn et al. 1995; Mahony et al. 2023; Meucci et al. 2015; Tidona et al. 2018), their CWPSs remain unexplored. Investigating these polysaccharides may uncover novel mechanisms of adaptation to dairy environments and provide insights into phage–host interactions, ultimately contributing to more robust fermentation systems. While the CWPSs of pseudolactococcal species have not been studied to date, the extensive analysis of those of L. lactis and L. cremoris has created an excellent foundation to characterize and compare their genetic and structural diversity.
Lactococcal CWPSs that have been characterized thus far consist of a neutral rhamnan component that is covalently attached to, and integrated within, the peptidoglycan layer, along with a surface‐exposed polysaccharide or polysaccharide pellicle (PSP) (Chapot‐Chartier et al. 2010; Mahony et al. 2013, 2020). Current existing models for CWPS biosynthesis include L. cremoris MG1363 (type C) (Theodorou et al. 2019) and L. lactis IL1403 (type B) (Mahony et al. 2020). In these, a dual biosynthetic pathway for both CWPS components is described, in which rhamnan is synthesized in the cytosol by the rhamnan precursor enzymes, RmlABCD and the rhamnosyltransferases RgpABFE, and transported to the outer part of the cell membrane via the ABC transporter system encoded by rgpCD. The PSP synthesis is also initiated in the cytosol by the priming glycosyltransferase, WpsA/YcbB, assisted by the membrane protein WpsB. The PSP is elongated and modified by a series of glycosyltransferases and modification enzymes, such as acetylases, epimerases, and mutases, among others. The PSP chain is subsequently translocated to the outer part of the cell membrane via the WpsG flippase and may or may not be polymerized depending on the presence or absence of the polymerase and co‐polymerase WpsI and WpsH (Theodorou et al. 2019). Finally, the PSP side‐chain is attached to the rhamnan moiety by the product of the gene wpsJ/ycaFG (a comprehensive figure illustrating this model is provided in Supporting Figure S1).
The CWPS components (PSP and rhamnan) are synthesized by a 20–30 kbp gene cluster known as the cwps cluster; however, some lactococcal strains possess glucose side chains attached to their rhamnan or PSP due to the activity of three‐component glycosylation systems (TGS) encoded in distinct loci from the cwps locus (Theodorou et al. 2020). The proximal (5’‐) region of the cwps cluster, which encodes rhamnan biosynthesis, is highly conserved, while the terminal (3’‐) region of the locus encodes PSP biosynthesis and displays significant genetic diversity. This variability likely contributes to the biochemical heterogeneity of CWPS structures, in which the PSP diversity underpins the narrow specificity of lactococcal phages (Mahony et al. 2020). Currently, all assessed lactococcal genomes harbor a cwps gene cluster that may be classified into one of four different genotypes (termed cwps A–D), with 11 subtypes within the C‐type clusters (Parlindungan et al. 2024).
CWPSs participate in several biological functions, including cell division (Theodorou et al. 2019) and cell wall biogenesis (Sadovskaya et al. 2017), defense against phagocytosis by macrophages (Chapot‐Chartier et al. 2010), and resistance against nonspecific host defense mechanisms (Reviewed by Guérin et al. 2022) in addition to their role in phage binding. Recent studies indicate that EPSs and CPSs also play a role in phage–host interactions involving lactococcal P335 phages (Millen et al. 2022), emphasizing the importance of a holistic approach in studying various cell surface‐associated polysaccharides.
EPSs are classified as homopolysaccharides (HoPS), composed of a single repeating monosaccharide, or heteropolysaccharides (HePS), which contain multiple monosaccharides and exhibit greater structural diversity due to variations in glycosidic bonds, molecular weight, and branching (Abarquero et al. 2022; Zhou et al. 2019). The synthesis of HoPS typically occurs via the dextrase/sucrase‐dependent pathways, whereas heteropolysaccharidic EPS and CPS in Gram‐positive bacteria are mainly synthesized en bloc via the Wzx/Wzy pathway (Tytgat and Lebeer 2014). The genes governing HePS biosynthesis are co‐located in the so‐called eps gene clusters, which typically range from 11 to 22 kbp and are variably present in the chromosome or in plasmids in different species (Zeidan et al. 2017). The eps clusters harbor genes associated with polysaccharide assembly (initiation epsE, polymerization wzy, export/flippase wzx, and attachment epsA), modulatory genes (phosphoregulatory module epsBCD), glycosyltransferase‐encoding genes, and genes required for synthesizing activated sugar precursors and the modification of the sugar residues (Cui et al. 2023; Zhou et al. 2019). The genes epsABCDE are located in the proximal region and are highly conserved, while the remainder of the cluster exhibits significant genetic variability. Additionally, L. lactis contains the conserved gene of unknown function epsX in the proximal region, and the gene of unknown function epsL and the orfY modulator represent a second conserved region at the terminal end (Zeidan et al. 2017). Less common pathways for the synthesis of EPS and CPS include the synthase‐dependent pathway, as well as ABC transporter‐dependent pathways commonly reported for Gram‐negative bacteria (Schmid 2018); however, none of these have so far been described in Lactococcus.
EPSs and CPSs may provide protection against environmental stresses, such as low pH or antimicrobial compounds (Caggianiello et al. 2016). Furthermore, they may promote biofilm formation and support probiotic activity by enhancing adhesion, pathogen resistance, and immune modulation (Zhou et al. 2019). Finally, EPSs are highly valued in the food industry due to their water‐retention and stabilizing effects, with the ability to improve the texture, viscosity, and stability of food products (Abarquero et al. 2022; Zhou et al. 2019).
The aim of this study was to investigate and characterize the genetic and structural diversity of cell surface‐associated polysaccharides in the recently reclassified dairy‐associated species P. laudensis and P. raffinolactis. To this end, the cwps and eps clusters of 21 P. laudensis and seven P. raffinolactis strains were identified and compared within and across pseudolactococcal, lactococcal, and other lactic acid bacteria species. Additionally, the CWPS structures of the reference strains P. laudensis DSM28961 and P. raffinolactis DSM 20443 (KACC13441) were resolved using nuclear magnetic resonance (NMR) spectroscopy.
This study represents the first cell wall‐associated polysaccharide analysis of the genus Pseudolactococcus, expanding current knowledge of the diversity and commonalities between the closely related genera Lactococcus and Pseudolactococcus, and providing foundational insights into polysaccharide biosynthesis pathways with potential implications for dairy fermentations and phage–host interactions.
Materials and Methods
2
DNA Extraction and Sequencing
2.1
P. laudensis isolates were obtained from a commercial source (DSM 28961, DSMZ, Braunschweig, Germany) or an undefined mesophilic starter culture (Li et al. 2024) after plating on M17 agar (Oxoid, Hampshire, UK) supplemented with 0.5% lactose (Merck, Darmstadt, Germany) (LM17 0.5%) at 30°C for 48 h. Single colonies were transferred into LM17 1% broth and incubated at 30°C for 48 h. The cultures were centrifuged at 4,402 × g for 10 min in a 5920 R centrifuge (Eppendorf, Hamburg, Germany). The supernatants were discarded, and pellets were resuspended in 1 mL of 0.9% NaCl, transferred into 1.5 mL tubes (Eppendorf, Hamburg, Germany), and centrifuged using a Micro Star 17 R microfuge (VWR, Radnor, USA) at 12,000 × g for 3 min. Genomic DNA was extracted from the resulting pellet utilizing the Bead‐Beat Micro AX 283 Gravity kit (cat # 106‐100‐M1, A&A Biotechnology, 284 Gdynia, Poland) following the instructions of the manufacturer. DNA concentration was measured utilizing a Qubit 4 fluorometer (Thermofisher, Waltham, USA) and normalized to 33.3 ng/μl (400 ng total) for library preparation using the Native Barcoding Sequencing protocol (SQK‐NBD114.24, Oxford Nanopore Technologies, Oxford, UK) following the manufacturer's instructions. Sequencing was performed on a PromethION 2 Solo platform. Basecalling was conducted with Dorado v0.7.1 (https://github.com/nanoporetech/dorado) using the [email protected] basecalling model.
