B cells inhibit IL-1 family cytokine production and Mycobacterium tuberculosis growth in human CD14+ cells
Abhinav Vankayalapati, Bharath Somasundram, Padmaja Paidipally, Karan P Singh, Ramakrishna Vankayalapati, Rajesh Kumar Radhakrishnan

TL;DR
B cells reduce inflammation and inhibit the growth of Mycobacterium tuberculosis in human monocytes.
Contribution
This study reveals a novel regulatory role of B cells in modulating cytokine production and Mtb growth in human macrophages.
Findings
B cells inhibit proinflammatory cytokines like IL-1α, IL-1β, and TNF-α in human CD14+ monocytes.
B cells enhance anti-inflammatory cytokines such as IL-1Ra and IL-10 and inhibit Mtb growth.
Neutralizing antibodies against IL-1α, IL-1β, and IL-18 increase Mtb growth, while antibodies against IL-1Ra reduce it.
Abstract
The IL-1 family of cytokines produced by antigen-presenting cells plays important roles in various diseases and infections, including Mycobacterium tuberculosis (Mtb) infection. In the present study, we infected human monocyte-derived macrophages (MDMs) with Mtb. Then, we measured the production of IL-1 superfamily (ILSF) cytokines (8 soluble factors) and determined the effects of ILSF cytokines on Mtb growth via the use of recombinant cytokines and antibodies. Mtb infection significantly increased the production of IL-1α, IL-1β, IL-18, and IL-37 and reduced the production of IL-1Ra by MDMs. Human recombinant IL-1α, IL-1β, and IL-18 reduced Mtb growth in MDMs. In contrast, human recombinant IL-1Ra enhanced Mtb growth in MDMs. Neutralizing antibodies against IL-1α, IL-1β, and IL-18 enhanced Mtb growth, and neutralizing antibodies against IL-1Ra and IL-33R reduced Mtb growth in MDMs. B…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
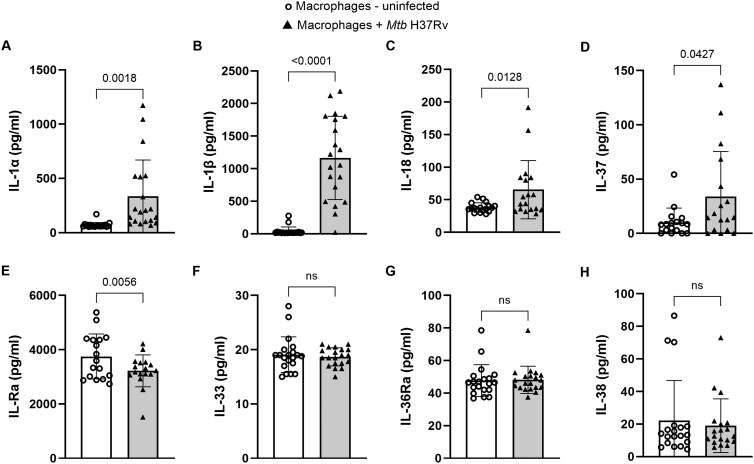 Figure 1
Figure 1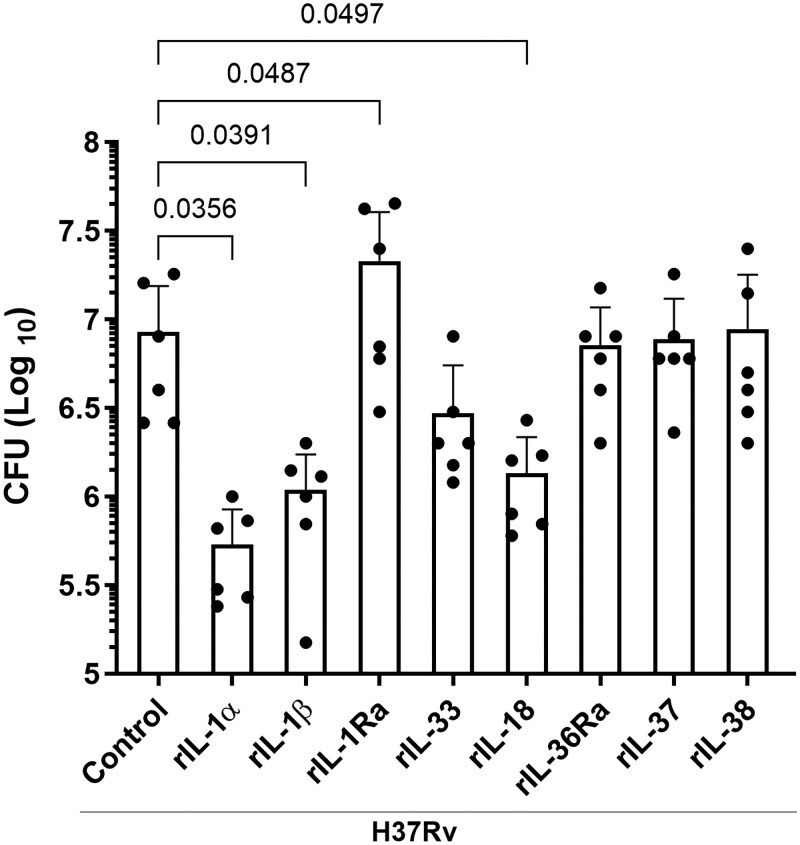 Figure 2
Figure 2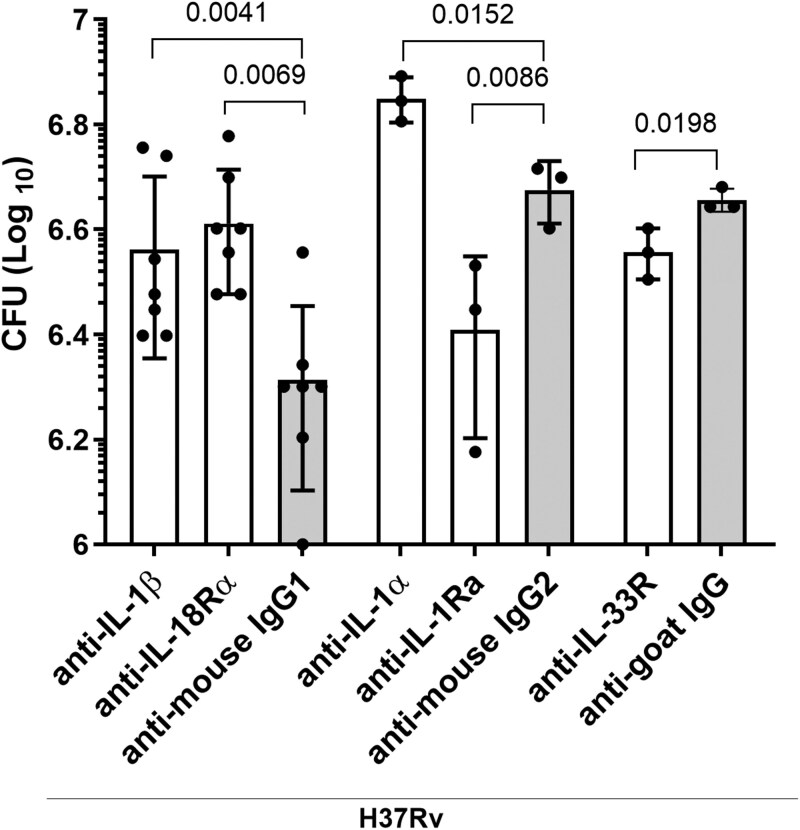 Figure 3
Figure 3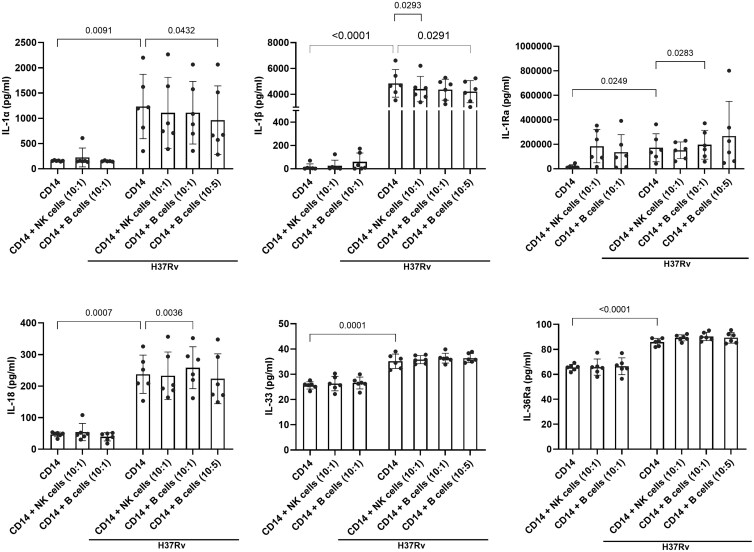 Figure 4
Figure 4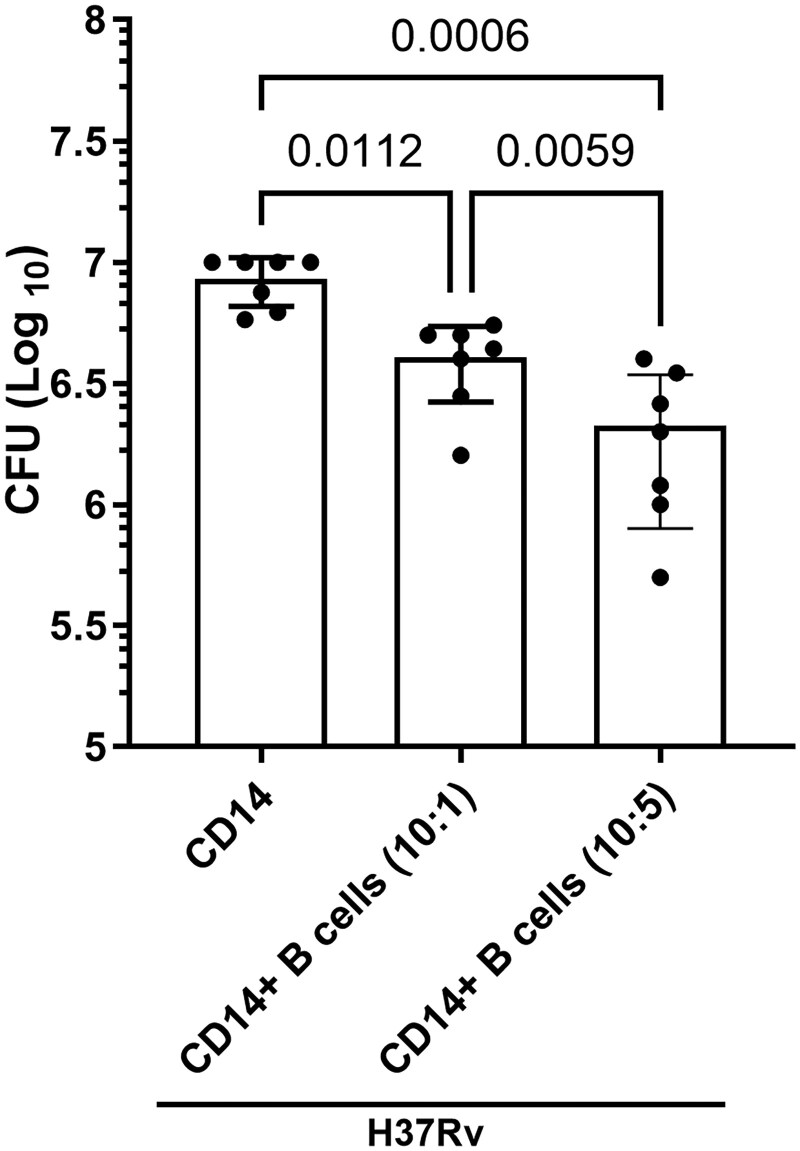 Figure 5
Figure 5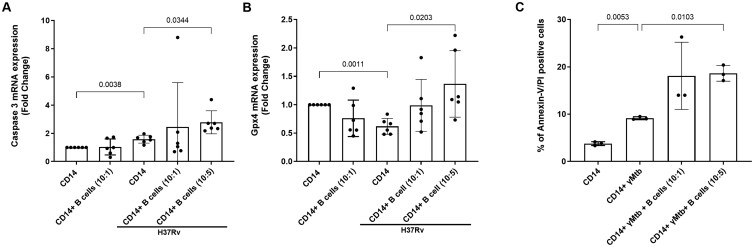 Figure 6
Figure 6- —Cain Foundation for Infectious Disease Research
- —University of Texas Health Science Center at Tyler, Texas, United States
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTuberculosis Research and Epidemiology · Infectious Diseases and Tuberculosis · Mycobacterium research and diagnosis
Introduction
Tuberculosis (TB) is the second leading cause of death from a single infectious agent, killing approximately 1.3 million people annually.1 Alveolar macrophages are involved in the initial innate defense against various lung pathogens, including Mycobacterium tuberculosis (Mtb).2^,^3 Various cytokines released by macrophages play important roles during Mtb infection.4 In-depth analysis of cytokine release from alveolar macrophages will provide a better understanding of immune responses to Mtb.5^,^6
