The complete chloroplast genome sequence of the endangered species: Ephedra rhytidosperma (Ephedraceae): insights into genome features and evolutionary relationships
Xiulian Liao, Jinfeng Niu, Yan Wang, Yuqing Wei, Lei Zhang

TL;DR
This paper presents the full chloroplast genome of Ephedra rhytidosperma, an endangered plant, and explores its genetic structure and evolutionary links to other Ephedra species.
Contribution
The first complete chloroplast genome assembly for Ephedra rhytidosperma, providing insights into its genome features and phylogenetic relationships.
Findings
The chloroplast genome is 109,545 bp long with a GC content of 36.8% and encodes 118 complete genes.
Phylogenetic analysis shows E. rhytidosperma and six other Ephedra species form a monophyletic group.
The genome provides a foundation for future genetic and genomic studies in the Ephedra genus.
Abstract
Ephedra rhytidosperma belongs to the genus Ephedra (Ephedraceae) and plays a significant role in desertification control. In this study, we assembled the complete chloroplast genome of E. rhytidosperma for the first time, revealing a typical quadripartite structure. The chloroplast genome was 109,545 base pair (bp) long with a GC content of 36.8%, encoding 118 complete genes. Phylogenetic reconstruction resolved E. rhytidosperma, E. minuta, E. likiangensis, E. equisetina, Ephedra gerardiana, E. fedtschenkoae, and E. monosperma formed a monophyletic group, indicating their close relationship. The chloroplast genome of E. rhytidosperma establishes a good foundation for further genetic and genomic studies of the Ephedra.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1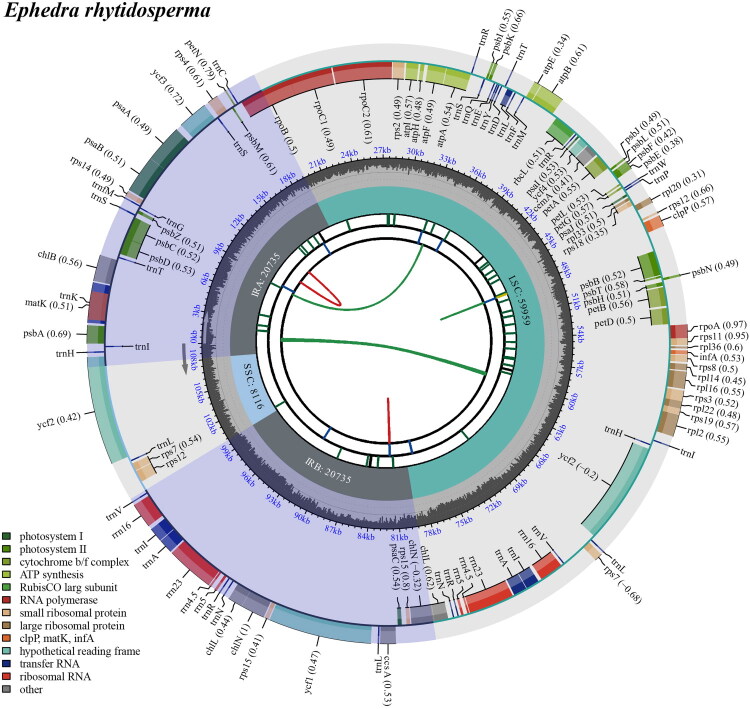 Figure 2
Figure 2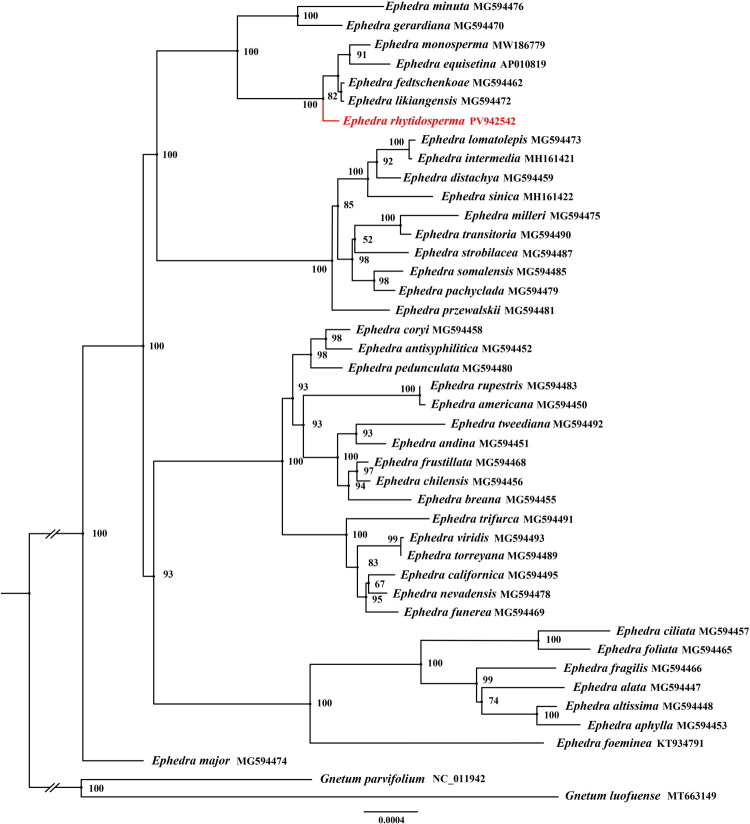 Figure 3
Figure 3- —Ningxia Innovative Training Program for College Students
- —Ningxia Natural Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Remote Sensing in Agriculture · Medicinal Plants and Bioactive Compounds
Introduction
