FAMA and bHLH090: Coordinating myrosin cell development and herbivory defense
Marcella Teixeira

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
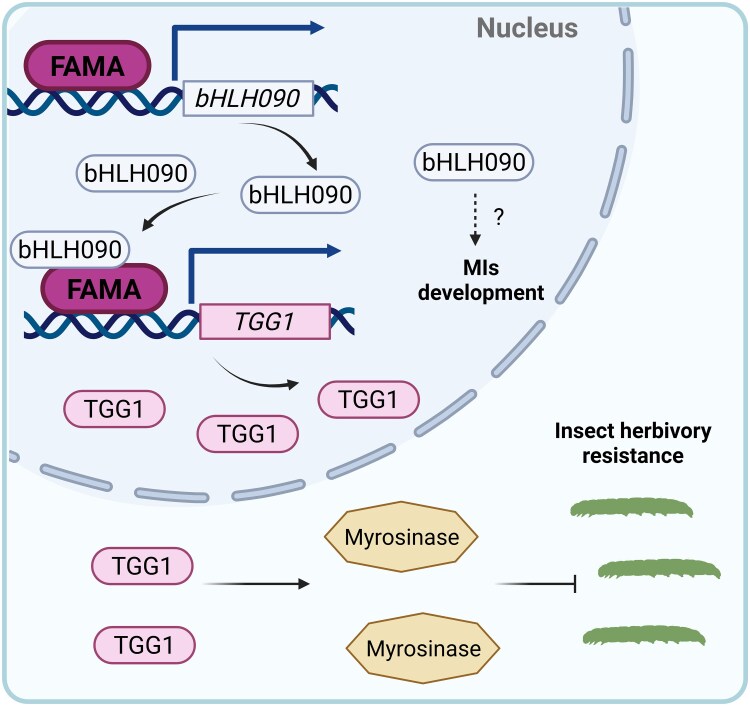 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Cell death mechanisms and regulation · Polysaccharides and Plant Cell Walls
Plants are exposed to different stresses during their life cycle. To fight off herbivores, the Brassicaceae family has developed a defense system known as “mustard oil bomb.” Under normal conditions, glucosinolates (GSLs) and myrosinase, which breaks down GSLs into compounds toxic to herbivores, are kept in separate compartments but positioned close enough to be quickly mixed (Luthy and Matile 1984; Ratzka et al. 2002; Shirakawa and Hara-Nishimura 2018). When an herbivore feeds and breaches plant tissue, the myrosinase and GSLs react and the bomb goes off, resulting in a burst of toxic metabolites that halt the herbivore attack. In a brilliant evolutionary adaptation, Brassicaceae plants show that survival often depends on timely and precise response to attack.
The “mustard oil bomb” has been well characterized in Arabidopsis. Myrosinases, encoded by Thioglucoside Glucohydrolase (TGG) genes, accumulate in specialized myrosin cells (MCs), including guard cells (GCs) and myrosin idioblast cells (MIs). Loss of 2 TGGs in the tgg1 tgg2 double mutant increases herbivore susceptibility (Barth and Jander 2006). TGGI and TGG2 accumulation and MI production are regulated by the basic-helix-loop-helix (bHLH) transcription factor FAMA (Li and Sack 2014; Shirakawa et al. 2014, 2016), although its targets in MI development are yet to be determined. Interestingly, the expression of another transcription factor, bHLH090, is upregulated in FAMA-overexpressing plants (Hachez et al., 2011; Shirakawa et al. 2014).
In this issue of Plant Physiology, Zhou et al. (2025) investigated the roles of bHLH090 in MI development and herbivory response. First, using a bHLH090 promoter-driven GUS reporter, the authors demonstrated spatial overlap of bHLH090 expression with the described MI distribution (Barth and Jander 2006; Shirakawa et al. 2014). Genetic analysis revealed that bHLH090 overexpression boosts MI production and myrosinase activity, culminating in enhanced resistance to insect herbivory (Fig. 1).
Given FAMA's function in MI development and bHLH090 high expression in FAMA-overexpressing lines, the authors investigated FAMA's role in bHLH090 regulation by introducing pbHLH090:GUS into the fama-1 mutant. They discovered that bHLH090 expression was reduced in fama mutant, suggesting that bHLH090 expression in MIs is dependent on FAMA function. Consistently, transient expression in Nicotiana benthamiana showed that FAMA activated bHLH090 expression. The authors further showed that bHLH090 activation is a result of FAMA's direct binding to bHLH090 promoter through chromatin immunoprecipitation-qPCR and electrophoretic mobility shift assay.