The IL-1 superfamily (ILSF) of cytokines produced by antigen-presenting cells (APCs) plays an important role in various diseases and infections, including Mtb infection.7^,^8 IL-1α, IL-1β, IL-18, IL-33, IL-36α, IL-36β, IL-36γ, IL-37, IL-38, IL-1Ra, and IL-36Ra are the 11 soluble factors of the ILSF of cytokines.8 Acute respiratory distress syndrome of the lungs, influenza, and COVID-19 are examples of diseases that involve both cytokine storms and IL-1α and IL-1β.9 Some viral infections of the lungs present with increased amounts of IL-33.9 IL-1α and IL-1β have important roles in controlling bacterial infections in mice.10 The ILSF of cytokines plays a major role in the immune response to TB.11–15 Reports have shown the utility of circulating IL-1Ra as a marker of TB.4 In patients with active TB, elevated levels of IL-1Ra and reduced levels of IL-1α and IL-1β after BCG stimulation have been noted.4 The cytokines IL-1α and IL-33 also act as damage-associated molecular patterns,16 and necrotic cells release IL-1α to function as an alarm for the initiation and recruitment of cytokine/chemokine and effector immune cells.9 IL-18 produced by alveolar macrophages and PBMCs enhances IFN-γ production and provides protective immunity against TB.17^,^18 However, the role of ILSF cytokines in Mtb growth in human macrophages has not been comprehensively elucidated.
T cells are known to regulate ILSF cytokine production and Mtb growth in APCs.19^,^20 B cells play an important role during Mtb infection.21–23 During chronic TB infection, B cells play a proinflammatory role by increasing the number of CD4^+^ T cells available so that they can generate more IFN-γ to allow for the development of granulomas.21 B cells can regulate the balance of pro- and anti-inflammatory responses in TB granulomas.24^,^25 Antigen-specific B cells control Mtb infection through the localization of T follicular helper–like cells in granuloma-associated lymphoid follicles.26 There is limited information available on the role of B cells in Mtb growth and ILSF cytokine production by human APCs.23^,^27^,^28
In the present study, we evaluated the production of ILSF cytokines by human monocyte-derived macrophages (MDMs) and investigated the comprehensive role of various ILSF cytokines in Mtb growth. We also determined the effects of B cells on ILSF cytokine production and Mtb growth in human monocytes.
Materials and methods
Ethical approval and blood sample collection
Healthy human volunteer blood samples were collected for this study, and written consent was obtained from the study participants. All studies were approved by the Institutional Review Board of the University of Texas Health Science Center at Tyler (HSC-1022) and Saint Louis University (34724 and 33195).
Antibodies and reagents