Genome Assembly and Reorientation
2.2
Raw data were filtered using NanoFilt v2.6.0 (De Coster et al. 2018) to a minimum quality of 8 and a minimum length of 1,000 bp. Assembly was performed utilizing Trycycler v0.5.4 (Wick et al. 2021) with the data obtained after splitting the reads into 12 subsets and assembling those with Flye v2.9.1 (Kolmogorov et al. 2019), Canu v2.2 (Koren et al. 2017), and Hybracter v0.9.0 (Bouras et al. 2024b) for optimized plasmid recovery (Bouras et al. 2024b; Johnson et al. 2023).
The complete genomes from the P. raffinolactis strains were obtained from the National Center for Biotechnology Information (NCBI).
All chromosome and plasmid contigs in this study, including those from P. raffinolactis obtained from NCBI, were reoriented with dnaapler v0.8.1 (Bouras et al. 2024a) to start with dnaA or repA, respectively.
The accession numbers of the genomes utilized in this study are indicated in Supporting Table S1.
Identification and Characterization of the cwps and eps Clusters
2.3
For the identification of the cwps and eps clusters, the genomes of the P. laudensis and P. raffinolactis strains were annotated utilizing Bakta v1.9.4 (Schwengers et al. 2021) and subjected to nucleotide Basic Local Alignment Search Tool (BLAST) (Camacho et al. 2009) analysis against well‐characterized reference clusters. Specifically, cwps clusters from L. lactis IL1403 (accession number CP033607.1, positions 196221 to 220264 bp) and L. cremoris MG1363 (accession number AM406671.1, positions 194607 to 218654 bp) were selected as they represent model strains with extensively studied cwps loci and are representatives of two of the three major cwps genotypes established to date, that is, cwps B and C, respectively (Mahony et al. 2020; Theodorou et al. 2019). Likewise, the eps cluster from L. lactis KLDS 4.0325 (accession number CP006766.1, positions 84302 to 101534 bp) was selected due to its complete annotation (Zeidan et al. 2017). Annotation files were manually curated for downstream analyses.
Nucleotide and amino acid sequence similarities were calculated using BLAST (Camacho et al. 2009), and these are reported in the manuscript as the product of Query Cover × Percentage Identity values. Gene cluster similarities and alignments were visualized using Clinker (Gilchrist and Chooi 2021) via the CAGECAT webserver, release 1.0 (van den Belt et al. 2023). Additionally, protein structure prediction with DeepTMHMM v1.0 (Hallgren et al. 2022), as well as analyses with HHPred v 57c8707149031cc9f8edceba362c71a3762bdbf8 (Hildebrand et al. 2009), Conserved Domain Database (CDD) v3.21 (Wang et al. 2023), and Prokka v1.14.6 (Seemann 2014) were performed to further characterize the cwps and eps genes/gene products.
To characterize the genetic context of both cwps and eps loci, the genomes of the P. laudensis and P. raffinolactis strains were analyzed with Phastest v3.0 (Wishart et al. 2023) to determine the presence of prophages and IslandCompare v1.0 (Bertelli et al. 2022) for the identification of other potential genomic islands. Furthermore, regions flanking the cwps and eps loci were analyzed with ICEfinder from the ICEberg 3.0 webserver (Wang et al. 2024) and mobileOG‐db v1.1.3 (Brown et al. 2022) through the Proksee webserver (Grant et al. 2023).
Extraction and Purification of Polysaccharides and Their Fragments
2.4
Pseudolactococcus laudensis DSM 28961
2.4.1
Cells derived from ~1.3 L of a fresh overnight culture of P. laudensis strain DSM 28961, grown in LM17 1% media, were harvested by centrifugation at 5,000 × g for 15 min and treated by autoclaving (120°C, 30 min). The cell debris was harvested by centrifugation (yielding ~11 g of debris) and treated first in 5% trichloroacetic acid (TCA) in water at 5°C for 48 h. Subsequently, the sample was treated with hot 0.01 M hydrochloric acid (HCl) for 20 min under stirring in a boiling water bath (100°C), followed by cooling, centrifugation at 12,000 x g, and resuspendion in 0.1 N HCl for a second 20 min treatment under the same conditions (Sadovskaya et al. 2017; Parlindungan et al. 2024). This approach facilitates the extraction of fractions enriched with either “outer cell wall (CW) layer” PSP (cold TCA extract) or “inner CW layer” rhamnan (often 0.1 M HCl extract). The following yields of crude extracts were obtained: autoclaved (AU) extract—200 mg; TCA extract—100 mg; 0.01 M HCl extract ~100 mg; 0.1 M HCl extract—46 mg. All extracts contained l‐dTal (6‐deoxytalose), d‐Glc (glucose), and d‐Gal (galactose) in different ratios as major monosaccharide components. The AU extract also contained arabinose. Traces of arabinose and xylose were present in other extracts. AU and 0.1 M HCl extracts were further fractionated by anion‐exchange chromatography. After desalting, fractions were screened by monosaccharide analysis and ^1^H‐NMR spectroscopy. Two CWPS fractions named CWPS1 and CWPS2 were eluted with ~ 0.1 M NaCl and 0.2–0.3 M NaCl, respectively. The neutral fraction contained a mixture of CWPS1 and CWPS2. A portion of CWPS1 was treated with NH_4_OH (50°C, 30 min) and desalted on a Sephadex G‐15 column to yield the de‐O‐acetylated CWPS1 (DPS).
Pseudolactococcus raffinolactis DSM 20443
2.4.2
The cell pellet derived from 4 L of a fresh overnight culture of P. raffinolactis DSM 20443 grown in tryptic soy yeast extract (TSB) broth supplemented with 0.3% yeast extract was first extracted by autoclaving (120°C, 30 min). Cell debris was removed by centrifugation, and the supernatant was deproteinated through the addition of 5% TCA, dialyzed, and lyophilized, yielding the crude AU extract (110 mg). Cell debris (~5 g) was subjected to consecutive acid extractions as described previously (Parlindungan et al. 2024), yielding a crude TCA extract (3 mg), a 0.01 M HCl extract (50 mg), and a 0.1 M HCl extract (45 mg).
Upon fractionation on a Sephadex G‐50 column, HCl extracts yielded one homogeneous peak (rhamnan) containing l‐Rha (rhamnose) and d‐Glc in an approximate ratio of 2.3:1 and trace amounts of GlcNAc (N‐acetyl glucosamine) and d‐Gal.
The AU extract eluted into two fractions: a high molecular weight (HMW) fraction with a composition identical to the rhamnan from the HCl extract and a low molecular weight (LMW) fraction, containing d‐Glc and d‐Gal in an approximate ratio of 1.3:1. The LMW fraction was further purified by anion‐exchange chromatography on Q‐Sepharose. The major fraction, eluted at the beginning of the gradient, was collected and desalted on a Biogel P2 column and named OS1 (Oligosaccharide 1).
General and Analytical Methods
2.5
Chromatography, monosaccharide and methylation analysis, electrospray ionization mass spectrometry (ESI‐MS), and NMR spectroscopy were performed as described previously (Mahony et al. 2020; Parlindungan et al. 2024). The absolute l‐configuration of 6‐deoxy‐d‐talose (6dTal) was determined by GC‐MS of the acetylated (R)‐(‐)‐2‐butyl glycoside derivative by comparison with an authentic standard. The O‐acetylated homopolysaccharide (OPS) from Pseudomonas (Burkholderia) plantarii DSM 6535 was used as the source of D‐6dTal (Zähringer et al. 1997).
