Research progress and strategies for vaccines and targeted drugs against highly virulent porcine epidemic diarrhea virus
Qiuxuan Wang, Songrui Liu, Hongrui Chen, Xuanyi Liu, Hanjia Zhang, Xuehui Wang, Qingfeng Meng, Hao Dong

TL;DR
This paper reviews progress in vaccines and drugs to combat a deadly pig disease caused by a highly virulent virus.
Contribution
The paper provides new strategies combining vaccines and targeted drugs to improve protection against highly virulent PEDV.
Findings
Current commercial vaccines offer only partial protection against new PEDV strains.
Combining natural compounds and targeted drugs can enhance prevention effectiveness.
Drugs targeting TLR3 can help compensate for interferon deficiencies in vaccines.
Abstract
As an acute and highly contagious enteric disease of swine, porcine epidemic diarrhea virus (PEDV) has caused high piglet mortality and significant economic losses. Commercialized vaccines provide only partial cross-protection against the novel, highly virulent PEDV strains. Developing new vaccines against highly virulent PEDV strains would help protect the pig industry from the serious challenges posed by novel, highly virulent PEDV infections. Natural compounds and chemical and biochemical source-targeted drugs designed to act on specific proteins, enzymes, or mechanisms can complement each other’s advantages when used in combination, thereby enhancing the effectiveness of drug-based prevention in the control of highly virulent PEDV. Drugs targeting Toll-like receptor 3 (TLR3) can aid vaccines to compensate for interferon (IFN) secretory deficiencies to protect pigs from highly…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
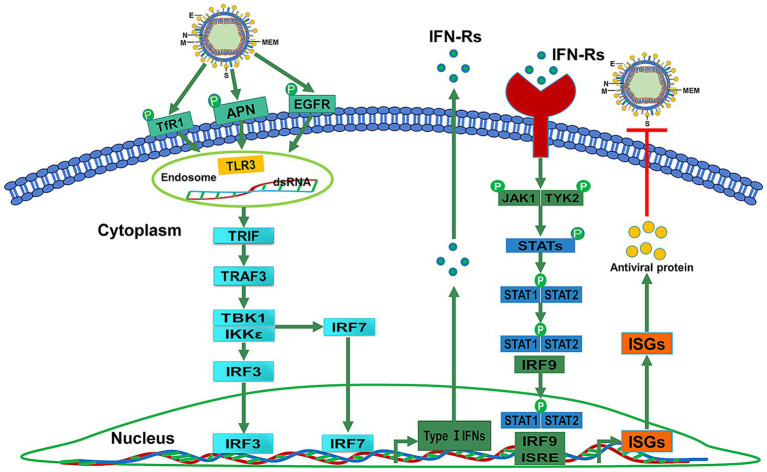 Figure 1
Figure 1| Vaccine | Immune induction | Protection efficiency | References |
|---|---|---|---|
| PEDV-S mRNA-LNP vaccine | PEDV-specific humoral and cellular immune responses. | Protected immunized piglets against the PEDV AH2012/12 strain | |
| Trimeric full-length S protein subunit vaccine | High levels of S-specific IgG, IgA, and neutralizing antibodies; increased expression levels of IFN-γ and IL-4. | Reduced intestinal pathological damage in immunized piglets infected with AH2012/12 |
|
| Recombinant rAJ1102-S2′JS2008 Vaccine | Induced neutralizing antibodies | Protected pigs from G1 and G2 PEDV infections. | |
| PLGA-KAg | Improved lymphocyte proliferation responses, IFN-γ levels, and PEDV-specific IgG and IgA antibody titers | Provided protective immunity against PEDV AH2012/12 strain infection in suckling piglets. |
|
| A flagellin -adjuvanted inactivated porcine epidemic diarrhea virus (PEDV) vaccine | Elicited high levels of IgG, IgA, and neutralizing antibodies | Conferred protective immunity to piglets against PEDV strain AH2012/12 infection. |
|
| PEDV VLPs with CCL25/28 | Modulated the immune responses by enhancing systemic anti-PEDV S-specific IgG, mucosal IgA, and cell immunity | Alleviated clinical signs in piglets infected with PEDVPT-P7. |
| Natural compounds | Inhibit PEDV in cells | Effective in pigs | Target | References |
|---|---|---|---|---|
| Flavonol | PEDV-YN145 | No report | Interacts with PEDV Mpro. |
|
| Tomatidine | PEDV MS, YZ, SH, and CV777 | No report | Inhibition of 3CLpro activity. |
|
| Wogonin | PEDV AH2012/12 | No report | Exerts an inhibitory effect on Mpro. |
|
| Licorice extract | PEDV HM2017 | Against PEDV HM2017 infection in piglets. | 1. Interfering with the PEDV N protein, ORF3 mRNA, and M protein |
|
| Buddlejasaponin IVb | PEDV AH-2018-HF1 | Relieving clinical symptoms in pigs. | Inhibits the NF-κB signaling pathway. |
|
| Hypericum japonicum extract | PEDV-CV777 and PEDV-G2 | Improving intestinal pathology in piglets | Interfering with interaction between the N protein and p53. |