The following antibodies and human recombinant proteins were obtained from R&D Systems: anti-ST2/IL-33R antibody (catalog # AF-523), anti-IL-1β/IL-1F2 antibody (catalog # MAB601), anti-IL-1α/IL-1F1 antibody (catalog # MAB200), anti-IL-1Ra/IL-1F3 antibody (catalog # MAB280), anti-IL-18Rα/IL-1R5 antibody (catalog # MAB840), anti-IL-36/IL-1F3 antibody (catalog # AF1078), anti-IL-37/IL-1F7 antibody (catalog # AF1975), anti-IL-38/IL-1F10 antibody (catalog # AF2427), anti-rIL-37/IL-1F7 protein (catalog # 1975-IL), mouse monoclonal IgG1 antibody (catalog # MAB002), mouse monoclonal IgG2A antibody (catalog # MAB003), and goat polyclonal IgG antibody (catalog # AB-108-C). rIL-1α (catalog # 570002), rIL-1β (catalog # 579402), rIL-18 (catalog # 592102), rIL-33 (catalog # 581802), rIL-1Ra/IL-1RN (catalog # 553902), and rIL-36Ra/IL-1F5 (catalog # 760904) were obtained from BioLegend, and the rIL-38/IL-1F10 protein (catalog # NBP2-52047) was obtained from Novus Biologicals.
Isolation of monocytes, B cells, and NK cells
Peripheral blood mononuclear cells (PBMCs) were isolated via Ficoll–Paque gradient differential centrifugation as previously described.29 On day 0, CD14^+^ monocytes were isolated via magnetic beads, and their purity was checked according to the manufacturer’s instructions (Miltenyi Biotec, catalog # 130-050-201). Isolated CD14^+^ monocytes (>95% pure cells) were plated in multiple wells of a 12-well plate at 1.2–2 × 10^6^ cells/well in complete cell culture RPMI medium supplemented with 10% heat-inactivated human serum, 1% sodium pyruvate, and 1% penicillin-streptomycin. Monocytes were incubated at 37 °C in a humidified CO_2_ incubator for 5 days to allow them to differentiate into MDMs. Adhered macrophages from the same donors were washed with 1× HBSS. Some cells were used as control and others were infected with Mtb. Some of the Mtb-infected cells were treated with either recombinant proteins or neutralizing antibodies.
For some experiments, CD14^+^ monocytes were either cocultured with autologous NK cells or B cells to determine the ILSF cytokine production, Mtb growth, and the cell death mechanisms. We used NK cells (10 monocytes with one NK cell) as a positive control30^,^31 and 2 different concentrations of B cells (10 monocytes with one B cell or 5 B cells). PBMCs were isolated from blood, and CD14^+^ monocytes, Pan B cells, and total NK cells were separated via magnetic beads from autologous PBMCs according to the manufacturer’s instructions (Miltenyi Biotec; Pan B Cell Isolation Kit, catalog # 130-101-638 and NK Cell Isolation Kit, catalog # 130-092-657) and cocultured on the same day of the experiment (day 0). Purity of NK cells (CD3^−^NK1.1^+^; >95%) and B cells (CD3^−^CD19^+^; >92%) were checked by flow cytometry (Thermo Attune NxT and BD LSR Fortessa X-20) and used for the subsequent experiments (Fig. S1A, B).