Results
3
Identification of cwps Clusters in P. laudensis and P. raffinolactis Genomes
3.1
The genomes of 21 P. laudensis strains were sequenced and assembled in the context of this study to investigate genes involved in cell surface‐associated polysaccharide biosynthesis. All assemblies yielded complete genomes comprising a circular chromosome and a variable number of circular plasmids. Additionally, the analysis included all seven P. raffinolactis genomes currently available as complete assemblies in public databases (NCBI).
The cwps clusters from all P. laudensis and P. raffinolactis strains were identified and extracted for further genomic analysis. All retrieved genomes were found to harbor a cwps cluster ranging in size from 18,445 to 30,217 bp and encoding between 16 and 28 genes. The strains were subsequently classified into genotypes based on their similarity regarding the PSP‐encoding region of their cwps cluster, that is, the terminal region of the cluster, starting from wpsA, which is predicted to encode the priming glycosyltransferase. The following criteria were used to assign the strains into different cwps genotypes: (i) the presence or absence of polymerase‐ and co‐polymerase‐encoding genes, (ii) the presence or absence of a flippase‐encoding gene, and (iii) the occurrence of two or more PSP‐encoding genes—excluding the flippase, polymerase, and co‐polymerase—sharing < 30% amino acid similarity with another genotype. Based on these criteria, eight novel cwps genotypes were identified: five exclusive to P. raffinolactis, one unique to P. laudensis, and two shared by a single strain of each species. A visual representation comparing the (pseudo)lactococcal cwps clusters is provided in Figure 1, highlighting the sequence similarity and unique regions of the respective clusters. The comparative analysis allowed the identification of novel genotypes using similar approaches to previous studies of lactococcal cwps genotyping, that is, the presence of at least one distinct glycosyltransferase‐encoding gene, which has been established to be associated with distinct saccharidic composition/structures. This analysis revealed a broader diversity of CWPSs in P. laudensis/raffinolactis than in L. lactis/cremoris, in which only four genotypes are described (A‐D) with similar classification criteria (Mahony et al. 2020), despite a greater diversity of the latter regarding their isolation sources. In that sense, all the P. laudensis strains except the reference DSM 28961 were isolated from the same commercial starter culture (Li et al. 2024; this study).
Overview of the organization and sequence similarity of the cwps gene clusters of P. laudensis (Pl.), P. raffinolactis (Pr.) types E‐L, and selected L. lactis (Ll.) and L. cremoris (Lc.) types A, B, C, and D. The proposed functions for each of the genes are color‐coded and have been inferred by gene annotation, as well as sequence and structure similarity. Red dashed lines separate the rhamnan (left) and PSP (right) encoding regions of the clusters.
Among the studied strains, P. raffinolactis Lr_18_12S was shown to exhibit the greatest similarity to known L. lactis and L. cremoris cwps genotypes, particularly to type A L. lactis UC509.9 (Figure 1). These findings, along with shared genotypes between P. laudensis and P. raffinolactis, suggest that additional shared genotypes may exist in other Lactococcus/Pseudolactococcus species. Thus, the novel genotypes were designated E–L, continuing the established cwps nomenclature. Table 1 details the distribution of genotypes, intra/inter‐species prevalence, as well as the genomic locations and length of the cwps clusters.
The most prevalent cwps genotype among the assessed strain collection was type F, present in 19 of the 21 P. laudensis strains. While these strains exhibit practically identical nucleotide composition in their cwps clusters, deeper analysis revealed transposase insertions at varying sites, disrupting the predicted wpsJ gene—responsible for transferring the PSP side chain onto the rhamnan backbone—and a transmembrane modification enzyme (Fig. S2). Additionally, the cwps cluster in strains T2H4 and T2G5 was found to be split into two in distant sites within the chromosome, most certainly due to the action of the aforementioned transposase, since it moved the genes from its position to the next transposase situated downstream of the cwps cluster. The functional consequences of these insertions and rearrangements on CWPS biosynthesis remain to be experimentally validated; however, their consistent occurrence in strains from the same starter culture may reflect selective pressure to modify or eliminate the PSP component, likely as an adaptive response to phage predation or niche adaptation to the processing environment. Regarding the other cwps genotypes, E and G were each present in a strain of both P. laudensis and P. raffinolactis, while the rest were unique to single strains (Table 1).
Similar to the previously studied L. cremoris and L. lactis, the proximal region of the clusters containing the deoxy‐thymidine‐disphospho‐rhamnose (dTDP‐Rha) precursor genes rmlABCD, the rhamnosyltransferases rgpAB, and the ABC transporter system rgpCD is highly conserved among all the lactococcal strains, though the latter genes are located further in the cluster in type E strains. Conversely, the PSP‐encoding regions exhibited high variability among the different clusters, especially in the number and composition of glycosyltransferases, other transferases, and modification enzymes (Figure 1).
Functional Predictions of the PSP Side Chain‐Encoding Regions of the cwps loci
3.2
The PSP priming glycosyltransferase WpsA, its assisting membrane anchor WpsB, the flippase WpsG, responsible for PSP translocation across the cell membrane, and WpsJ, involved in attaching the PSP to the rhamnan backbone (Theodorou et al. 2019), play essential roles in the PSP biosynthesis. In most Pseudolactococcus strains analyzed, these genes were either conserved or could be confidently inferred based on TMHMM modeling predictions or HHPred structural homology predictions (Theodorou et al. 2019) (Figure 1; Supporting Data). Specifically, when nucleotide or amino acid similarity was insufficient for identification, wpsJ and wpsG were inferred based on analysis of gene products with transmembrane DUF2142‐containing proteins and predicted structures with 12–14 transmembrane helices (TMHs), respectively (Hvorup et al. 2003; Kuk et al. 2022). However, in genotypes E and G, neither sequence homology nor structural predictions enabled identification of wpsA, wpsB, wpsG, or wpsJ homologs, strongly suggesting that these genotypes utilize an alternative CWPS assembly mechanism, distinct from the dual biosynthetic pathway previously characterized in Lactococcus.
Among the studied strains, only P. raffinolactis Lr_18_12S (type K) harbored gene products with sequence similarity to the putative WpsI and WpsH oligosaccharide polymerase and co‐polymerase proteins from L. lactis and L. cremoris type C and D strains. DeepTMHMM analysis confirmed these findings, predicting protein structures identical to those of L. cremoris NZ9000 (type C_1_): WpsI containing a DUF2142 domain and 11 TMHs, while WpsH featuring a large loop in the outer part of the cell membrane flanked by a signal peptide and a TMH in the N‐ and C‐termini, respectively.
DeepTMHMM analysis also revealed WpsI‐ and WpsH‐like candidates in several other strains. Specifically, a gene annotated by Bakta as a bactoprenol glycosyltransferase exhibited two TMHs at the terminal end, resembling the WpsH co‐polymerase structure, which contains a single TMH instead. This gene was found at the 3’‐end of the corresponding cwps loci, adjacent to genes annotated as glycosyltransferases containing a DUF2142 and 10‐12 TMHs, which we hypothesize encode the polymerase function. The presence of these genes in all P. laudensis and P. raffinolactis cwps genotypes, except in type J, suggests that the strains produce polymeric PSP, as is the case for the type C and D L. lactis and L. cremoris strains studied to date (Mahony et al. 2020; Parlindungan et al. 2024; Theodorou et al. 2019). In contrast, in type J, the PSP is likely an oligomer.
