Co-cultivation rescues suicidal Paenibacillus amylolyticus swarms
Dana Ronin, Mads Frederik Hansen, Mette Burmølle

TL;DR
Co-cultivating two bacteria allows one to rescue the other from a non-growing state by stabilizing the environment's pH.
Contribution
Demonstrates how interspecies interactions can rescue bacterial swarms from functional decline through pH stabilization.
Findings
Dual-species swarms of Paenibacillus amylolyticus and Stenotrophomonas maltophilia are thicker and allow S. maltophilia to expand into new areas.
S. maltophilia rescues P. amylolyticus from a non-growing state by alkalizing the environment.
Monospecies swarms of P. amylolyticus lose growth ability over time despite retaining reductase activity.
Abstract
Bacterial locomotion is integral to acquiring resources and getting access to new niches. Swarming, a type of motility where flagellated bacteria cooperatively move together across a semi-solid surface, is one example of how bacteria can colonize new territories. This collective behavior is temporally and spatially orchestrated, requiring task specialization of community members. In this study, we paired a swarming bacterium, Paenibacillus amylolyticus, with a non-swarmer, Stenotrophomonas maltophilia, to investigate the impact on fitness of each strain. In dual-species conditions, the community swarm became significantly thicker and improved the ability of S. maltophilia to range into new territories. Swarming enabled P. amylolyticus to cross barriers of antimicrobials, whereas the thicker, dual-species swarm did not empower S. maltophilia to cross. Comparative studies of population…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
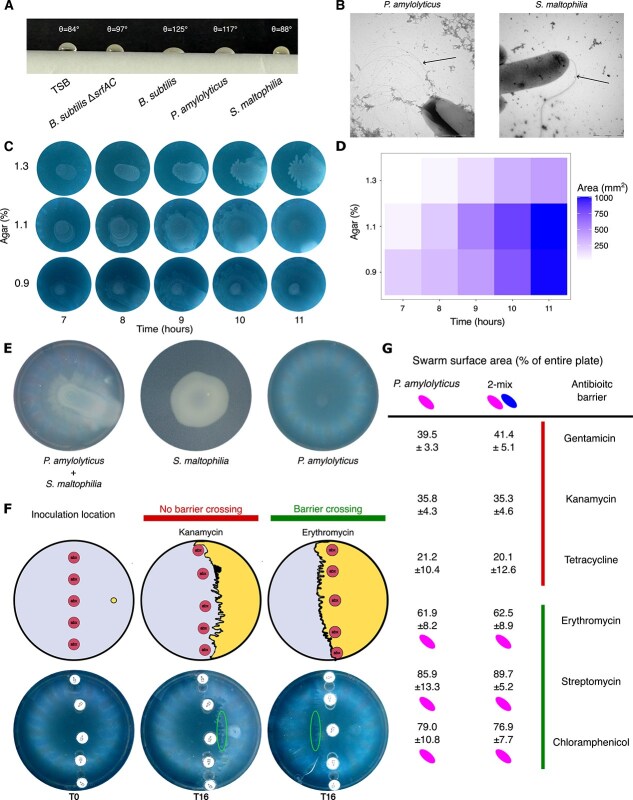 Figure 1
Figure 1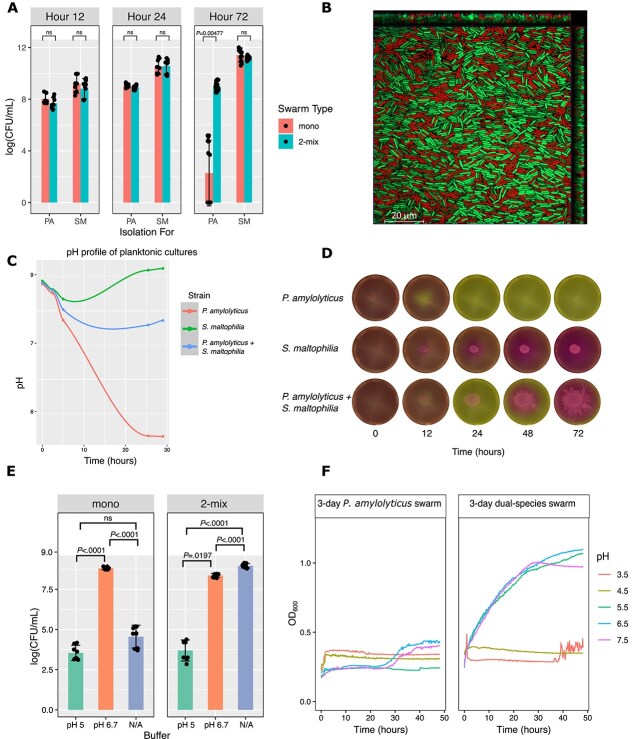 Figure 2
Figure 2- —European Research Council10.13039/100010663
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial biofilms and quorum sensing · Micro and Nano Robotics · Evolutionary Game Theory and Cooperation
For many living organisms, locomotion is an essential aspect of life. The ability to migrate according to environmental conditions and stimuli enables positioning in favorable habitats and determines success [1, 2]. In nature, cells are often organized in multicellular populations that require coordinated behavior. One such example is swarming, in which flagellated bacteria produce biosurfactants to reduce surface tension and collectively migrate through a thin film [2]. This collective behavior facilitates expansion over nutrient sources and translocation to new ecological niches. Over time, swarm expansion is a dynamic process with a high level of heterogeneity, cell–cell contact, and intercellular interactions [3, 4]. Most studies have focused on single species swarm development; however, bacteria typically live in multispecies environments [5]. Given previously described interspecies interactions such as cross-feeding, pH stabilization, and spatial intermixing [6–8], we investigated the effect of pairing swarming Paenibacillus amylolyticus with non-swarming Stenotrophomonas maltophilia. To evaluate how co-swarming affected the fitness of both members, we measured two key indicators: (i) access to new niches, assessed by surface expansion and antibiotic tolerance and (ii) cell numbers of each member in mono- and dual-species swarms.