Macrophage/monocyte infection, coculture with B cells and NK cells, and ILSF proteins/cytokines
Some of the MDMs were infected with Mtb H37Rv at a multiplicity of infection of 1:2.5 in antibiotic-free medium and incubated for 2 hours at 37 °C in a humidified CO_2_ incubator. After incubation, the cells were washed with 1× HBSS for a total of 3 washes to remove extracellular bacilli and incubated with antibiotic-free RPMI medium containing 10% heat-inactivated human serum and 1% sodium pyruvate. In some experiments, recombinant proteins were added to MDMs cultured with Mtb at concentrations of 10 ng/mL (rIL-1α, rIL-1β, rIL-33, rIL-36Ra, and rIL-38), 100 ng/mL (rIL-1Ra and rIL-18), and 3 µg/mL (rIL-37). Concentrations for recombinant proteins were selected based on previous published studies and manufacturers recommendations.32^,^33 For neutralization experiments, concentrations were determined as per the manufacturer’s instructions and previously published studies. Neutralizing anti-IL-1α (anti-mouse IgG2k), anti-IL-1β (anti-mouse IgG1k), anti-IL-1Ra (anti-mouse IgG2k), anti-IL-18Rα (anti-mouse IgG1k), and anti-IL-33R (anti-goat IgG) antibodies or their IgG isotype controls were used at a concentration of 10 µg/mL.34–37
For B cell and NK cell coculture experiments, freshly isolated monocytes, B cells, and NK cells were counted via the trypan blue exclusion method. In some experiments, freshly isolated B cells were added to CD14^+^ monocytes at a concentration of 1 or 5 cells per 10 monocytes (1 × 10^6^ CD14^+^ to 1 or 5 × 10^5^ CD19^+^ B cells), and NK cells were added to monocytes at a concentration of 1 cell per 10 monocytes (1 × 10^6^ CD14^+^ to 1 × 10^5^ NK cells) on the same day that CD14^+^ cells were isolated. The cocultured cells (CD14: B or CD14: NK) were infected with Mtb H37Rv immediately. At 24, 48, 72, or 120 hours postinfection, the cell pellets were used to determine the bacterial burden and RNA isolation, and the cell culture supernatants were collected for various cytokine/chemokine analyses.
Determination of the bacterial burden in Mtb-infected MDMs or monocytes
At 120 hours postinfection, the MDMs or monocytes were lysed in 1 mL of 7H9 broth containing 0.25% SDS and 1% BSA, serially diluted, and plated on 7H10 agar plates (Thermo Scientific, catalog # 01600) in triplicate. Colony-forming units (CFUs) were determined 21 days after incubation by counting the number of colonies on the agar plates.
Measurement of cytokine production by ELISA
The following cytokines were measured in the cell culture supernatants: IL-6 (catalog # 430504, BioLegend), IL-10 (catalog # 430604, BioLegend), TNF-α (catalog # 430204, BioLegend), IL-1β (catalog # 437004, BioLegend), IL-37 (catalog # DY1975, R&D Systems), and IL-38 (catalog # DY9110, R&D Systems). In some experiments, IL-1 family cytokines were measured via a multiplex ELISA kit via magnetic Luminex assays (catalog # CUSTOM-LXSA-H-16 lot # C0008953, R&D Systems).
RNA isolation and qRT-PCR
Total RNA was extracted from Mtb-infected MDMs or monocytes via TRIzol (Invitrogen) according to the manufacturer’s instructions. The isolated RNA was quantified and reverse transcribed (iScript Reverse Transcription SuperMix kit; Bio-Rad), and real-time PCR was performed on a Bio-Rad CFX384 instrument with iTaq Universal SYBR Green Supermix (Bio-Rad). mRNA expression was quantified via the 2^−ΔΔCt^ method, and gene expression levels were normalized to those of β-actin internal controls. The forward and reverse primers used for this study are listed in Table S1.
Annexin V/propidium iodide staining
Isolated CD14^+^ monocytes were cocultured with B cells and stimulated with γ-Mtb (gamma-irradiated Mtb H37Rv) at 10 µg/mL. After 48 hours of incubation, the cells were collected and stained with Annexin V and propidium iodide (PI), and staining results were acquired via flow cytometry (Thermo Attune NxT and BD LSR Fortessa X-20).
Statistical analysis
Statistical analysis was performed via Prism 10.2.00 software (GraphPad). The same donor samples were cultured in multiple wells. For statistical analysis, we have used either 2-tailed paired t-test or repeated-measures one-way ANOVA since the comparisons were made between the same donor samples (control vs treated) and considered as same cohort. To reduce type 1 error (false-positive), we have used post hoc Fisher least significant difference (LSD) test or Tukey multiple comparisons test. The results are shown as the mean ± SD. P < 0.05 was considered statistically significant.
Results
ILSF cytokine production by Mtb H37Rv–infected human MDMs
Human MDMs were infected with Mtb H37Rv as described in the Materials and methods. Cytokine levels in culture supernatants were measured by multiplex ELISA after 24 hours. In 17 to 20 healthy donors, Mtb infection significantly increased IL-1α (P = 0.0018, Fig. 1A), IL-1β (P < 0.0001, Fig. 1B), IL-18 (P = 0.0128, Fig. 1C), and IL-37 (P = 0.0427, Fig. 1D) production (among the 8 ILSFs measured) by MDMs while reducing IL-1Ra (P = 0.0056, Fig. 1E) production compared with that in uninfected MDM culture supernatants. There was no significant difference in IL-33, IL-36Ra, or IL-38 production (Fig. 1F–H) by MDMs after Mtb infection.
ILSF cytokine production by Mtb H37Rv–infected MDMs. Human CD14+ monocytes were isolated from 17 to 20 healthy donors, converted to MDMs, and infected with Mtb H37Rv as described in the Materials and methods. After 24 hours, the levels of the cytokines IL-1α (A), IL-1β (B), IL-18 (C), IL-37 (D), IL-1Ra (E), IL-33 (F), IL-36Ra (G), and IL-38 (H) in the culture supernatants were measured via multiplex ELISA. Data are shown as mean ± SD. The statistical analysis was performed via 2-tailed paired t-tests, and P values are shown. ns, not significant.