Among the identified DUF2142 domain‐containing putative polymerases, three groups can be distinguished: (i) proteins with lactococcal genotype C and D WpsI‐like predicted structures, although with low amino acid sequence similarity, found in types I and L; (ii) proteins containing an ALG6/ALG8 domain, identified as by CDD in type F strains; and (iii) proteins containing a Protein O‐Mannosyltransferase 2 superfamily domain as indicated by HHPred and CDD analyses, present in the type E and G clusters. The latter of these three putative polymerase groups exhibits significant structure similarity with YcaA from the type B L. lactis IL1403, the only gene product within this cluster that has not yet been functionally assigned (Mahony et al. 2020). This raises the possibility that YcaA encodes a polymerase with limited or defective activity due to the absence of an associated co‐polymerase. Other YcaA‐like proteins are present in several of the cwps loci, in variable locations within the clusters (Figure 1). Alternatively, YcaA‐like proteins could be involved in glycan modification, since Protein‐O‐mannosyltransferases are conserved among domains of life and known to be responsible for protein O‐mannosylation (Lommel and Strahl 2009). Finally, the structure of WpsJ is very similar to this of the WpsI polymerase in type C and D strains, such as L. cremoris NZ9000. Thus, it is possible that these DUF2142‐containing YcaA‐like proteins function as WpsJ PSP‐tranferases in strains including L. lactis KLDS4.0325 and P. raffinolactis APC3967, given the lack of other candidates performing such functions (Figure 1). Experimental validation is required to confirm the putative functions of these genes, which is beyond the scope of the present study.
Identification of the eps Clusters
3.3
The recent discovery of EPS‐dependent lactococcal phage–host interactions (Millen et al. 2022) accentuates the need to adopt a holistic approach when studying cell surface‐associated polysaccharides among lactococci and beyond. Among the studied strains, all except P. laudensis T2E11 and P. raffinolactis Lr_19_5 and APC3967 were found to harbor an eps cluster. A schematic representation of the eps clusters is presented in Figure 2. Based on sequence similarity, 11 distinct genotypes (I–XI) were identified. The distribution of these genotypes, along with the length and genomic location of the eps clusters, is detailed in Table 1.
Overview of the organization and sequence similarity of the eps gene clusters of P. laudensis (P.) and P. raffinolactis (Pr.) types I‐XI, L. lactis (Ll.) KLDS 4.0325, and selected Enterococcus faecalis and Enterococcus hirae strains. The proposed functions for each of the genes are color‐coded and have been inferred by gene annotation, as well as sequence and structure similarity.
The epsRXABCD genes at the proximal end of the cluster are conserved among the I, III, IV, V, and IX genotypes, whereas genotypes II, VI, VII, and VIII contain only phosphoregulatory epsABC genes (Figure 2). Within these genotypes, types I, II, and IV possess a flippase‐encoding gene with similarity to the wzx in L. lactis KLDS 4.0325, while all other types possess a gene encoding a protein with the typical membrane topology and characteristics of flippases (i.e., 12‐14 transmembrane helices) from the multidrug/oligosaccharidyl‐lipid/polysaccharide (MOP) exporter superfamily (Hvorup et al. 2003; Kuk et al. 2022). Bakta annotations revealed the presence of genes encoding the likely wzy polymerase within the gene clusters of eps genotypes V and VII, and DeepTMHMM enabled the inference of genes encoding transmembrane polymerases for genotypes I‐IX, confirming those of the V and VII types (Figure 2; Supporting Data). The presence of the regulatory‐associated genes, in addition to the flippase‐ and polymerase‐encoding genes, indicates that the strains belonging to the I‐IX genotypes likely synthesize heterosaccharidic EPS en bloc via the Wzy‐dependent pathway (Tytgat and Lebeer 2014).
In contrast, genotypes X and XI harbor an ABC transporter system, lack the signature genes of the Wzx/Wzy pathway, and share gene similarity with genotypes I–IX only in the epsL gene of unknown function and the orfY modulator (Figure 2). The Wzy‐independent or ABC transporter‐dependent pathway is associated with the sequential synthesis and export of HePS, which remain bound to the cell envelope, that is, CPS, in Gram‐negative bacteria (Schmid 2018; Whitfield et al. 2024). To the best of our knowledge, no ABC transporter‐dependent pathways have been reported for the synthesis of EPS or CPS in Gram‐positive bacteria.
The different eps genotypes exhibit an elevated degree of variability regarding their glycosyltransferases and modification enzymes composition. For example, strains like T2G11 (type III) encode 8 different glycosyltransferases and only a single modification enzyme annotated as a putative phosphotransferase. Conversely, type VI strains possess 4 glycosyltransferases and diverse modification enzymes, including putative acetyltransferases, dehydratases, and epimerases (Figure 2). Such genetic variability among the different genotypes is presumed to underlie the potential structural diversity among the synthesized EPS and/or CPS; however, it remains to be proven that this genetic potential is linked to the actual production of EPS under laboratory or food production conditions.
Interestingly, eps genotypes VI and VII exhibit an elevated sequence similarity to eps clusters from certain Enterococcus strains (Figure 2). Nucleotide BLAST revealed > 99.5% nucleotide similarity between type VI clusters and Enterococcus hirae FDAARGOS_1124 and E. hirae FDAARGOS_1123 across the entire cluster, as well as > 97% nucleotide similarity between type VII and Enterococcus faecalis CQNBZ21‐135 and E. hirae FDAARGOS_1057, spanning from the start of the cluster to the Wzy‐like polymerase‐encoding gene (Figure 2). These E. faecalis and E. hirae strains were isolated in feces in 2021 in China and from an unknown source in Germany, respectively. Given their sequence similarity, and the variability of glycosyltransferases and modification enzymes encoded by eps clusters, it is unlikely that these clusters evolved independently in strains from different genera.
The intriguing similarity between the eps loci of P. laudensis and the aforementioned Enterococcus strains motivated further investigation regarding the genetic context of these clusters, with the aim of elucidating their evolutionary origins and potential horizontal gene transfer (HGT) events. Similarly, exploring the genetic context of the cwps clusters may reveal additional insights into their functional organization and mobility.
Genetic Context of the eps Gene Clusters
3.4
Both eps and cwps gene clusters of all analyzed P. laudensis and P. raffinolactis strains were observed to be chromosomally located. The finding of chromosomally encoded eps loci in these species is in contrast to the majority of L. lactis/cremoris eps loci, which are more commonly plasmid‐associated (Parlindungan et al. 2024). To characterize their genetic context and explore their possible origin and mobility, the genomic regions flanking both loci were analyzed.
Bakta and mobileOG‐db annotations suggested a HGT origin for the eps clusters. In all P. laudensis and P. raffinolactis strains, the regions directly up‐ and downstream of the eps cluster contained several genes annotated as phage and plasmid‐associated genes, transposases, and conjugal transfer proteins. Additionally, these regions also contained putative antimicrobial resistance genes (i.e., Enterocin A immunity) and phage defense systems (e.g., AbiV in T2C1 or T2A4), commonly associated with HGT (Makarova et al. 2011).
IslandCompare analysis confirmed that the eps loci, along with adjacent regions, constitute genomic islands integrated into the lactococcal chromosomes. After manual curation, genomic islands were determined to range in size from ~45 to 55 kbp, with the upstream regions consistently larger than the downstream regions (Figure 3). For all the P. laudensis strains, as well as P. raffinolactis WiKim0068 and DSM 20443, these genomic islands were integrated in the same location within their chromosome, as can be observed in Table 1 by the location of the eps cluster. Interestingly, the island was conserved in all P. laudensis strains, including T2E11, which lacks an eps cluster.