|
| Puerarin | PEDV Yunnan province strain | Reduction of PEDV impact on growth performance in piglets. | Regulating the IFN and NF-κB signaling pathways. |
|
| Aloe extract | PEDV GDS01 | Protects newborn piglets from PEDV variant GDS01 infection | Blocking the transcription of viral N genes. |
|
| Drug | Inhibit PEDV in cells | Effective in pigs | Target | References |
|---|---|---|---|---|
| Niclosamide | CV777, HNXX, HB, HW, and recombinant PEDV strains DR13-GFP and DR13-Rlu | No report | Targeting STAT3. |
|
| Tubercidin | CV777, HNAY, HNXX, HB, DR13-GFP, and GDU-GFP | No report | Forms hydrophobic interactions with the RdRp of PEDV. |
|
| Ivermectin | CV777, HW, HNXX, HB (GII subtype), and PEDV (DR13-GFP) | No report | Induces cell cycle arrest to inhibit viral release. |
|
| PA-824 | PEDV AH-2018-HF | Reducing the pathogenic effect of PEDV in piglets | Suppressing PEDV-induced p53 activation. |
|
| Drug | Inhibit PEDV in cells | Effective in pigs | Target | References |
|---|---|---|---|---|
| RNA G-quadruplexes | CV777, HNAY, HW, and DR13-GFP strains | No report | Targeting the G4 structure in Nsp5. |
|
| KPNA2 | PEDV strain GDS01 | No report | Degrading the viral E protein by autophagy |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Virus Infections Studies · Viral gastroenteritis research and epidemiology · Virus-based gene therapy research
Introduction
1
As an acute and highly contagious enteric disease of pigs, porcine epidemic diarrhea (PED) can result in dehydration, vomiting, diarrhea, and severe enteritis. Its lethality is particularly high in suckling pigs (Stadler et al., 2015). Porcine epidemic diarrhea virus (PEDV), a causative agent belonging to the genus Alphacoronavirus in the family Coronaviridae, is an enveloped, positive, single-stranded RNA virus (Karte et al., 2020). The PEDV genome is approximately 28 kb and contains at least seven open reading frames (ORFs), which encode two large polyprotein precursors (pp1a and pp1ab); the spike (S), membrane (M), and envelope (E) structural proteins; and nucleocapsid (N) proteins, as well as an accessory protein, ORF3 (Zhuang et al., 2025). The virus genotype includes G1 (classical G1a and recombinant G1b) and G2 (local epidemic G2a and global epidemic G2b) (Jang et al., 2023).
The highly pathogenic (HP)-G2b PEDV caused a pandemic that severely impacted pig-producing nations in America and Asia during 2013–2014 and also threatened the global pig industry (Park et al., 2007; Puranaveja et al., 2009; Lee et al., 2024). Most PEDV strains isolated from Vietnam belonged to the genotypes G1 and G2 and had very high genetic similarity with strains isolated from China and Thailand (Nguyen et al., 2023). The virus was first recognized in Europe in the 1970s and caused high piglet mortality and significant economic losses in Germany, France, Belgium, Ukraine, Austria, Portugal, and the Netherlands in 2014 (Dastjerdi et al., 2015; Grasland et al., 2015; Mesquita et al., 2015; Stadler et al., 2015; Steinrigl et al., 2015; Theuns et al., 2015; Dortmans et al., 2018). It was reported that the prevalence of PEDV-positive piglets during the first week on Spanish farms ranged from 3.7 to 12.9% in 2014 (Mesonero-Escuredo et al., 2018; Vidal et al., 2019). A recent investigation of 106 Spanish pig farms between 2017 and 2019 showed that the detected PEDV rate was 38.7% (Monteagudo et al., 2022). The investigation showed that PED can rapidly spread in PEDV-negative herds and cause 100% morbidity and 30 to 90% mortality in piglets (Jang et al., 2023). PEDV can also cause a 12.6% reduction in the farrowing rate and result in a 5.7% failure-to-breed rate, a 1.3% abortion rate, and 2.0% mummified fetuses, negatively affecting the reproductive performance of mature sows (Weng et al., 2016).