Effect of recombinant ILSF cytokines on Mtb H37Rv growth in human MDMs
We determined the effects of ILSF cytokines on Mtb growth in human MDMs. Some infected MDMs were cultured with or without recombinant ILSF cytokines. After 120 hours, the bacterial burden was measured (Fig. 2). Mtb-infected MDMs cultured without any human recombinants from the ILSF were used as controls. Compared with that of the control, the bacterial burden was significantly decreased when Mtb H37Rv-infected MDMs were cultured with human recombinant IL-1α (P = 0.0356), IL-1β (P = 0.0391), or IL-18 (P = 0.0497) (Fig. 2). In contrast, compared with the control, the CFUs increased when Mtb-infected MDMs were cultured with human recombinant IL-1Ra (P = 0.0487). There was no significant difference in bacterial growth between the control and the Mtb H37Rv-infected MDMs cultured with rIL-33, rIL-36Ra, rIL-37, or rIL-38 (Fig. 2).
Recombinant ILSF cytokines alter Mtb H37Rv growth in human MDMs. Human CD14+ monocytes were isolated, converted to MDMs, and infected with Mtb H37Rv. Some of the infected MDMs were cultured with various recombinant ILSF cytokines. rIL-1α, rIL-1β, rIL-36Ra, rIL-33, and rIL-38 were used at 10 ng/mL; rIL-1Ra and rIL-18 were used at 100 ng/mL; and rIL-37 was used at 3 µg/mL. After 5 days, the CFUs were determined as described in the Materials and methods. Data are shown as mean ± SD. MDMs from 6 donors were used for this study. The statistical analysis was performed via by repeated-measures one-way ANOVA followed by post hoc Fisher LSD test, and P values are shown.
Neutralization of ILSF cytokine antibodies altered Mtb growth in MDMs
The above findings revealed that the IL-1 subfamily cytokines IL-1α, IL-1β, and IL-18 inhibited Mtb growth and that IL-1Ra increased the bacterial burden in MDMs (Fig. 2). Next, we determined whether neutralization of these cytokines affects the bacterial burden in Mtb-infected MDMs. Some infected MDMs were cultured with and without anti-ILSF cytokine-blocking antibodies. Mtb-infected MDMs cultured with IgG isotype control antibodies were used as internal controls. After 120 hours, the bacterial burden was determined (Fig. 3). The bacterial burdens of Mtb-infected MDMs cultured with neutralizing antibodies against human IL-1β (P = 0.0041; 3.6 × 10^6^ ± 1.3 × 10^6^ vs 2.05 × 10^6^ ± 7.8 × 10^5^), IL-18Rα (P = 0.0069; 4.08 × 10^6^ ± 1.09 × 10^6^ vs 2.05 × 10^6^ ± 7.8 × 10^5^), and IL-1α (P = 0.0152; 7.06 × 10^6^ ± 7.0 × 10^5^ vs 4.7 × 10^6^ ± 6.4 × 10^5^) were significantly greater than those of their respective control antibodies (Fig. 3). In contrast, bacterial growth significantly decreased when Mtb-infected MDMs were cultured with neutralizing antibodies against human IL-1Ra (P = 0.0086; 2.56 × 10^6^ ± 9.7 × 10^5^ vs 4.7 × 10^6^ ± 6.4 × 10^5^) and IL-33R (P = 0.0198; 3.6 × 10^6^ ± 4 × 10^5^ vs 4.53 × 10^6^ ± 2.3 × 10^5^) compared with the IgG controls (Fig. 3).
Effect of anti-ILSF cytokines on Mtb H37Rv growth in human MDMs. Human CD14+ monocytes were isolated, converted to MDMs, and infected with Mtb H37Rv. Some of the infected MDMs were cultured with neutralizing antibodies against various ILSF cytokines (10 µg/mL of each cytokine was used). Five days postinfection, the bacterial burden was determined as described in the Materials and methods. Data are shown as mean ± SD. MDMs from 3 to 7 donors were used for this study. The statistical analysis was performed via 2-tailed paired t-tests; the respective IgG isotype controls were compared with each neutralization antibody, and P values are shown.
Effect of B cells on ILSF cytokine production by Mtb H37Rv–infected monocytes
We determined the effect of B cells on ILSF cytokine production by Mtb-infected monocytes. NK cells are known to enhance the functions of Mtb-infected monocytes,29^,^38^,^39 so we also cultured Mtb-infected monocytes with NK cells as a control. We used freshly isolated CD14^+^ monocytes instead of MDMs and cultured them with autologous B or NK cells to avoid using blood from the same donor twice. Freshly isolated human monocytes were infected with Mtb H37Rv, and some of the Mtb-infected monocytes were cultured with 2 different ratios of freshly isolated B cells (10 monocytes with one or 5 B cells) (Fig. 4 and Fig. S1A, B). Some of the Mtb-infected monocytes were cultured with freshly isolated NK cells (10 monocytes with one NK cell) (Fig. 4). After 48 hours, the levels of the cytokines IL-1α, IL-1β, IL-1Ra, IL-18, IL-33, IL-36Ra, IL-6, IL-10, and TNF-α in the culture supernatants were measured via ELISA (Fig. 4 and Fig. S1C). Compared with uninfected monocytes, after 48 hours, Mtb-infected monocytes without B cells produced significantly greater amounts of IL-1α (P = 0.0091), IL-1β (P < 0.0001), IL-1Ra (P = 0.0249), IL-18 (P = 0.0007), IL-33 (P = 0.0001), IL-36Ra (P < 0.0001), IL-6 (P = 0.0130), IL-10 (P < 0.0001), and TNF-α (P = 0.0131) (Fig. 4 and Fig. S1C). Although Mtb infection reduced IL-1Ra production by MDMs after 24 hours (Fig. 1E), IL-1Ra production increased after 48 hours (P = 0.0249, Fig. 4) compared with that in uninfected monocytes. Coculturing B cells (10 monocytes with 5 B cells) with Mtb-infected monocytes for 48 hours resulted in reduced production of IL-1α (P = 0.0432), IL-1β (P = 0.0291), IL-6 (P = 0.0197), and TNF-α (P = 0.0181) compared with that in Mtb-infected monocytes without B cells (Fig. 4 and Fig. S1C). In contrast, coculturing B cells (10 monocytes with one B cell) with Mtb-infected monocytes for 48 hours resulted in increased production of IL-1Ra (P = 0.0283) and IL-18 (P = 0.0036) compared with that of Mtb-infected monocytes without B cells (Fig. 4). A similar increase in IL-10 production (P = 0.0310) was noted when 10 monocytes and 5 B cells were cocultured with Mtb-infected monocytes for 48 hours (Fig. S1C). There was no significant difference in IL-33 or IL-36Ra production between Mtb-infected monocytes and Mtb-infected monocytes cultured with B cells (Fig. 4). NK cells reduced IL-1β (P = 0.0293) and IL-10 (P = 0.0278) production by Mtb-infected monocytes (Fig. 4 and Fig. S1C). NK cells had no effect on the production of other ILSF cytokines, and IL-6 and TNF-α, by Mtb-infected monocytes (Fig. 4 and Fig. S1C).