Genetic context of the eps cluster in selected lactococcal and enterococcal strains, showing (left) the upstream region and (right) the downstream region. For the P. laudensis (Plau.), P. raffinolactis (Lr.), and P. paracarnosus (Pp.) strains, the regions represent genomic islands. For L. lactis (Ll.) KLDS 4.0325 and E. hirae (Eh.) and E. faecalis (Ef.) strains, the size of the flanking regions was decided arbitrarily. The eps cluster has been manually removed for better visualization of the up‐ and downstream regions.
Nucleotide BLAST analysis revealed elevated sequence similarity between the genomic islands of all P. laudensis and P. raffinolactis strains, particularly in the upstream regions (80–98% identity). Interestingly, a comparable level of similarity was observed with a chromosomal region in the meat isolate Pseudolactococcus paracarnosus TMW 2.1615, the only strain of its species with a complete genome available in NCBI (Figure 3). Notably, this region also precedes an eps gene cluster in P. paracarnosus.
Further analysis with Phastest indicated that the eps upstream flanking regions of P. laudensis and P. raffinolactis strains contain a putative incomplete prophage, yet the predictions were inconsistent between the strains despite their sequence similarity. Moreover, in P. laudensis DSM 28961, the eps downstream region was identified as an intact prophage. However, the latter does not present a typical prophage structure (i.e., no apparent lysis, structural, or replication modules), and most annotated phage genes comprise transposases, along with a couple of glycosyltransferases, suggesting a false positive result from Phastest from what appears to be a mobile genetic element (MGE). Notably, the insertion site of the MGE is within the eps cluster, specifically downstream of the second transposase (Figure 2). This insertion modifies the genetic composition of the eps cluster and likely influences EPS composition in this strain. A comparable mechanism occurs in pathogenic bacteria such as Shigella flexneri, where prophage‐encoded glycosylation or acetylation modifies the O‐antigen, a process known as serotype conversion (Jakhetia et al. 2013; Markine‐Goriaynoff et al. 2004).
Finally, ICEfinder analysis identified the upstream and downstream regions of the eps clusters as integrative conjugative elements (ICEs) and integrative mobile elements (IMEs), respectively. Although the upstream regions contain genes encoding a predicted relaxase, a type IV secretion system (T4SS), and a type IV coupling protein (T4CP)‐encoding gene, they appear to lack an origin of transfer (OriT), preventing their self‐transmission.
The eps clusters of E. hirae FDAARGOS_1124, FDAARGOS_1123, and FDAARGOS_1057 and E. faecalis CQNBZ21‐135 were analyzed to investigate their genetic context. In E. hirae FDAARGOS_1124 and FDAARGOS_1123, the identified eps clusters are located on plasmids, whereas in E. faecalis CQNBZ21‐135, the eps cluster is inserted in the chromosome flanked by MGEs. This strongly suggests that these clusters were acquired through HGT. Conversely, in E. hirae FDAARGOS_1057, the eps cluster was located in the chromosome but showed no association with mobile elements or HGT markers, suggesting it is native to this strain. Thus, it may represent the origin of eps genotype VII. Notably, aside from the eps clusters themselves, these Enterococcus strains did not share sequence similarity in their adjacent regions with the Pseudoactococcus strains analyzed in this study (Figure 3).
Genetic Context of the cwps Gene Clusters
3.5
Unlike the eps clusters, the cwps clusters were not identified as part of genomic islands and appear to be stably located within the chromosomes, consistent with previous findings in L. lactis and L. cremoris (Mahony et al. 2020). These species generally harbor the cwps clusters in positions ranging from ~150,000 bp to ~250,000 bp within the chromosome, similar to most P. raffinolactis strains analyzed in this study and P. laudensis DSM 28961 and T2E11. However, some intrachromosomal mobility was observed, as described earlier, where transposase insertions caused genomic rearrangements in P. laudensis T2H4 and T2G5 (Table 1).
All P. laudensis and P. raffinolactis strains share common genes flanking their cwps clusters. In the region upstream of the predicted cwps locus, these are genes that encode proteins that may assist in CWPS biosynthesis and cell function but are not part of the primary biosynthetic functions, including a DUF4649 domain‐containing protein (annotated as ribose‐phosphate pyrophosphokinase in some Firmicutes), a 30S ribosomal protein, and a translation elongation factor, likely ensuring efficient protein synthesis. Additionally, a septation ring regulator (ezrA) suggests coordinated CWPS integration into the peptidoglycan layer during cell division. Downstream genes may aid CWPS maturation and cell wall integration, including molecular chaperones (groES and groEL), which ensure proper protein folding, and genes encoding endo‐β‐N‐acetylglucosaminidase, glutamyl aminopeptidase, and CHAP domain‐containing proteins, likely involved in cell wall remodeling.
Notably, the flanking regions of P. raffinolactis and P. laudensis cwps loci differ significantly from those in L. lactis IL1403, UC509.9, KLDS 4.0325, and L. cremoris NZ9000 (used as reference strains for the species). Apart from a ribosomal protein gene upstream and an amidase (N‐acetylmuramoyl‐l‐alanine amidase) downstream, no other genes are apparently sharing functionality. Common genes in L. lactis and L. cremoris strains include those predicted to encode FeoA and FeoB iron transporters, a metalloprotease, an IMP dehydrogenase, and a NadD nicotinate‐nucleotide adenylyltransferase, among others.
Structure of Cell Surface‐Associated Polysaccharides
3.6
P. laudensis DSM 28961
3.6.1
To establish the chemical structure of cell surface‐associated polysaccharides (i.e., CWPS, CPS, and EPS) in emerging dairy pseudolactococcal species, the polysaccharides of the reference strains P. laudensis DSM 28961 and P. raffinolactis DSM 20443 were extracted.
Fractions obtained after separation of the different extracts of P. laudensis DSM 28961 on an anion‐exchange column contained l‐dTal, d‐Glc, and d‐Gal in different ratios. The absolute configuration of 6‐deoxytalose was determined as the l‐form by GC‐MS of the acetylated (R)‐(‐)‐2‐butyl derivative by comparison with the authentic standard, 6‐deoxy‐d‐talose from the O‐acetylated PS from Pseudomonas (Burkholderia) plantarii DSM 6535 (Zähringer et al. 1997) (Supporting Figure S3).
2D NMR analysis allowed to identify two distinct CWPSs: CWPS1, containing a relatively rare monosaccharide 6‐deoxy‐l‐talose, and a β‐(1‐4)‐galactan, CWPS2 (Figure 4ab, Supporting Table S2, Fig. S4). CWPS1 was found to be identical to the polysaccharide from Bifidobacterium breve 7017 (Manuscript in preparation), differing only by the degree of O‐acetylation (100% acetate in B. breve 7017% and 20% in P. laudensis DSM 28961, Supporting Figure S4 ). Upon deacylation, polysaccharides from both species produced an identical polysaccharide product.
CWPS structures. (a) CWPS1 and (b) CWPS2 in P. laudensis DSM 28961; (c) Rhamnan and (d) PSP moieties in P. raffinolactis DSM 20443.
CWPS2 is a common polysaccharide in plants, typically encountered as a side chain of rhamnogalacturonan I (Liwanag et al. 2013). Some bacteria such as Bacillus licheniformis can metabolize this polysaccharide (Ryttersgaard et al. 2004), yet to the best of our knowledge, it has not been previously reported as a bacterial CWPS.
P. raffinolactis DSM 20443
3.6.2
P. raffinolactis 20443 was subjected to consecutive extractions, followed by fractionation on a Sephadex G50 column. HMW fractions from all extracts had similar ^1^H NMR spectra, although they did not look like a pure single (homogeneous) compound. These fractions contained rhamnose and glucose and were designated here as rhamnan.