If bHLH090 is the sole target of FAMA, we would expect that overexpression of bHLH090 in fama-1 mutant plants would be sufficient to rescue the defects of MI development in fama. Interestingly, bHLH090 overexpression in fama-1 background could not rescue fama-1 mutants' MI defect, suggesting that FAMA's role in MI development is more than simply activating bHLH090 expression. Therefore, besides activation of bHLH090 expression, FAMA is necessary for bHLH090 function. Because bHLH transcription factors have been shown to form protein complexes to regulate gene expression (Kanaoka et al. 2008), the authors hypothesized that FAMA and bHLH090 would be part of a regulatory protein complex. Consistently, FAMA and bHLH090 physically interact, as demonstrated by results from yeast 2-hybrid, pull down, and firefly luciferase complementation imaging.
Finally, because TGG1 is regulated by both FAMA and bHLH090, the authors asked if the interaction between FAMA and bHLH090 regulates TGG1 expression. Using a transient expression assay, they showed that coexpression of FAMA and bHLH090 results in increased expression of TGG1, revealing a synergistic effect between the two proteins (Fig. 1).
In an orchestrated dance, Arabidopsis uses the transcription factor bHLH090 to amplify a defense response through the activation of TGG1 (Fig. 1). By enhancing or rewiring this defense switch, Brassicaceae crops can be engineered to start a rapid and localized response against herbivores, reducing dependence on chemical pesticides. On a broader perspective, understanding how plants balance the energetic cost of defense with growth can inform strategies for designing stress-resilient crops.
On the molecular mechanism's perspective, the work presented by Zhou et al. (2025) raises several questions. Which other bHLH family members might have a role in MI development? Since bHLH090 overexpression was insufficient to rescue MI development in fama-1, what additional factors are involved in MI development and function? How do herbivory cues integrate with the FAMA-bHLH090-TGG1 cascade to trigger timely activation of the defense bomb?
The Zhou et al. (2025) study adds a new layer of complexity to the mustard oil bomb and opens the door to leveraging this evolutionary innovation in agriculture. The challenge now is to expand these insights beyond Arabidopsis, translating molecular choreography into major crops in field-ready strategies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barth C, Jander G. Arabidopsis myrosinases TGG 1 and TGG 2 have redundant function in glucosinolate breakdown and insect defense. Plant J. 2006:46(4):549–562. 10.1111/j.1365-313X.2006.02716.x 16640593 · doi ↗ · pubmed ↗
- 2Hachez C, Ohashi-Ito K, Dong J, Bergmann DC. Differentiation of Arabidopsis guard cells: analysis of the networks incorporating the basic Helix-Loop-Helix transcription factor, FAMA. Plant Physiol. 2011:155(3):1458–1472. 10.1104/pp.110.16771821245191 PMC 3046599 · doi ↗ · pubmed ↗
- 3Kanaoka MM, Pillitteri LJ, Fujii H, Yoshida Y, Bogenschutz NL, Takabayashi J, Zhu JK, Torii KU. SCREAM/ICE 1 and SCREAM 2 specify three cell-state transitional steps leading to Arabidopsis stomatal differentiation. Plant Cell. 2008:20(7):1775–1785. 10.1105/tpc.108.06084818641265 PMC 2518248 · doi ↗ · pubmed ↗
- 4Li M, Sack FD. Myrosin idioblast cell fate and development are regulated by the Arabidopsis transcription factor FAMA, the auxin pathway, and vesicular trafficking. Plant Cell. 2014:26(10):4053–4066. 10.1105/tpc.114.12972625304201 PMC 4247575 · doi ↗ · pubmed ↗
- 5Luthy B, Matile P. The mustard oil bomb: rectified analysis of the subcellular organization of the myrosinase system. Biochem Physiol Pflanz. 1984:179(1–2):5–12. 10.1016/S 0015-3796(84)80059-1 · doi ↗
- 6Ratzka A, Vogel H, Kliebenstein DJ, Mitchell-Olds T, Kroymann J. Disarming the mustard oil bomb. Proc Natl Acad Sci U S A. 2002:99(17):11223–11228. 10.1073/pnas.17211289912161563 PMC 123237 · doi ↗ · pubmed ↗
- 7Shirakawa M, Hara-Nishimura I. Specialized vacuoles of myrosin cells: chemical defense strategy in Brassicales plants. Plant Cell Physiol. 2018:59(7):1309–1316. 10.1093/pcp/pcy 08229897512 · doi ↗ · pubmed ↗
- 8Shirakawa M, Ueda H, Nagano AJ, Shimada T, Kohchi T, Hara-Nishimura I. FAMA is an essential component for the differentiation of two distinct cell types, myrosin cells and guard cells, in Arabidopsis. Plant Cell. 2014:26(10):4039–4052. 10.1105/tpc.114.12987425304202 PMC 4247577 · doi ↗ · pubmed ↗