To fight the novel highly virulent PEDV infection, this review summarizes recent progress in the development of vaccines against highly virulent PEDV, natural compounds, and chemical and biochemical source-targeted drugs that have been explored in cell and pig models with clearly defined mechanisms. It also aims to provide comprehensive strategies for the prevention and control of highly virulent PEDV infections in pigs.
Progress and strategies in vaccines against highly virulent PEDV
2
Viral entry, attachment, induction of neutralizing antibodies, and membrane fusion are mediated by the S1and S2 domains of the PEDV S glycoprotein. The CO-26 K-equivalent (COE) and N-terminal domain (NTD) in the S1 region are crucial neutralizing epitopes and potential co-receptor binding sites for the vaccine development of PEDV (Kirchdoerfer et al., 2021). Lipid nanoparticle (LNP)-encapsulated mRNA (mRNA-LNP) vaccines encoding a PEDV multiepitope chimeric spike (Sm) protein (PEDV-S mRNA-LNP) have been demonstrated to activate CD4 + and CD8 + T cells and induce PEDV-specific IgG and IgA in the serum and colostrum of S-mRNA-immunized sows, which could be transferred to suckling neonatal piglets, providing protection against AH2012/12 infection (Kirchdoerfer et al., 2021; Zhao et al., 2024).
Whole-virus vaccines in traditional PEDV vaccines include inactivated and attenuated vaccines. In contrast to traditional PEDV vaccines, subunit vaccines can provide safety, without viral nucleic acids, the redesigned antigens and multiple antigens combination with the adjuvant addition in immunity efficacy elevation (Du et al., 2016). A complete subunit vaccine production system would greatly facilitate a quick response to emergency epidemics (Li Z. et al., 2020). The study showed that the full-length S protein subunit vaccine could effectively induce high levels of S-specific IgG, IgA, and neutralizing antibodies in pigs infected with AH2012/12. It also increased the proliferation of peripheral blood mononuclear cells and increased interferon-γ (IFN-γ) and interleukin-4 (IL-4) expression levels in peripheral blood to reduce diarrheal index scores, fecal viral loads, and intestinal pathological damage in immunized piglets (Guo et al., 2024).
The addition of trypsin is crucial but also increases the complexity of vaccine production and cost in the propagation of PEDV. It has been reported that PEDV trypsin independence is associated with the S2′ site and Y976/977 of the PEDV spike (S) protein (Li M. et al., 2023). Li M et al. used AJ1102 and the trypsin-independent genotype 1 (G1) PEDV strain JS2008 to generate a recombinant PEDV carrying a chimeric S protein and successfully constructed the trypsin-independent PEDV strain rAJ1102-S2′JS2008 (Li M. et al., 2023). It was able to effectively replicate in the absence of trypsin and could induce neutralizing antibodies against AJ1102 and JS2008, providing protection to pigs against G1 and G2 PEDV infections (Niu and Wang, 2022; Li M. et al., 2023).
Immunizing sows with PEDV vaccines between 20 and 30 days will provide substantial passive immunity to their newborn piglets, especially mucosal immunity, which is essential for the sows (Lin et al., 2016). As a particle-mediated delivery system for vaccines, biodegradable and biocompatible poly (D, L-lactide-co-glycolic acid) (PLGA) nanoparticles (NPs) can protect the entrapped vaccine from protease-mediated degradation at mucosal surfaces and stimulate the underlying mucosal immune cells to provide protection for sows infected with AH2012/12 (Binjawadagi et al., 2014a; Binjawadagi et al., 2014b). PLGA nanoparticle-entrapped PEDV killed vaccine antigens (KAg) (PLGA-KAg) have been shown to improve PEDV-specific IgG and IgA antibody titers, induce lymphocyte proliferation responses, and increase IFN-γ levels in pregnant sows and their suckling piglets (Li et al., 2017).