NMR spectra of the rhamnan, that is, Correlated Spectra (COSY), Total Correlated Spectroscopy (TOCSY), Rotating frame Overhauser Effect Spectroscopy ROESY, ^1^H‐^13^C Heteronuclear Single Quantum Coherence (HSQC), and ^1^H‐^13^C Heteronuclear Multiple Bond Correlation (HMBC), contained several spin‐systems of α‐rhamnose and one α‐glucose (all in pyranose form). 2D ^1^H‐ and ^13^C spectra were fully assigned (Supporting Table S3, Fig. S5). The following Nuclear Overhauser Effect (NOE) cross‐peaks were observed: A1:B1,2,3,4,5; B1:A2; B1:E1; C1:D2; D1:C3 and HMBC: A1:B3; B1:A2; C1:D2; D1:C3 cross‐peaks were observed. This allowed us to conclude that there were two main repeating units, differing by the presence of glucose side chains (Figure 4c). There was approximately twice as much of the branched structure as the linear structure.
Methylation analysis of the rhamnan indicated the presence of 2‐, 3‐, and 2,3‐linked Rha and terminal Glc, in agreement with the structure established by NMR.
These units could be present in one or different PS chains, but there were no indications to decide what is correct. Spectra contained many small rhamnose signals, which could be due to linkages between different repeating units.
The structure of the rhamnan is essentially identical to one of the rhamnose‐glucose polysaccharides from Bifidobacterium breve 2003 and its EPS^‐^ mutant derivatives (Manuscript in preparation), except for the Glc residue E (Figure 4d), which is in an α‐configuration in P. raffinolactis and in a β‐configuration in B. breve.
The LMW product OS1, obtained from the AU extract, contained only Glc and Gal and had a distinct ^1^H‐NMR spectrum compared to the rhamnan. NMR analysis of the OS1 extract using 2D methods (COSY, TOCSY, ROESY, ^1^H‐^13^C HSQC, ^1^H‐^13^C HMBC, ^1^H‐^31^P HMQC) showed that it was a hexasaccharide with a mixture of α‐ and β‐Glc at the reducing end (Supporting Figure S6 and S7). To remove signal duplication due to anomeric configurations of Glc X, OS1 was reduced with NaBD_4_ to yield OS1red and analyzed by NMR (Fig. S6, Table S3). The following transglycoside NOE and HMBC correlations were observed: A1:D3; B1:A6, C1:B4, D1:C4; E1:B6 (Supporting Figure S7). This ultimately allowed the determination of the sequence of saccharides as presented in Figure S6.
OS1 contained a phosphate ester, which yielded a ^31^P signal at 1.72 ppm. It correlated with the A‐3 proton, indicating phosphorylation at terminal α‐Gal A3. The composition of the OS was confirmed by positive ion mode ESI MS (for the reduced with NaBD_4_ OS MS calc. 1073, observed neg. ion 1071.9 amu). We assume that OS1 represents the repeating unit of the PSP. Similar to the recently described PSP of L. lactis Tempeh6L (Parlindungan et al. 2024), the repeating units are most probably linked via an acid‐labile glycosyl phosphate‐phosphodiester bond between Glc F and α‐Gal A at position 3 (F‐1‐P‐3‐A). Upon cleavage of the unstable glycosyl phosphate bond, the phosphate group remains at position 3 of the α‐Gal A. Anomeric configuration of glycosylphosphates in Lactococcus polysaccharides is usually α, which is presented in the proposed structure of the original polysaccharide (Figure 4d and Supporting Figure S6).
Establishing the Genotype‐Structure Links
3.7
Previous studies (Mahony et al. 2020; Parlindungan et al. 2024; Theodorou et al. 2019) explored the relationship between lactococcal cwps loci and CWPS structures, paving the way for future structure prediction efforts. This study integrates genetic and chemical analyses of P. laudensis DSM 28961 and P. raffinolactis DSM 20443 to link cwps genes to their corresponding CWPS structures.
In P. laudensis DSM 28961, two independent CWPS structures, a 6‐deoxy‐αL‐talan (CWPS1) and a β‐(1,4)‐galactan homopolysaccharide (CWPS2), were identified. The CWPS1 structure resembles a lactococcal type B disaccharide rhamnan structure with a glucose side chain. The corresponding cwps locus contains the conserved lactococcal dTDP‐Rha precursor genes rmlABCD, and the rhamnosyltransferases rgpAB, followed by an additional rhamnosyltransferase and a NAD+‐dependent oxidoreductase, annotated as dTDP‐6‐deoxy‐l‐talose 4‐dehydrogenase (EC 1.1.1.134). This enzyme catalyzes a reversible reaction, interconverting dTDP‐6‐deoxy‐β‐l‐talose and dTDP‐4‐dehydro‐β‐l‐rhamnose (Mäki and Renkonen 2004), explaining the final 6‐deoxy‐α‐l‐talan structure (Figure 5a). Furthermore, a transmembrane protein with 10 TMHs was identified as an OafA superfamily acetylase by CDD, explaining the observed O‐acetylation at the C moiety (Figure 4a). The last modification enzyme, annotated as a sulfatase‐like hydrolase/transferase with 11 predicted TMHs, requires further investigation to elucidate its specific function. The glucose side chain could be either added by the remaining glycosyltransferase in the cwps locus (Figure 5a) or by other mechanisms, such as the TGS described for L. cremoris NZ9000 (Theodorou et al. 2020). In that sense, a homology search using nucleotide and protein BLAST was conducted to identify TGS candidates in the P. laudensis DSM 28961 genome, specifically homologs of the L. cremoris NZ9000 TGS genes, that is, csdABCDEF (Theodorou et al. 2020). The analysis revealed potential candidates, of which a system with a gene annotated as a putative membrane protein, followed by a hypothetical protein and a Bactoprenol glycosyltransferase, encoded in the chromosome in the positions 782310 to 785539 bp, is the most likely candidate. The Bactoprenol glycosyltransferase gene shares 56.3% amino acid sequence similarity and identical TMHs distribution with CsdA from L. cremoris NZ9000, whereas the putative membrane protein shares structure similarity with CsdB, but not sequence homology. Finally, the hypothetical protein possesses a structure resembling that of the WpsH co‐polymerase. Given that CsdAB is responsible for rhamnan glycosylation in L. cremoris NZ9000, it is reasonable to hypothesize that the described system in P. laudensis DSM 28961 may have the same role. Finally, the presence of genes resembling the YcaA‐like putative polymerase and WpsH‐like putative co‐polymerase from L. lactis and L. cremoris C and D genotypes is likely responsible for the talan polymerization (Figure 5a).
Models of heteropolysaccharidic CWPSs biosynthesis in (a) P. laudensis DSM 28961 and (b) P. raffinolactis DSM 20443. The proposed pathways remain to be validated by mutational analyses. Abbreviations: Ac, acetyl group; 6dTal, 6‐deoxy‐talose; Gal, galactose; Glc, glucose; GlcNAc, N‐acetylglucosamine; GT, glycosyltransferase; P, phosphate; Rha, rhamnose; RT, rhamnosyltransferase; TDP, thymidine diphosphate; Und‐P, undecaprenyl phosphate. Figure created with BioRender.
The β‐(1,4)‐galactan homopolysaccharide (CWPS2) is not linked to CWPS1, and that makes feasible that it is synthesized independently by genes encoded outside the cwps cluster. In Lactobacillus johnsonii FI9785, a bactoprenol glycosyltransferase is responsible for the synthesis of an EPS glucan homopolysaccharide (Mayer et al. 2020). Therefore, the system encoded in the positions 782310 to 785539 bp, described in the former paragraph, is also a likely candidate for the synthesis of this structure. Alternatively, CWPS2 could be synthesized by the glycosyltransferase within the cwps locus (Figure 4a), exported via the rgpCD ABC transporter, and polymerized by the YcaA‐like putative polymerase and WpsH‐like putative co‐polymerase. Experimental validation is necessary to confirm the genes encoding the synthesis of CWPS2.