Effect of B cells on ILSF cytokine production by Mtb H37Rv–infected CD14+ monocytes. Human CD14+ monocytes were isolated and infected with Mtb H37Rv. Some of the infected monocytes were cultured with different ratios of B cells (monocyte to B-cell ratios of 10:1 and 10:5) or NK cells (monocyte to NK cell ratios of 10:1), as mentioned in the Materials and methods. After 48 hours, the IL-1α, IL-1β, IL-1Ra, IL-18, IL-33, and IL-36Ra levels in the culture supernatants were measured via multiplex ELISA. Data are shown as mean ± SD. Monocytes from 6 donors were used for this study. The statistical analysis was performed via repeated-measures one-way ANOVA followed by post hoc Fisher LSD test, and P values are shown.
Effect of B cells on Mtb H37Rv growth in human monocytes
We determined the effect of B cells on Mtb growth in monocytes. In this experiment, some of the Mtb-infected monocytes were cultured with 2 different ratios of freshly isolated B cells (10 Mtb-infected monocytes with one or 5 B cells) (Fig. 5). As a control, some Mtb-infected monocytes were cultured for 120 hours without B cells. We found that B cells significantly reduced Mtb H37Rv growth in monocytes compared with that in monocytes infected with Mtb alone (Fig. 5). Culturing Mtb-infected monocytes with more B cells reduced Mtb growth more efficiently. (10:1 ratio, P = 0.0112; 10:5 ratio, P = 0.0006) (Fig. 5).
B cells reduce the bacterial burden of Mtb H37Rv–infected monocytes. Human CD14+ monocytes were isolated and infected with Mtb H37Rv, and some of the infected monocytes were cultured with different ratios (monocyte to B-cell ratios of 10:1 and 10:5) of B cells, as mentioned in the Materials and methods. After 120 hours, the bacterial burden was determined. Data are shown as mean ± SD. Monocytes from 7 donors were used for this study. The statistical analysis was performed via repeated-measures one-way ANOVA followed by Tukey multiple comparisons test, and P values are shown.
B cells increased death pathway gene expression in Mtb H37Rv–infected human monocytes
We determined whether B cells enhance the apoptosis of Mtb-infected monocytes to restrict their growth. Uninfected and Mtb-infected monocytes were cultured with or without B cells. B cells were added at 2 different concentrations (10 Mtb-infected monocytes with one or 5 B cells) (Fig. 6). After 48 hours of Mtb infection, expression of various genes involved in cell death was determined via qRT-PCR. As shown in Fig. 6A and B, culturing Mtb-infected monocytes with B cells (10 *Mtb-*infected monocytes with 5 B cells) significantly upregulated Caspase 3 expression (P = 0.0344) and GPX4 expression (P = 0.0203). There was no difference in the expression of LC3B, Caspase 1, RIPK3, or MLKL among the groups (Fig. S2A–D). Furthermore, we found that B cells enhance (10 *Mtb-*infected monocytes with 5 B cells) the apoptosis of γ-Mtb-cultured monocytes, as determined by Annexin V/PI staining (P = 0.0103; Fig. 6C and Fig. S2E).
Mtb stimulation alters cell death pathway gene expression and apoptosis of human monocytes in the presence of B cells. Human CD14+ monocytes were isolated and infected with Mtb H37Rv. Some of the infected monocytes were cultured with B cells at different concentrations (monocyte to B-cell ratios of 10:1 and 10:5). (A and B) After 48 hours, the expression of genes involved in various death pathways was determined via qRT-PCR. Monocytes and B cells from 6 donors were used for this study. (C) Monocytes were cultured with B cells (monocyte to B-cell ratios of 10:1 and 10:5) and stimulated with γ-Mtb at 10 µg/mL. At 48 hours poststimulation, apoptosis was determined via Annexin V/PI staining. Data are shown as mean ± SD (n = 3). The statistical analysis was performed via repeated-measures one-way ANOVA followed by post hoc Fisher LSD test, and P values are shown.
Discussion
ILSF cytokines play important roles during Mtb infection.4^,^7^,^40^,^41 However, only limited information is available concerning the comprehensive role of these cytokines during human Mtb infection.7^,^42 In the present study, we found that compared with uninfected MDMs, human MDMs infected with Mtb produced significantly higher levels of IL-1α, IL-1β, IL-18, and IL-37 and lower levels of IL-1Ra. Human recombinant IL-1α, IL-1β, and IL-18 significantly reduced Mtb growth in MDMs while human recombinant IL-1Ra enhanced its growth. Furthermore, we found that neutralization of the cytokines IL-1α, IL-1β, and IL-18α significantly increased the bacterial burden in Mtb-infected MDMs. In contrast, neutralization of IL-1Ra and IL-33R significantly reduced the bacterial burden. We also found that B cells significantly increased IL-1Ra, IL-18, and IL-10 production and reduced IL-1α, IL-1β, IL-6, and TNF-α production and the bacterial burden in Mtb-infected monocytes.