As a potential adjuvant, flagellin can induce Th1 and Th2 mixed-cell responses (Li et al., 2018). Flagellin can be used in combination with inactivated or killed PEDV vaccines to elevate mucosal and systemic IgG and IgA levels, thereby protecting piglets from PEDV AH2012/12 infection (Xu X. et al., 2020).
For the highly virulent PEDV G2 strains, traditional vaccines can only provide partial cross-protection (Wang et al., 2016). Commercialized vaccines, including recombinant PEDV S protein, an inactivated whole-virus vaccine based on a non-S INDEL PEDV strain, and a subunit vaccine using HEK-293 T cell-expressed PEDV S1 proteins, have been used to control virulent G2 viruses in the United States (Makadiya et al., 2016). However, commercialized vaccines cannot provide consistency in stimulating solid lactogenic immunity to protect suckling piglets from G2 virus infection (Crawford et al., 2016). Virus-like particles (VLPs) can improve immunogenicity, drain freely into lymph nodes, and be efficiently taken up by antigen-presenting cells to promote CD4 + T helper cell and CD8 + cytotoxic T cell responses (Dudziak et al., 2007; Mohsen et al., 2017). As characterized nanoparticles of conformational epitopes, VLPs can induce the subsequent humoral immunity by interacting with B cells (Manolova et al., 2008; Hsueh et al., 2020). In the development of safe, effective, and economical vaccines against enteric viral diseases, VLP vaccines represent an important strategy by stimulating cellular, mucosal, and humoral immunity. In the current study, PEDV VLPs of CCL25/28 were demonstrated to protect pigs from PEDVPT-P7infection by increasing systemic anti-PEDV S-specific IgG, mucosal IgA, and cellular immunity (Leidenberger et al., 2017; Hsu et al., 2020; Lu et al., 2020).
mRNA-LNP vaccines, the full-length S protein subunit, the trypsin-independent genotype 1 (G1) PEDV JS2008 strain, PLGA nanoparticle-entrapped PEDV killed vaccines, flagellin, and PEDV VLPs of CCL25/28 have shown different immune regulation efficiencies in enhancing systemic anti-PEDV S-specific IgG, mucosal IgA, and cell immunity to protect pigs from highly virulent PEDV infection (Table 1). To continue exploring vaccines, it is indispensable to prevent and control PED infections caused by different novel highly virulent PEDV strains in pigs.
Progress and strategies in drugs targeting highly virulent PEDV
3
Maternal antibodies from colostrum and milk are important to protect piglets from PEDV infection (Leidenberger et al., 2017). PEDV mutation could decrease the full protection of the vaccines (Li S. et al., 2020). Therefore, it is necessary to update vaccines based on prevalent PEDV strains and explore new strategies (Yang et al., 2023). Antiviral natural compounds from plant extracts and Chinese herbal medicines have been increasingly demonstrated in recent years. In view of rich sources, unique chemical structures, and diverse activities of natural compounds in the development of new anti-highly virulent PEDV drugs, natural compounds will compensate for the vaccine deficiency in against PEDV prevalent strains (Russo et al., 2020; Gong et al., 2023). Recently, the anti-highly virulent PEDV of natural products target drugs have become a hot spot because of its lower side effects, cheaper investment and avoidable risk in developing resistance (Behzadi et al., 2023; Liang et al., 2024). Many natural compounds have also been reported to be effective in inhibiting highly virulent PEDV (Sun et al., 2022). Drugs including flavonol, tomatidine, and wogonin have been reported to affect highly virulent PEDV by interacting with the Mpro or 3CLpro proteins of PEDV in vitro (Wang et al., 2020; Wang J. et al., 2023; Liang et al., 2024) (Table 2). These compounds can be good candidate drugs against highly virulent PEDV in cells or pigs, pending further demonstration in in vivo studies. Based on their effects on highly virulent PEDV in vitro and vivo, licorice extract, buddlejasaponin IVb, hypericum japonicum extract, puerarin, and aloe extract have been shown to inhibit highly virulent PEDV by interfering with the N protein, ORF3 mRNA, and M protein; inhibiting the PI3K and NF-κB signaling pathways; and blocking the transcription of viral N genes (Wu et al., 2020; Xu Z. et al., 2020; Su et al., 2021; Sun et al., 2022; Rao et al., 2023; Yang et al., 2023). These drugs could reduce the replication of highly virulent PEDV and also alleviate clinical symptoms in pigs. They hold promising clinical value for future exploration of their effects against highly virulent PEDV both in vitro and in vivo.