The CWPS of P. raffinolactis DSM 20443 consists of a rhamnan and a PSP moiety, typical of lactococcal species, and aligns with the structure of its cwps locus. This locus is clearly divided into the PSP‐encoding region (starting from the priming glycosyltransferase WpsA) and the rhamnan‐encoding region (spanning from the RmlA nucleotidyltransferase up to, but excluding, WpsA) (Figure 5b).
The rhamnan of P. raffinolactis DSM 20443 is composed of a rhamnose disaccharide, resembling those of lactococcal B‐type strains (Vinogradov et al. 2018) and Streptococcus thermophilus UCC St50 and ST64987 (Lavelle et al. 2022; McDonnell et al. 2020). Furthermore, this rhamnan structure is eventually further decorated with a glucose side chain, likely added by a glycosyltransferase encoded within the rhamnan‐encoding region of the cwps cluster (Figure 5b) or, alternatively, by a TGS encoded elsewhere within the genome. Protein BLAST results revealed a glycosyltransferase in position 2041507 to 2042436 bp sharing 56.5% amino acid sequence similarity with the CsdA from L. cremoris NZ9000; however, no apparent candidates were found for a putative CsdB‐like encoding gene.
The PSP of P. raffinolactis DSM 20443 consists of a repeating hexasaccharide unit, composed of a linear pentasaccharide of galactose‐glucose with a galactose side chain. This hexasaccharidic structure matches the six glycosyltransferases, including the WpsA priming glycosyltransferase, present within the PSP‐encoding region of the cluster. This suggests that no TGS system located elsewhere within the genome has an influence on the final PSP structure, such as csdCD in L. cremoris NZ9000 (Theodorou et al. 2020). It is predicted that the glycosyltransferases encoded within this cluster sequentially add the individual saccharides to the growing PSP side chain subunits as has been proven in L. lactis/cremoris (Figure 5b); however, this requires validation through mutational analysis (Theodorou et al. 2019).
Regarding its glucose‐galactose composition, the NAD‐dependent epimerase within the cluster likely catalyzes the interconversion of UDP‐glucose to UDP‐galactose (Figure 5b), ensuring the availability of both monosaccharides required for PSP synthesis.
Finally, an unmatched feature is the phosphate group attached to the A and F moieties (Figure 4d). Given the lack of an apparent kinase or phosphotransferase within the cwps gene cluster, the most plausible explanation is that these galactose and glucose moieties are phosphorylated before their participation in the CWPS biosynthesis, where they are incorporated as such into the PSP.
Discussion
4
CWPSs are involved in several functions, including cell division and cell wall biogenesis, defense against phagocytosis, or bacteriophage adsorption (Chapot‐Chartier et al. 2010; Mahony et al. 2013, 2017; Theodorou et al. 2019), among others. Given these essential roles, it is not surprising that cwps clusters have been identified for the totality of studied L. lactis and L. cremoris up to date (Mahony et al. 2020; Parlindungan et al. 2024). The present study provides evidence for their presence in all analyzed P. laudensis and P. raffinolactis strains, suggesting that CWPSs have equally critical roles in these species. The rmlABCD and rgpABCD genes, located at the proximal end of the cwps clusters, that is, in the rhamnan encoding region, are highly conserved across L. lactis and L. cremoris strains. Our analysis revealed that these genes are conserved as well in P. laudensis and P. raffinolactis, confirming their utility as marker genes for identifying cwps clusters in other (pseudo)lactococcal species.
The current study identified eight novel cwps genotypes (E–L), significantly expanding the diversity of these gene clusters within the Lactococcus and Pseudolactococcus genera, which previously comprised four genotypes (A–D), including 11 subtypes within type C (C_1_‐C_11_) (Parlindungan et al. 2024).
The A–D genotypes feature a cwps cluster where the rhamnan and PSP components are biosynthesized and exported independently across the cell membrane, where they are subsequently assembled by WpsJ, as proposed in existing models (Mahony et al. 2020; Theodorou et al. 2019). Notably, a similar cwps organization was observed in all the newly identified P. laudensis and P. raffinolactis genotypes, except for E and G.
In the E and G cwps genotypes, the absence of genes encoding key PSP biosynthesis components, such as the priming glycosyltransferase WpsA, the assisting membrane protein WpsB, the WpsG flippase responsible for translocating the PSP to the external side of the cell membrane, and WpsJ, which likely attaches the PSP to the rhamnan component (Theodorou et al. 2020), suggests a CWPS assembly pathway distinct from previously described Lactococcus models. It is reasonable to speculate that, in these strains, the entire rhamnan‐PSP structure may be assembled on the inner side of the cell membrane and subsequently transported as a complete unit via ABC transporters. Additionally, these cwps clusters encode fewer glycosyltransferases, indicating a less complex side chain. Interestingly, in the type E strains, only a single glycosyltransferase was identified alongside the encoded rhamnosyltransferases, raising the possibility of an absent PSP component.
Chemical analysis of the reference strain P. laudensis DSM 28961 (type E) revealed the presence of two distinct CWPSs: a 6‐deoxy‐α‐l‐talan polysaccharide (CWPS1) and β‐(1,4)‐galactan homopolysaccharide (CWPS2). These findings represent novel discoveries in (pseudo)lactococcal CWPS structures. First, CWPS1 replaces the rhamnan component typically observed in Lactococcus strains with a 6‐deoxy‐α‐l‐talose‐containing backbone. This substitution highlights a unique genetic or enzymatic adaptation within type E strains. Second, CWPS2 is a homopolysaccharide, contrasting with all previously described lactococcal CWPS, which are characterized by heteropolysaccharidic structures. Finally, the structural independence of CWPS1 and CWPS2 confirms the absence of a typical PSP in P. laudensis DSM 28961. In the classical definition of CWPS, the polysaccharide pellicle (PSP) acts as a side chain attached to the rhamnan core. Here, the two polysaccharides are not covalently linked, confirming a distinct mechanism of CWPS biosynthesis and assembly in type E strains. Additionally, this study also provided the CWPS structure of the reference P. raffinolactis DSM 20443 (type I), confirming its classical rhamnan‐PSP structure and increasing the ever‐expanding catalog of (pseudo)lactococcal CWPS compositions (Supporting Tables S4 and S5).
Recent studies (Mahony et al. 2020; Parlindungan et al. 2024; Theodorou et al. 2019, 2020) have established the foundation for CWPS structure predictions based on the genetic composition of the cwps loci. The present study contributes to this growing knowledge base by identifying heretofore undiscovered genotypes and resolving novel CWPS structures. Lactococcal phages primarily utilize the PSP as a receptor for identifying and binding to their hosts (Mahony et al. 2017). Therefore, accurate prediction of CWPS, particularly of their PSP region, would represent a significant step toward palliating fermentation failures in the dairy industry caused by phage infections. Notably, some lactococcal phages are known to utilize receptors other than CWPSs, namely membrane proteinaceous receptors in ceduoviruses (Millen and Romero 2016) or other saccharidic structures such as CPSs, as have recently been described for certain P335 phages (Millen et al. 2022).
The findings of Millen et al. (2022) highlighted the importance of adopting a holistic approach when studying cell surface‐associated polysaccharides. In line with this, genomic inspection of P. laudensis and P. raffinolactis strains revealed eps loci for HePS production in 25 out of 28 strains, identifying 11 distinct genotypes (I–XI). These findings suggest that most of the analyzed strains are producers of either EPS or CPS. Notably, all eps clusters were shown to be located within the chromosomes of these strains.