Ephedra rhytidosperma Pachomova (Pachomova 1967), a typically dioecious small shrub belonging to the genus Ephedra (Ephedraceae), is distributed in Ningxia, where it thrives on hillsides and proluvial fan (Fu et al. 1999; Hollander et al. 2010; Shi et al. 2022). As an ecologically significant species, it plays a vital role in desertification control through its remarkable sand-binding capacity, attributed to its well-developed root system with powerful resistance to drought, cold, and sand burial (Han et al. 2022) It has also long been an important medicinal plant in China. Containing a plethora of chemical components, it can be used to treat a variety of diseases including cold, asthma, hay fever, and urticaria (Barker and Antia 2007; Mei et al. 2021). Despite these important attributes, the species remains understudied compared to other Ephedraceae members. In particular, the chloroplast genome of E. rhytidosperma remains unreported.
Chloroplasts are semi-autonomous organelles essential to plant cells (Sato et al. 1999; Hollingsworth et al. 2016). They possess their own genetic material, namely the chloroplast genome (Li et al. 2015; Daniell et al. 2016; Zhang et al. 2018; Ran et al. 2024). In most plants, chloroplast genome exhibits maternal inheritance and is composed of circular double-stranded DNA molecules ranging from 70 to 210 kb, contains approximately 130 genes (Hu et al. 2016; Guo et al. 2017; Yang et al. 2022). Due to its stable structure and conservative gene content, the chloroplast genome plays a vital role in studying plant origin (Yisilam et al. 2025), phylogenetic relationships (Kan et al. 2024; Yisilam et al. 2025), species identification (Quan et al. 2024), and classification (Gu et al. 2023), serving as a key tool for investigating evolutionary relationships among plant species (Zhang et al. 2012, 2018; Xie et al. 2022).
Here, we report the complete chloroplast genome of E. rhytidosperma, the first such assembly for this species. Our work characterizes its genomic structure and resolves the phylogenetic placement of E. rhytidosperma, offering valuable data to support taxonomy and evolutionary studies in Ephedra.