Many Paenibacillus spp. can swarm [9–11]; one well-known swarming requirement is surfactant production. To verify surfactant production in P. amylolyticus, we identified genes with homology to those in the Bacillus subtilis surfactin synthetase cluster (Suppl. Fig. 1) [12]. Surfactin is synthesized by non-ribosomal peptide synthesis (NRPS) systems, three of which were previously identified in the P. amylolyticus genome [13]; low homology hits for surfactin synthesis may also include related genes from other NRPS systems. Also, the presence of a surface tension-reducing compound in the culture was verified (Fig. 1A). Additionally, swarming motility is powered by rotating flagella movement [2]; we identified lophotrichous flagella in P. amylolyticus (Fig. 1B) and observed swarming behavior at various agar concentrations, with reduced swarm speed at higher concentrations (Fig. 1C and D). Once swarming initiated, P. amylolyticus formed a thin swarm layer (Suppl. Fig. 2) that spread over the entire agar plate after about 11 h of incubation (Fig. 1D, 0.9% agar). When mixed with S. maltophilia, the swarm layer became significantly thicker (Fig. 1E; Suppl. Fig. 2). This thicker part was visibly asymmetric (Fig. 1E), indicating that S. maltophilia could perform passive sliding motility, using surfactant as a public good to access new areas [14]. To determine whether S. maltophilia was translocated along with P. amylolyticus, we sampled throughout the dual-species swarm from the center to the edge. At the swarm periphery, P. amylolyticus constituted the forefront of the community, whereas S. maltophilia was present solely in the thicker areas of the swarm (Suppl. Fig. 2). Nevertheless, S. maltophilia displayed territory expansion when it gained the ability to spread in a dual-species swarm.
Previous studies have shown that swarm behavior can desensitize the susceptibility to antibiotics [15, 16]. Gaining antibiotic tolerance during motility can provide protection when colonizing new niches. We tested the ability of mono- and dual-species swarms to cross antibiotic barriers. Neither mono- nor dual-species swarms could cross kanamycin, gentamicin, or tetracycline barriers (Fig. 1F,G), which is consistent with the essential swarmer, P. amylolyticus, being sensitive to these compounds (Suppl. Fig. 3). For erythromycin, streptomycin, and chloramphenicol barriers, both the mono- and dual-species swarms were able to cross (Fig. 1F,G). P. amylolyticus is resistant to streptomycin, but sensitive to erythromycin and chloramphenicol (Suppl. Fig. 3, Suppl. Fig. 4). When determining which members crossed the antibiotic barrier in the dual-species swarms, solely P. amylolyticus crossed (Fig. 1G); the presence of S. maltophilia in the dual-species swarm did not provide any apparent advantage for antibiotic susceptibility that was not already achieved by the monospecies swarm on its own. S. maltophilia could not utilize the P. amylolyticus-associated surfactant to expand its reach into new areas, even across barriers for which it has resistance (e.g., kanamycin) (Fig. 1G and F). Although being in a dual-species swarm did not induce further antibiotic tolerance, swarming motility in general benefited P. amylolyticus, allowing it to grow in the presence of antibiotics it is normally sensitive to (erythromycin and chloramphenicol).
Having established that swarming resulted in growth area expansion both in presence and absence of antibiotics, we next assessed specific abundances as a second measure of fitness during co-swarming. P. amylolyticus and S. maltophilia abundances were quantified over time in mono- and dual-species swarms. There was no significant difference between the two conditions for S. maltophilia cell counts at any of the respective timepoints (Fig. 2A), indicating no benefit or cost for this species during co-swarming despite the territory expansion previously reported. For 12- and 24-h old swarms, there was no difference in P. amylolyticus cell counts between the mono- and dual-species swarms. However, for 72-h old swarms, we observed significantly lower cell counts for the monospecies P. amylolyticus swarm compared to the dual-species swarm (Fig. 2A). Thus, instead of competing for shared resources, P. amylolyticus required the presence of S. maltophilia to uphold a viable population of cells. To test whether P. amylolyticus cells were dead after 72 h in the monospecies swarm, we applied a bacterial reductase activity stain, which revealed a plethora of active and vital cells (Fig. 2B), suggesting that they were not dead but had entered a non-growth state.