Further genomic analysis revealed the HGT origin of the eps clusters, which apparently had been integrated into the chromosomes as part of genomic islands on recombination hotspots or putative grounded prophages. These regions are known to be hubs for the integration of defense islands, antimicrobial resistance genes, and other genes conferring advantageous traits for their bacterial hosts (Ramisetty and Sudhakari 2019), in this case, EPS or CPS biosynthesis. Additionally, this study highlights the mobility of eps clusters, providing evidence of inter‐genera HGT between enterococcal and type VI and VII P. laudensis strains. Moreover, it unraveled the presence of what appears to be an ABC transporter‐dependent pathway for the synthesis of EPS or CPS for type X and XI P. raffinolactis strains, a pathway typically exclusively associated with CPS production in Gram‐negative bacteria (Schmid 2018).
Under the growth and extraction conditions applied to P. laudensis DSM 28961 and P. raffinolactis DSM 20443, no significant yields of either EPS were observed; therefore, their structures could not be resolved. Since production of EPSs is growth‐condition dependent (Cui et al. 2023), further investigation is needed to ascertain the conditions supporting their production in these strains to elucidate later their chemical structures.
Conclusions
5
The current study has provided novel insights into the emergent dairy species P. laudensis and PP. raffinolactis, expanding the known genetic diversity and metabolic pathways for CWPS and EPS/CPS biosynthesis within the dairy‐associated representatives of the Lactococcus‐Pseudolactococcus clade. It also sheds light on the cwps and eps gene clusters and their genetic contexts, while presenting the first resolved CWPS structures for pseudolactococcal species, thereby laying down a solid foundation for future investigations.
These findings not only deepen our understanding of Pseudolactococcus cell surface‐associated polysaccharides but also offer valuable insights into microbial adaptation mechanisms and phage resistance, with significant potential for improving industrial fermentation systems.
Finally, further research should include targeted gene knockouts combined with structural analysis to validate the CWPS biosynthetic pathways. Furthermore, to strengthen the predictions of the activity of the specific glycosyltranferases (and other gene products), additional CWPS structures will need to be elucidated in the future. Additionally, transcriptomic studies performed under varied growth conditions will be essential to elucidate the regulatory mechanisms and the conditions driving EPS/CPS expression and production.
Author Contributions
Axel Soto‐Serrano: writing – original draft, writing – review and editing, investigation, visualization, validation, formal analysis, software, project administration, data curation, methodology. Irina Sadovskaya: data curation, methodology, investigation, writing – review and editing, visualization, validation, formal analysis, resources. Evgeny Vinogradov: data curation, formal analysis, investigation, methodology, resources, writing – review and editing, validation. Wenwen Li: investigation, methodology, writing – review and editing. Jun‐Hyeok Yu: writing – review and editing, investigation. Kelsey White: investigation, writing – review and editing. Douwe Sinderen: writing – review and editing, resources, supervision, funding acquisition. Lukasz Krych: writing – review and editing, resources, supervision, methodology, software, funding acquisition. Paulina Deptula: funding acquisition, investigation, project administration, formal analysis, supervision, resources, data curation, validation, writing – review and editing. Jennifer Mahony: project administration, formal analysis, data curation, supervision, resources, methodology, validation, writing – review and editing, funding acquisition, investigation, conceptualisation.
Ethics Statement
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Figure S1: Generic dual chain assembly model for lactococcal CWPS assembly. Figure S2: Overview of the organization and sequence similarity of the cwps gene clusters of P. laudensis type F. Figure S3: Determination of the absolute configuration of 6dTal. Figure S4: Overlap of the HSQC spectra of the CWPS of P. laudensis DSM 28961 (green), B. breve 7017 (red), and O‐deacylated B. breve 7017 (black). Figure S5: ¹H–¹³C HSQC of the rhamnan from P. raffinolactis DSM 20443. Figure S6: Structure of the oligosaccharide products OS1 and OS1red from P. raffinolactis DSM 20443 and a proposed structure of PSP. Figure S7: HSQC spectrum of OS1 from P. raffinolactis DSM 20443 (blue‐green) and HMBC correlations from the anomeric protons in magenta. “a” and “b” denote α‐ and β‐anomers, respectively. Table S1: Accession numbers. Table S2: ¹H and ¹³C NMR data (δ, ppm, D₂O, 25°C, 600 MHz) for the galactan from L. laudensis DSM 28961. Table S3: ¹H and ¹³C NMR data (δ, ppm, D₂O, 25°C, 600 MHz) for the rhamnan from P. raffinolactis DSM 20443. Table S4: NMR data for the free (OS) and reduced (OSred) oligosaccharides from P. raffinolactis DSM 20443 (δ, ppm; Bruker AVANCE III 600 MHz, 25°C). Table S5: Chemical structures of the rhamnans of selected lactococcal strains. Table S6: Chemical structures and characteristics of PSP components of cwps C‐ and D‐type lactococcal strains.
MO SupplementaryData 060825.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abarquero, D. , E. Renes , J. M. Fresno , and M. E. Tornadijo . 2022. “Study of Exopolysaccharides From Lactic Acid Bacteria and Their Industrial Applications: A Review.” International Journal of Food Science & Technology 57: 16–26. 10.1111/ijfs.15227. · doi ↗
- 2Abe, K. , M. Yuki , Y. Imagawa , et al. 2025. “Genome‐Based Reclassification of the Genus Lactococcus and Two Novel Species Pseudolactococcus Yaeyamensis Gen. Nov., Sp. Nov. and Lactovum Odontotermitis Sp. Nov. Isolated From the Gut of Termites.” International Journal of Systematic and Evolutionary Microbiology 75, no. 6. 10.1099/ijsem.0.006803.PMC 1228173740471659 · doi ↗ · pubmed ↗
- 3van den Belt, M. , C. Gilchrist , T. J. Booth , Y. H. Chooi , M. H. Medema , and M. Alanjary . 2023. “CAGECAT: The Comparative Gene Cluster Analysis Toolbox for Rapid Search and Visualisation of Homologous Gene Clusters.” BMC Bioinformatics 24: 181. 10.1186/s 12859-023-05311-2.37131131 PMC 10155394 · doi ↗ · pubmed ↗
- 4Bertelli, C. , K. L. Gray , N. Woods , et al. 2022. “Enabling Genomic Island Prediction and Comparison in Multiple Genomes to Investigate Bacterial Evolution and Outbreaks.” Microbial Genomics 8, no. 5. 10.1099/mgen.0.000818.PMC 946507235584003 · doi ↗ · pubmed ↗
- 5Bouras, G. , S. R. Grigson , B. Papudeshi , V. Mallawaarachchi , and M. J. Roach . 2024 a. “Dnaapler: A Tool to Reorient Circular Microbial Genomes.” Journal of Open Source Software 9: 5968. 10.21105/joss.05968. · doi ↗
- 6Bouras, G. , G. Houtak , R. R. Wick , et al. 2024 b. “Hybracter: Enabling Scalable, Automated, Complete and Accurate Bacterial Genome Assemblies.” Microbial Genomics 10, no. 5. 10.1099/mgen.0.001244.PMC 1116563838717808 · doi ↗ · pubmed ↗
- 7Brown, C. L. , J. Mullet , F. Hindi , et al. 2022. “mobile OG‐db: A Manually Curated Database of Protein Families Mediating the Life Cycle of Bacterial Mobile Genetic Elements.” Applied and Environmental Microbiology 88, no. 18: e 0099122. 10.1128/aem.00991-22.36036594 PMC 9499024 · doi ↗ · pubmed ↗
- 8Caggianiello, G. , M. Kleerebezem , and G. Spano . 2016. “Exopolysaccharides Produced by Lactic Acid Bacteria: From Health‐Promoting Benefits to Stress Tolerance Mechanisms.” Applied Microbiology and Biotechnology 100: 3877–3886. 10.1007/s 00253-016-7471-2.27020288 · doi ↗ · pubmed ↗