There are chemical drugs targeting highly virulent PEDV, including niclosamide, tubercidin, and ivermectin, that can inhibit the proliferation of highly virulent PEDV in vitro by targeting the specific viral mechanisms (Wang Y. et al., 2023; Wang et al., 2024; Xu et al., 2024) (Table 3). Considering the evasive strategies of PEDV, it is important to regulate the proliferation of highly virulent PEDV by targeting the signal transducer and activator of transcription 3 (STAT3) and RNA-dependent RNA polymerase (RdRp) (Wang Y. et al., 2023; Wang et al., 2024; Xu et al., 2024). Although these targeted drugs have only been tested in vitro, they still offer extraordinary therapy strategies for the prevention of highly virulent PEDV. Among these drugs, PA-824 has been demonstrated to inhibit the proliferation of highly virulent PEDV and alleviate diarrhea symptoms in pigs caused by PEDV AH-2018-HF infection by suppressing PEDV-induced p53 activation in vitro and in vivo (Li et al., 2024). Especially, the specially target drugs tested in pigs will be priority in synergistic therapy and increase anti-highly virulent PEDV efficiency.
Biochemical source drugs, including RNA G-quadruplexes and Karyopherin α2 (KPNA2), have shown substantial inhibition of highly virulent PEDV replication by targeting the G4 structure in Nsp5 and the E protein, respectively, in vitro (Gao et al., 2023; Li Y. et al., 2023). The highly virulent PEDV genome and structural proteins (S, E, M, and N) (Table 4) are crucial determinants of the molecular epidemiological characteristics of PEDV (Karte et al., 2020; Jang et al., 2023; Zhuang et al., 2025). It is necessary to further explore and test biochemical source drugs in vivo, as they may provide new options to face emerging challenges from PEDV variant strains. As an important direction for future studies, there is a real demand in veterinary clinics to explore and screen high-efficiency, low-toxicity, and low-residue drugs with targeted therapy against highly virulent PEDV (Behzadi et al., 2023). Natural compounds and chemical and biochemical source-targeted drugs can complement each other’s advantages through drug combination, thereby promoting the efficacy of drug-based prevention and control of highly virulent PEDV.
Progress and strategies in drugs targeting toll-like receptor 3 (TLR3)
4
Inducing antiviral innate immune and inflammatory responses is an important precondition for repelling viral infections (Yang and Shu, 2020). Studies have shown that the production of type I or type III IFNs could be inhibited by highly virulent PEDV N proteins, such as nsp1, PLP2, nsp5, nsp15, and nsp16 (Deng et al., 2019; Shi et al., 2019). This inhibition benefits highly virulent PEDV by enabling immune evasion through suppression of IFN production pathways and disruption of transcription factor activation involved in IFN induction (Cao et al., 2015a; Guo et al., 2016; Li S. et al., 2020). Pattern recognition receptors (PRRs) can specifically recognize pathogen-associated molecular patterns (PAMPs) by activating IFN- and interleukin-1 (IL-1)-mediated proinflammatory responses in animals (Rai, 2020). As a member of the virus-perceiving PRRs, Toll-like receptor 3 (TLR3) can initiate downstream signal transduction, upregulate the IFN-α/β expression, and induce antiviral protein (AVP) synthesis activity by recognizing viral double-stranded RNA (dsRNA) (Unterholzner et al., 2010; Matsumoto et al., 2011). Within the TLR family, TLR3 is the only receptor that induces IFN-β production through the Toll/IL-1 receptor (TIR) domain-containing adapter-inducing interferon-β (TRIF) pathway (Yang and Shu, 2020). The TRIF-dependent nuclear factor-κB (NF-κB) and IFN regulatory factor 3/7 (IRF3/7) pathways are regulated by TLR3 (Matsumoto et al., 2011). When TLR3 is activated by viral dsRNA, TRIF could elicit a cascade of reactions by triggering tumor necrosis factor (TNF) receptor-associated factor 3 (TRAF3) and TRAF6 (Fang et al., 2013; Bugge et al., 2017) (Figure 1).