ILSF cytokines play distinct roles in various diseases.10^,^43^,^44 It is well known that innate immune cells produce the IL-1 family of cytokines and exhibit protective immunity against TB.7^,^20^,^45–47 IL-1α/β double knockout mice infected with Mtb exhibited larger granulomatous lesions and greater bacterial loads in their lungs.48 Similarly, IL-1R^−/−^ mice are highly susceptible to Mtb infection through a process mediated by IL-1α production.12 In the present study, we found that MDMs infected with Mtb secreted significantly higher levels of IL-1α, IL-1β, IL-18, and IL-37 and lower amounts of IL-1Ra than uninfected MDMs did (Fig. 1). Control and Mtb-infected MDMs produced similar amounts of IL-33, IL-36Ra, and IL-38. IL-1α, IL-1β, IL-18, and IL-37 production depends on caspase or inflammasome activation.15^,^49–52 Our findings suggest that early Mtb infection (24 hours after infection) initiates caspase- or inflammasome-dependent events that initiate a proinflammatory reaction to eradicate Mtb in macrophages.
Limited information is available about the effects of recombinant and anti-IL-1 cytokines on various diseases and infections, including Mtb infection.53 In Mtb-infected MDMs, human recombinant IL-1α, IL-1β, and IL-18 reduced Mtb growth while human recombinant IL-1Ra enhanced Mtb growth (Fig. 2). Blocking antibodies against IL-1α, IL-1β, or IL-18 enhanced Mtb growth in MDMs. In contrast, blocking antibodies against IL-1Ra or IL-33R decreased Mtb growth in MDMs (Fig. 3). Exogenous addition of IL-1α and IL-1β was reported to reduce mycobacterial growth in macrophages.54 Using animal models, several other studies have demonstrated the essential roles of IL-1α, IL-1β, and IL-18 in the control of Mtb infection.54–57 IL-18 plays a protective role in both mice and humans.17^,^18^,^58 Taken together, our findings demonstrate that IL-1α, IL-1β, IL-18, and IL-1Ra have similar effects in both animal models and human MDMs.
Although IL-1Ra, IL-1α, and IL-1β are found in the cytoplasm, the anti-inflammatory effects of IL-1Ra differ from those of IL-1α and IL-1β.16 Excess amounts of IL-1Ra increase the susceptibility of hosts to Mtb infection.14^,^15 Our current findings further demonstrate that IL-1Ra can enhance Mtb growth in human MDMs (Fig. 2). Previous studies have demonstrated that IL-33 can clear ongoing Mtb infection.59 Recombinant IL-33 moderately reduced Mtb growth in MDMs (Fig. 2), and anti-IL33R also reduced Mtb growth in MDMs (Fig. 3), suggesting that excess IL-33 can function as a proinflammatory cytokine. Murine macrophages produce IL-36 upon Mtb infection, and IL-1β and IL-18 can also induce the production of IL-36; IL-36 signaling then enhances antimicrobial peptide production and Mtb growth inhibition.60 IL-36 also enhances antimicrobial peptide production through cholesterol metabolism in the THP-1 cell line.61 However, we found that the inability of human MDMs to produce IL-36Ra and treatment with exogeneous recombinant IL-36Ra had no effect on Mtb growth, demonstrating the differences among mouse, THP-1 and human studies.60^,^61
IL-37 is highly expressed in various autoimmune diseases, such as rheumatoid arthritis, systemic lupus erythematosus, and Sjögren syndrome.40 Compared with those in healthy donors, plasma IL-37 levels are higher in TB patients, and IL-37 inhibits the phagocytic capacity of Mtb-infected THP-1 cells and induces the M2 phenotype.62 We found that Mtb-infected human MDMs produce significant amounts of IL-37 and that human recombinant IL-37 has no effect on Mtb growth in human MDMs (Fig.1 and Fig. 2). IL-38 plays a role in viral infections such as hepatitis B and C,63 and the plasma IL-38 levels of type 2 diabetes patients with latent TB infection are significantly higher than those of individuals without latent TB infection. In the present study, Mtb-infected and uninfected human MDMs produced equal amounts of IL-38, and recombinant IL-38 had no effect on Mtb growth in MDMs (Figs. 1 and 2).
B cells play multiple roles depending on the circumstances. For example, IL-1Ra deficiency can influence the actions of a subtype of B cells on the immune system’s proinflammatory cascades.64 B cells can mature through the help of IL-1α and IL-1β stimulation.65 Regulatory B cells (Bregs) secrete IL-10 to suppress the human immune system, resulting in the control of some autoimmune diseases.66 These findings highlight the dual status of B cells, which function as chaperones of both the pro- and anti-inflammatory responses. In this current study, compared with Mtb-infected monocytes without B cells, the culture of B cells with Mtb-infected monocytes significantly reduced the production of IL-1α, IL-1β, IL-6, and TNF-α; increased the production of IL-1Ra, IL-18, and IL-10 (Fig. 4 and Fig. S1C); and reduced Mtb growth (Fig. 5). For the B-cell coculture experiments, we used monocytes instead of MDMs for autologous culture to avoid using blood from the same donor twice. Recent studies have demonstrated that B cells play multiple roles in Mtb infection.67–72 During chronic Mtb infection, a significant number of B cells are found on the boundaries of granulomas.5 B cells have dual properties during the chronic stages of Mtb infection to carry out the chief executive functions of the adaptive immune response. B cells interact with CD4^+^ helper T cells in the granulomas of Mtb-infected mice and enhance proinflammatory responses.21 The ability of B cells to inhibit these excess inflammatory cytokines is important in Mtb granulomas to reduce inflammatory pathology and the spread of infection.67 IL-1α and IL-1β can induce B cell maturation, differentiation, and immunoglobulin production, suggesting that B-cell and IL-1 crosstalk occurs.65 Our findings further demonstrated that B cells can inhibit IL-1α, IL-1β, IL-6, and TNF-α production and increase IL-1Ra, IL-18, and IL-10 production to suppress inflammation during the chronic stages of human Mtb infection (Fig. 4 and Fig. S1C).
In response to Mtb, NK cells produce IFN-γ and inhibit Mtb growth in monocytes.31^,^39 Therefore, we also measured ILSF cytokine production by Mtb-infected monocytes in the presence of NK cells. NK cells reduced IL-1β and IL-10 production by Mtb-infected monocytes but had no effect on the production of other ILSF cytokines, such as IL-6 and TNF-α, by Mtb-infected monocytes (Fig. 4 and Fig. S1C). These results demonstrate that NK cells inhibit ILSF cytokine production by Mtb-infected monocytes.