Materials
Healthy fresh stems of E. rhytidosperma were collected from Helan Mountain (Yinchuan, Ningxia, China; coordinates: 105.9058E, 38.9738N) (by Lei Zhang: [email protected]) (Figure 1), and desiccated using silica gel. In addition, collected the branches with flowers or fruits to prepare voucher specimens. The voucher specimen was archived in the Herbarium of North Minzu University (NMU: https://www.cvh.ac.cn/ins/info.php?code=NMU) with an accession number of NMU02465.
The photos of E. rhytidosperma. (A) Whole plant and flowers (photographed by Dr. Lei Zhang). (B) Herbarium of E. rhytidosperma (plant is short and nearly pad-like, leaves opposite, seeds 2, exceeding bracts, prominently longitudinally ridged).
Methods
Total genomic DNA was isolated with a modified CTAB method (Doyle and Doyle 1987). Sequencing libraries were prepared with the NEBNext DNA Library Kit, during which genomic DNA was fragmented into 350 base pair (bp). High-throughput sequencing was subsequently carried out on the Illumina NovaSeq 6000 platform (San Diego, CA) with 150 bp paired-end read length. We acquired 6.2 gigabases (Gb) of high-quality data after adapter trimming.
For chloroplast genome assembly, the de novo assembly was implemented in NOVOPlasty 4.3.3 (Dierckxsens et al. 2017) with the specified parameters: k-mer = 39 and genome range 120,000–200,000 bp, using the CPG of Ephedra intermedia (MG594471) as a reference seed. Genome annotation was executed through Plann v1.1 (Huang and Cronk 2015) with subsequent manual verification in Geneious v11.0.3 (Kearse et al. 2012). Sequencing depth analysis was quantified using Samtools (Li et al. 2009). The final assembly and annotation of E. rhytidosperma was submitted to GenBank (Benson et al. 2013). CPGView website (http://www.1kmpg.cn/cpgview/) was applied to visualize the gene map and the schematic maps of the cis- and trans-splicing genes (Liu et al. 2023).
To establish the phylogenetic placement of E. rhytidosperma in Ephedra, the chloroplast genomes of 42 representative species were retrieved from NCBI GenBank to reconstruct the chloroplast genome phylogenetic tree, with Gnetum luofuense and G. parvifolium serving as an outgroup. Protein-coding genes (PCGs) were extracted from the GenBank formatted file containing 43 plastomes using customized Perl scripts, removing start and end codons. A total of 60 PCGs were retained for all species. Each PCG was aligned using PRANK v.130410 (Löytynoja and Goldman 2008) according to the translated amino acid sequences. Phylogenetic reconstruction employed both maximum likelihood (ML) and Bayesian inference (BI) approaches: ML analysis was conducted in RAxML v8.1.24 (Stamatakis 2014) under the GTR + Γ model, while BI analysis in MrBayes v3.2.6 (Ronquist et al. 2012) utilized the GTR + I + G model selected through jModeltest. Final phylogenetic trees were visualized using FigTree v1.4.2 (Rambaut 2012). The results of the comparative analysis of the CPGs were visualized with the mVISTA program (Frazer et al. 2004) and the annotated CPG of E. intermedia was used as the reference in the LAGAN mode (Brudno et al. 2003).
Results
Following quality filtering and preprocessing, we obtained at least 4.2 Gb of whole-genome sequencing data were retained. These clean reads were used to assemble high-quality chloroplast genomes through a reference-guided approach. The total chloroplast genome of E. rhytidosperma (PV942542) was 109,545 bp long, with sequencing depth analysis revealed maximal, minimal, and average coverage values of 9321×, 27×, and 4478.58×, respectively (Fig. S1). The chloroplast genome exhibited a typical quadripartite structural organization, comprising a large single-copy (LSC) region, two inverted repeat (IR) regions, and a small single-copy (SSC) region (Figure 2 and Figure S2).