TLR3 upregulates IFN-α/β expression and induces AVP synthesis activity by recognizing dsRNA. The TLR3-mediated signaling pathway includes the TRIF-dependent NF-κB and IRF3/7 pathways. TRIF interacts with TRAF3 to elicit a cascade of reactions to induce the production of IFNs-1.
TLR3 localizes to endosomes and the cell surface in macrophages and mast cells (MCs) but is restricted to endosomes in myeloid dendritic cells (DCs) (Matsumoto et al., 2003; Matsumoto et al., 2011; Agier et al., 2016). Type I IFNs (α and β) are associated with viral clearance and can be produced by DCs (Matsumoto et al., 2003). TLR3 can drive antigen-presenting DCs to induce IFN production (Soto et al., 2020). TLR3 also promotes IRF3, type I and II IFN receptor, and major histocompatibility complex (MHC) I expression in MCs to enhance the cellular antiviral response (Soto et al., 2020; Witczak et al., 2020). Viral infection leads to TLR3 expression upregulation in DCs of mice and humans. Virus dsRNA is recognized by the TLR3 ectodomain (ECD) (Negishi et al., 2008; Abe et al., 2012). The TIR domain of TLR3 can recruit TRIF (Chattopadhyay and Sen, 2014). It can stimulate the phosphorylation of IRF3, which leads to the production of type I IFNs (Takeda and Akira, 2004). A study demonstrated that TLR3 positively contributes to NF-kB activation in response to PEDV infection (Cao et al., 2015b). TLR3 activates NF-κB signaling through TRIF-dependent conscription of two cascades. It is most noteworthy pathway of TLR3 signaling that can provoke TNF, IL-1, CCL2, CXCL8, endothelial adhesion molecules, and type I IFNs to against viruses (Komal et al., 2021). The TLR3 agonist can upregulate the expression of IFN-α/β and induce AVP synthesis activity by recognizing virus dsRNA to activate TLR3 downstream signal transduction (Unterholzner et al., 2010; Matsumoto et al., 2011).
Novel TLR3 agonists include RGC100, Poly-IC, and ARNAX. As a novel TLR3 agonist, RGC100 can target endosomal TLR3 and activate murine myeloid DCs to promote proinflammatory cytokine secretion in a dose-dependent manner (Naumann et al., 2013). Considering its immunological properties, RGC100 may represent a promising candidate for prevention and therapy vaccination strategies against PEDV. As a synthetic dsRNA analog, polycytidylic acid (PolyI:C) can be recognized by RIG-I/MDA-5 and TLR3 receptors to activate transcription factors that are responsible for the expression of type I IFNs and inflammatory cytokines/chemokines (Kanmani and Kim, 2019). Poly-IC12U is an altered form of poly IC. It reduces poly IC-associated toxicity and regulates IFN expression by activating the TLR3 receptor (Martins et al., 2015). As a synthetically derived form of poly-IC, Poly-ICLC can induce strong Th1 cytokine responses, including IL-6, IL-12, TNF-α, IFN-γ, and type 1 IFNs (Komal et al., 2021). ARNAX is a synthetic DNA–dsRNA hybrid compound and can activate MDA5 (Komal et al., 2021). ARNAX cannot activate the TLR3 pathway.
Different from ARNAX, poly-IC can activate both TLR3 and MDA5 (Shime et al., 2017). In addition, the inflammatory status of macrophages and DCs can also be changed by poly-IC and RGC100 (Longhi et al., 2009; Gupta et al., 2016; Takeda et al., 2018). Therefore, TLR3 agonists, poly-IC and RGC100, ought to be considered as adjuvants for highly virulent PEDV vaccination.