B cells are unable to suppress proinflammatory immune responses when there is a deficiency of IL-1Ra.64 During the chronic stages of Mtb infection, excess IL-1 production can cause pathology and tissue damage.73 Antibiotic treatment along with IL-1R blockade can reduce disease severity and inflammation in Mtb-infected mice and macaques.73 Our findings and the above published studies demonstrated that B cells enhance IL-1Ra production to reduce immunopathology by inhibiting the production of the proinflammatory cytokines IL-1α, IL-1β, IL-6, and TNF-α. Bregs are a unique type of B cell that produce IL-10 to suppress proinflammatory immune responses and are necessary for the proper control of autoimmune disorders.66^,^74^,^75 Proinflammatory cytokines such as IL-1β can activate Bregs.76^,^77 In the present study, B cells cocultured with Mtb-infected monocytes significantly increased IL-1Ra, IL-18, and IL-10 production (Fig. 4 and Fig. S1C), suggesting a possible role of IL-10–producing Bregs in regulating excess inflammation in granulomas.
IL-1Ra is made in 2 forms: a secretory IL-1Ra form and a cytoplasmic IL-1Ra form.78 B cells may play a more robust role in activating all forms of IL-1Ra via their interaction with Mtb-infected monocytes (Fig. 4). Further studies are needed to determine whether Bregs or B cells initiate a downstream signaling pathway that leads to the activation of IL-1Ra in the cytosol of human monocytes ([Figs. 4–6](#vlaf046-F4 vlaf046-F5 vlaf046-F6)). In the promoter regions of IL-1Ra, secretory IL-1Ra has a TATA box with a similar sequence of nucleotides for NF-kB, but cytoplasmic IL-1Ra does not have a TATA box with that sequence of nucleotides.79 The binding of IL-1Ra to its receptor IL-1R1 leads to an anti-inflammatory response.16
B cells at 2 different ratios (10:1 and 10:5 [monocytes to B cells]) inhibited Mtb growth in monocytes (Fig. 5). Culturing B cells with Mtb-infected monocytes significantly increased Caspase 3 and GPX4 mRNA expression and increased apoptosis at a ratio of 10:5 (Fig. 6A–C). We found that B cells cocultured with monocytes at a ratio of 10:5 efficiently inhibited Mtb growth compared with those in the 10:1 ratio (Fig. 5). Prior studies demonstrated that a reduced Mtb burden was associated with an increased number of B cells and anti-Mtb antibodies, such as IgA and IgG.80 B cells depleted in nonhuman primate cynomolgus macaques infected with Mtb exacerbated the bacterial burden and inflammation.25 Mice lacking B cells experienced worsening TB disease due to the recruitment of neutrophils.81 During Mtb infection, marginal zone B cells (MZB cells) accumulate in the mice lungs and spleen.82 These MZB cells provide systemic control of Mtb infection and display activated and memory-like phenotypes.82
Antibody production is the major function of B cells in various diseases and infections.83 Antibody production to enhance monocyte-mediated cytotoxicity and enhance the induction of Caspase 3 expression by Mtb-infected monocytes may be the major mechanism by which Mtb growth is inhibited (Figs. 5 and 6). It is possible that B cells enhance the Fc portion of IgG1 or IgG3 Abs (constant region) bound to target cells, which may increase FcγRIIA (CD16)–mediated killing of target cells to restrict Mtb growth.84 However, further studies are needed to understand the B cell–mediated inhibition of Mtb growth.
In conclusion, among the 8 ILSF cytokines, 3 (IL-1α, IL-1β, and IL-18) restrict Mtb growth in human MDMs, one enhances growth (IL-1Ra), and 4 (IL-33, IL-36Ra, IL-37, and IL-38) play no role in Mtb growth. B cells increased IL-1Ra, IL-18, and IL-10 production and inhibited IL-1α, IL-1β, IL-6, and TNF-α production by Mtb-infected monocytes. A better understanding of the mechanisms by which ILSF family cytokines affect Mtb growth in human alveolar macrophages and the role of B cells in the production of these cytokines may facilitate the development of therapies to prevent or treat TB.
Supplementary Material
vlaf046_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Global tuberculosis report 2023. Geneva: World Health Organization; 2023.
- 2Chandra P , Grigsby SJ, Philips JA. Immune evasion and provocation by Mycobacterium tuberculosis. Nat Rev Microbiol. 2022;20:750–766.35879556 10.1038/s 41579-022-00763-4PMC 9310001 · doi ↗ · pubmed ↗
- 3Sadee W et al Human alveolar macrophage response to Mycobacterium tuberculosis: immune characteristics underlying large inter-individual variability. Commun Biol. 2025;8:950.40550843 10.1038/s 42003-025-08337-9PMC 12185700 · doi ↗ · pubmed ↗
- 4Llibre A et al Tuberculosis alters immune-metabolic pathways resulting in perturbed IL-1 responses. Front Immunol. 2022;13:897193.36591308 10.3389/fimmu.2022.897193 PMC 9795069 · doi ↗ · pubmed ↗
- 5Hunter L et al Dynamics of macrophage, T and B cell infiltration within pulmonary granulomas induced by Mycobacterium tuberculosis in two non-human primate models of aerosol infection. Front Immunol. 2022;12:776913.35069548 10.3389/fimmu.2021.776913 PMC 8770544 · doi ↗ · pubmed ↗
- 6Berrington WR , Hawn TR. Mycobacterium tuberculosis, macrophages, and the innate immune response: does common variation matter? Immunol Rev. 2007;219:167–186.17850489 10.1111/j.1600-065X.2007.00545.x PMC 2859969 · doi ↗ · pubmed ↗
- 7Silvério D et al Advances on the role and applications of interleukin-1 in tuberculosis. m Bio. 2021;12:e 0313421.34809460 10.1128/m Bio.03134-21PMC 8609357 · doi ↗ · pubmed ↗
- 8Van Den Eeckhout B , Tavernier J, Gerlo S. Interleukin-1 as innate mediator of T cell immunity. Front Immunol. 2021;11:621931.33584721 10.3389/fimmu.2020.621931 PMC 7873566 · doi ↗ · pubmed ↗