The detailed genome map of E. rhytidosperma cp genome. The species name is shown in the top left corner. The map contains six tracks in default. From the center outward, the first track shows the dispersed repeats. The dispersed repeats consist of direct and palindromic repeats, connected with red and green arcs. The second track shows the long tandem repeats as short blue bars. The third track shows the short tandem repeats or microsatellite sequences. The fourth track displays the genome length. The fifth track shows the GC content along the genome, while the sixth track sounds the genes. The gene names are followed by optional information about codon usage bias and color-coded based on their functional classification. The inner genes are transcribed clockwise, and the outer genes are transcribed anticlockwise. The functional type of the genes is shown in the bottom left corner.
The chloroplast genome of E. rhytidosperma annotation identified 118 complete genes (Table S1), comprising 73 PCGs, eight ribosomal RNA genes (rRNAs), and 37 tRNA genes (tRNAs). Additionally, the genome contained one trans-splicing gene (Fig. S3) and seven cis-splicing genes (Fig. S4), with an overall GC content of 36.8%. Phylogenetic reconstruction using both ML and BI methods robustly placed E. rhytidosperma within the Ephedra (Figure 3 and Figure S5). In these trees, the 41 Ephedra species dividing into five strongly supported clades (BS = 100%; PP = 1) (Figure 3 and Figure S5). Notably, E. rhytidosperma clustered with E. minuta, Ephedra gerardiana, E. monosperma, E. equisetina, E. fedtschenkoae, and E. likiangensis (BS = 100%; PP = 1), indicating a close evolutionary relationship between these species.
Phylogenetic tree obtained using the maximum likelihood (ML) and Bayesian inference (BI) methods of Ephedra species based on based on 60 PCGs. GenBank accession numbers: Ephedra minuta MG594476 (Yang et al. 2024), Ephedra gerardiana MG594470 (Han et al. 2022), Ephedra monosperma MW186779 (Zhao et al. 2021), Ephedra equisetina AP010819 (Wu et al. 2009), Ephedra likiangensis MG594472 (Rydin et al. 2021), Ephedra fedtschenkoae MG594462 (Rydin et al. 2021), Ephedra lomatolepis MG594473 (Yang et al. 2024), Ephedra intermedia MH161421 (Chen et al. 2019), Ephedra distachya MG594459 (Rydin et al. 2021), Ephedra sinica MH161422 (Li et al. 2019), Ephedra milleri MG594475 (Rydin et al. 2021), Ephedra transitoria MG594490 (Rydin et al. 2021), Ephedra strobilacea MG594487 (Rydin et al. 2021), Ephedra somalensis MG594485 (Zhao et al. 2021), Ephedra pachyclada MG594479 (Yang et al. 2024), Ephedra przewalskii MG594481 (Rydin et al. 2021), Ephedra coryi MG594458 (Rydin et al. 2021), Ephedra antisyphilitica MG594452 (Rydin et al. 2021), Ephedra pedunculata MG594480 (Rydin et al. 2021), Ephedra rupestris MG594483 (Rydin et al. 2021), Ephedra americana MG594450 (Rydin et al. 2021), Ephedra tweediana MG594492 (Rydin et al. 2021), Ephedra andina MG594451 (Rydin et al. 2021), Ephedra frustillata MG594468 (Rydin et al. 2021), Ephedra chilensis MG594456 (Rydin et al. 2021), Ephedra breana MG594455 (Rydin et al. 2021), Ephedra trifurca MG594491 (Rydin et al. 2021), Ephedra viridis MG594493 (Rydin et al. 2021), Ephedra torreyana MG594489 (Rydin et al. 2021), Ephedra californica MG594495 (Rydin et al. 2021), Ephedra nevadensis MG594478 (Rydin et al. 2021), Ephedra funerea MG594469 (Rydin et al. 2021), Ephedra ciliata MG594457 (Rydin et al. 2021), Ephedra foliata MG594465 (Rydin et al. 2021), Ephedra fragilis MG594466 (Rydin et al. 2021), Ephedra alata MG594447 (Rydin et al. 2021), Ephedra altissima MG594448 (Rydin et al. 2021), Ephedra aphylla MG594453 (Rydin et al. 2021), Ephedra foeminea KT934791 (Li et al. 2019), Ephedra major MG594474 (Rydin et al. 2021), Gnetum parvifolium NC_011942 (Chang et al. 2020), Gnetum luofuense MT663149 (Yang et al. 2024), and Ephedra rhytidosperma PV942542.