A previous study found that Xanthomonas perforans modulated the swarming behavior of Paenibacillus spp. by pH fluctuations [11]. Given that P. amylolyticus and S. maltophilia have pH stabilization attributes when together (Fig. 2C) [8], we applied phenol red to monitor pH changes over time during swarming. When P. amylolyticus was swarming independently, it greatly acidified its environment after 24 h (Fig. 2D), meaning that 72-h old swarms were exposed to an acidic environment for an extended period. However, in the dual-species swarm, S. maltophilia increased the pH after an initial low pH caused by P. amylolyticus; the pH of the swarming plate increased after 24 h, starting from the center and expanding to the edges over time (Fig. 2D). To verify that pH was the key determinant of P. amylolyticus survival after 72 h of swarming, swarming plates were buffered to pH 5.0 and pH 6.7. Phenol red was used to confirm the stability of the pH buffer over time, despite bacterial growth (Suppl. Fig. 5). Similarly to co-cultivation with S. maltophilia, monospecies swarming plates buffered to pH 6.7 ensured a high abundance of P. amylolyticus after 72 h of growth. Conversely, dual-species swarming plates buffered to pH 5.0 reduced the P. amylolyticus cell counts to levels comparable to those recorded in unbuffered monospecies swarms (Fig. 2E). Finally, to test if P. amylolyticus was rescued after entering its non-growing state, we re-cultured 3-day old P. amylolyticus swarms in liquid TSB at different pHs (ranging from 3.5 to 7.5). No significant growth was detected over 48 h, indicating that a change in pH did not rescue the cells. In fact, low pH (pH < 5.5) also prevented regrowth of P. amylolyticus cells originating from dual-species swarm communities (Fig. 2F).
“Ecological suicide” has been previously described in Paenibacillus sp.; at increasing cell densities, Paenibacillus sp. strongly acidifies its environment causing its own extinction [17]. In our study, when P. amylolyticus was paired with S. maltophilia, the latter raised the pH after a period of low pH (Fig. 2C and D), effectively rescuing P. amylolyticus from its self-imposed cytotoxic environment. Our data demonstrate how interspecies interactions may benefit co-culture partners; P. amylolyticus directly benefitted from the presence of S. maltophilia, which rescued it from extinction. Even though S. maltophilia did not benefit in cell numbers when in the dual-species swarm, it co-localized along with P. amylolyticus and gained access to new areas. In natural environments, such colony expansion may give access to new niches, supporting growth as well as survival.
Supplementary Material
figure1_spellingFixedinG_wraf225
supplMaterialCompiled_wraf225
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sourjik V, Wingreen NS. Responding to chemical gradients: bacterial chemotaxis. Curr Opin Cell Biol 2012;24:262–8. 10.1016/j.ceb.2011.11.00822169400 PMC 3320702 · doi ↗ · pubmed ↗
- 2Partridge JD . Surveying a swarm: experimental techniques to establish and examine bacterial collective motion. Appl Environ Microbiol 2022;88:e 01853–21. 10.1128/aem.01853-2134878816 PMC 8824199 · doi ↗ · pubmed ↗
- 3Jeckel H, Jelli E, Hartmann R. et al. Learning the space-time phase diagram of bacterial swarm expansion. Proc Natl Acad Sci USA 2019;116:1489–94. 10.1073/pnas.181172211630635422 PMC 6358709 · doi ↗ · pubmed ↗
- 4Jeckel H, Nosho K, Neuhaus K. et al. Simultaneous spatiotemporal transcriptomics and microscopy of Bacillus subtilis swarm development reveal cooperation across generations. Nat Microbiol 2023;8:2378–91. 10.1038/s 41564-023-01518-437973866 PMC 10686836 · doi ↗ · pubmed ↗
- 5Tan CH, Lee KWK, Burmølle M. et al. All together now: experimental multispecies biofilm model systems. Environ Microbiol 2017;19:42–53. 10.1111/1462-2920.1359427878947 · doi ↗ · pubmed ↗
- 6Herschend J, Damholt ZBV, Marquard AM. et al. A meta-proteomics approach to study the interspecies interactions affecting microbial biofilm development in a model community. Sci Rep 2017;7:16483. 10.1038/s 41598-017-16633-629184101 PMC 5705676 · doi ↗ · pubmed ↗
- 7Liu W, Russel J, Burmølle M. et al. Micro-scale intermixing: a requisite for stable and synergistic co-establishment in a four-species biofilm. ISME J 2018;12:1940–51. 10.1038/s 41396-018-0112-229670216 PMC 6052071 · doi ↗ · pubmed ↗
- 8Herschend J, Koren K, Røder HL. et al. In vitro community synergy between bacterial soil isolates can be facilitated by p H stabilization of the environment. Appl Environ Microbiol 2018;84:e 01450–18. 10.1128/AEM.01450-1830143509 PMC 6193382 · doi ↗ · pubmed ↗