Conclusion
5
As an acute and highly contagious enteric disease of swine, highly virulent PEDV causes high piglet mortality and significant economic losses. However, commercialized vaccines can only provide partial cross-protection against novel highly virulent PEDV strains. The development of new vaccines against highly virulent PEDV including mRNA-LNP, subunit, trypsin-independent, nanoparticle-entrapped killed PEDV, and virus-like particle (VLP) vaccines will help protect the swine industry from the serious challenges posed by highly virulent PEDV infection. Natural compounds and chemical and biochemical source-targeted drugs can enhance the effectiveness of drug-based prevention in controlling highly virulent PEDV. As adjuvants, TLR3 agonists can aid vaccines to compensate for IFN secretory deficiencies to protect pigs from highly virulent PEDV infection. Researchers working on the vaccines and drugs mentioned in this review need more time to complete in-depth studies on vaccines and targeted drugs against highly virulent porcine epidemic diarrhea virus. Continued focus on the ongoing research of these vaccines and drugs will provide valuable scientific information for their application in PEDV control and prevention, once sufficient evidence supports effective strategies for managing the disease.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abe Y.Fujii K.Nagata N.Takeuchi O.Akira S.Oshiumi H.. (2012). The toll-like receptor 3-mediated antiviral response is important for protection against poliovirus infection in poliovirus receptor transgenic mice. J. Virol. 86, 185–194. doi: 10.1128/jvi.05245-11, PMID: 22072781 PMC 3255933 · doi ↗ · pubmed ↗
- 2Agier J.Żelechowska P.Kozłowska E.Brzezińska-Błaszczyk E. (2016). Expression of surface and intracellular toll-like receptors by mature mast cells. Cent Eur J Immunol 4, 333–338. doi: 10.5114/ceji.2016.65131, PMID: 28450795 PMC 5382879 · doi ↗ · pubmed ↗
- 3Behzadi A.Imani S.Deravi N.Mohammad Taheri Z.Mohammadian F.Moraveji Z.. (2023). Antiviral potential of Melissa officinalis L.: a literature review. Nutr. Metab. Insights 16:11786388221146683. doi: 10.1177/11786388221146683, PMID: 36655201 PMC 9841880 · doi ↗ · pubmed ↗
- 4Binjawadagi B.Dwivedi V.Manickam C.Ouyang K.Torrelles J. B.Renukaradhya G. J. (2014 a). An innovative approach to induce cross-protective immunity against porcine reproductive and respiratory syndrome virus in the lungs of pigs through adjuvanted nanotechnology-based vaccination. Int. J. Nanomedicine 9, 1519–1535. doi: 10.2147/ijn.S 59924, PMID: 24711701 PMC 3969340 · doi ↗ · pubmed ↗
- 5Binjawadagi B.Dwivedi V.Manickam C.Ouyang K.Wu Y.Lee L. J.. (2014 b). Adjuvanted poly(lactic-co-glycolic) acid nanoparticle-entrapped inactivated porcine reproductive and respiratory syndrome virus vaccine elicits cross-protective immune response in pigs. Int. J. Nanomedicine 9, 679–694. doi: 10.2147/ijn.S 5612724493925 PMC 3908835 · doi ↗ · pubmed ↗
- 6Bugge M.Bergstrom B.Eide O. K.Solli H.Kjønstad I. F.Stenvik J.. (2017). Surface toll-like receptor 3 expression in metastatic intestinal epithelial cells induces inflammatory cytokine production and promotes invasiveness. J. Biol. Chem. 292, 15408–15425. doi: 10.1074/jbc.M 117.784090, PMID: 28717003 PMC 5602399 · doi ↗ · pubmed ↗
- 7Cao L.Ge X.Gao Y.Herrler G.Ren Y.Ren X.. (2015 a). Porcine epidemic diarrhea virus inhibits ds RNA-induced interferon-β production in porcine intestinal epithelial cells by blockade of the RIG-I-mediated pathway. Virol. J. 12:127. doi: 10.1186/s 12985-015-0345-x, PMID: 26283628 PMC 4539884 · doi ↗ · pubmed ↗
- 8Cao L.Ge X.Gao Y.Ren Y.Ren X.Li G. (2015 b). Porcine epidemic diarrhea virus infection induces NF-κB activation through the TLR 2, TLR 3 and TLR 9 pathways in porcine intestinal epithelial cells. J. Gen. Virol. 96, 1757–1767. doi: 10.1099/vir.0.000133, PMID: 25814121 · doi ↗ · pubmed ↗