Discussion and conclusions
In this study, we assembled the CPG of E. rhytidosperma for the first time, revealing a typical quadripartite structure consisting of a pair of IRs, a LSC region, and a SSC region. The assembled genome has a total length of 109,545 bp, contains 118 complete genes, and exhibits a GC content of 36.8%. These characteristics are consistent with those reported for other Ephedra species (Chen et al. 2019; Han et al. 2022).
The cp genomes have become a central focus in molecular biology research due to their significant potential for resolving phylogenetic relationships among higher plants. In this study, phylogenetic analyses using BI and ML methods revealed that Ephedra cp genomes form five strongly supported main clades (BS = 100%; PP = 1), a result consistent with previous studies (Chen et al. 2019; Zhao et al. 2021). Notably, E. rhytidosperma, E. minuta, E. gerardiana, E. monosperma, E. equisetina, E. fedtschenkoae, and E. likiangensis form a distinct monophyletic group. The sequence alignment of the CPGs of the above seven Ephedra species indicated high sequence similarity among the CPGs of the seven Ephedra species (Fig. S6). However, sequence divergence in non-coding regions was greater than that in coding regions. The faster evolutionary rate in non-coding regions provides abundant raw material for natural selection. To further advance our understanding of E. rhytidosperma, future studies should prioritize a comprehensive analysis of its codon usage bias and population-level genomic diversity. Such investigations are key to elucidating structural variations in its chloroplast genome. Moreover, expanding the repertoire of complete chloroplast genomes across the Ephedra genus will be crucial for clarifying the evolutionary dynamics of this ecologically significant group.
Supplementary Material
Supplementary File.docx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barker WD, Antia U. 2007. A study of the use of Ephedra in the manufacture of methamphetamine. Forensic Sci Int. 166(2–3):102–109. 10.1016/j.forsciint.2006.04.00516707238 · doi ↗ · pubmed ↗
- 2Benson DA et al. 2013. Gen Bank. Nucleic Acids Res. 41:D 36–D 42. 10.1093/nar/gks 119523193287 PMC 3531190 · doi ↗ · pubmed ↗
- 3Brudno M et al. 2003. Glocal alignment: finding rearrangements during alignment. Bioinformatics. 19(Suppl. 1):i 54–i 62. 10.1093/bioinformatics/btg 100512855437 · doi ↗ · pubmed ↗
- 4Chang AC, Bautista MA, Zheng Y, Wan T, Chen T. 2020. Plastome structure and phylogeny of Gnetum luofuense CY Cheng (Gnetaceae, Gnetales). Mitochondrial DNA B Resour. 5(3):3369–3370. 10.1080/23802359.2020.182181833458172 PMC 7783052 · doi ↗ · pubmed ↗
- 5Chen XL et al. 2019. Identification and phylogenetic analysis of the complete chloroplast genomes of three Ephedra herbs containing Ephedrine. Biomed Res Int. 2019:5921725. 10.1155/2019/592172530941367 PMC 6420972 · doi ↗ · pubmed ↗
- 6Daniell H, Lin CS, Yu M, Chang WJ. 2016. Chloroplast genomes: diversity, evolution, and applications in genetic engineering. Genome Biol. 17(1):134. 10.1186/s 13059-016-1004-227339192 PMC 4918201 · doi ↗ · pubmed ↗
- 7Dierckxsens N, Mardulyn P, Smits G. 2017. NOVO Plasty: de novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 45(4):e 18. 10.1093/nar/gkw 95528204566 PMC 5389512 · doi ↗ · pubmed ↗
- 8Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull. 19:11–15.